Abstract
Oceanic islands of volcanic origin have a narrow shelf and a steep slope that should lead to considerable spatial overlap among coastal and oceanic fauna. During six pelagic surveys in the Canarian archipelago, Eastern Central Atlantic, over 65,000 fishes belonging to 211 species were collected at depths between 8 and 1035 m. The mesopelagic families of the lanternfishes (Myctophidae) and the bristlemouths (Gonostomatidae) accounted for about 50% of all specimens. By multivariate classification and ordination methods four different assemblages associated with mesopelagic, epipelagic–oceanic or coastal habitats could be identified. Two of these assemblages were coastal, differing in the proportion of meso- and epipelagic species. These data indicate intense horizontal migrations of mesopelagic fishes (mainly Myctophidae) into the neritic realm and increased interactions between coastal and oceanic habitats. Alpha diversity indices were higher and dominance was lower in oceanic habitats compared to the coastal realm. No marked differences among oceanographically similar areas of the entire archipelago were found. Beta diversity as a measure of similarity among sites or samples revealed variabilities between areas south of Gran Canaria and Fuerteventura islands. A considerable heterogeneity in species distribution was found off SE Fuerteventura in an area with high hydrographic variability. Therefore, both topography and hydrography are important factors influencing the distribution and abundance of pelagic fishes in this oceanic archipelago.
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
Introduction
Apart from continents, shallow seamounts, or oceanic islands, the vast pelagic realm actually lacks any physical structure which could serve as a distribution barrier. Nevertheless, many of its inhabitants have well-defined regions of occurrence, sometimes being limited to relatively small areas. Conditions in the horizontal strata are spatially homogenous with patterns of seasonal and latitudinal variability (Robison Citation2004). Topographic and meso-scale hydrographic features proved to be the most influential for species distribution for islands of volcanic origin, like the Canarian archipelago, in the Eastern Central Atlantic. The special topographic conditions around such islands should intensify the interactions among coastal, oceanic, benthic, and benthopelagic organisms (Uiblein & Bordes Citation1999). This hypothesis was supported by Bordes et al. (Citation1999), who reported the occurrence of two typical shelf-dwelling species (Scomber colias Gmelin, 1789 and Sardinella aurita Valenciennes, 1847) in oceanic waters together with mesopelagic fishes (e.g. Lampanyctus alatus Goode & Bean, 1896, Lobianchia dofleini (Zugmayer, 1911)), and Wienerroither (Citation2003), who found numerous lanternfishes (e.g. Diaphus dumerilii (Bleeker, 1856), Hygophum hygomii (Luetken, 1892), Lobianchia dofleini) in coastal waters.
In addition, the Canarian archipelago shows a particular hydrology. The islands lie in the southward-directed Canary Current and are also under the influence of deep-water masses from the North Atlantic, the Mediterranean, and the Antarctic (Mittelstaedt Citation1983). Topologically induced deep water meanders (Longhurst Citation1998) and eddies (Arístegui et al. Citation1994, Citation1997) as well as seasonally varying upwellings (Uiblein et al. Citation1998), offshore water filaments, eddies, and island wakes (Barton et al. Citation1998) lead to a high complexity and small-scale hydrographic variability both horizontally and vertically. These influence chlorophyll values and primary production and hence the distribution of fishes.
The structure of mesopelagic micronekton assemblages has been the subject of several investigations (Hulley Citation1992; González & Sánchez Citation2002; Fock et al. Citation2004; Pusch et al. Citation2004a), revealing a close relationship to physical and trophic conditions. The decreasing water depth around seamounts reduces density and diversity of mesopelagic fish (Pusch et al. Citation2004b), which respond in different ways to the impingement with the demersal fauna (Porteiro & Sutton Citation2007). Auster et al. (Citation1992) ascribe the aggregation behaviour of myctophid fishes with other pelagic fauna to predation risk reduction and more profitable search for prey. In a study using acoustics and a deployed camera for in-situ identification of pelagic fauna off Hawaiian Islands, Benoit-Bird & Au (Citation2006) observed a highly dynamic use of adjacent oceanic and coastal habitats by mesopelagic fishes. They ascribed their findings to horizontal migrations of certain faunal groups including myctophid fishes, and found in an earlier study (Benoit-Bird & Au Citation2003) large differences in the overall distribution patterns and densities of mesopelagic animals, but did not investigate spatial distribution, assemblage structure, and diversity patterns at high taxonomic resolution. The understanding of the relationship between species richness and its spatial change is fundamental to the assessment of spatial diversity patterns (Koleff & Gaston Citation2002). To our knowledge few studies have been undertaken so far to thoroughly understand the shallow and deep pelagic fish assemblage structure of oceanic islands.
In the present study we further examine earlier hypotheses (see Bordes et al. Citation1999; Uiblein & Bordes Citation1999; Wienerroither Citation2003) that the narrow shelf and the steep slope around oceanic islands facilitate the use of the productive coastal zone by vertically migrating mesopelagic fishes and the increased abundance of the coastal fauna in the epipelagic oceanic realm. The objective of this study is to identify the species composition and distribution of pelagic fishes in the immediate surroundings of the Canary Islands in order to determine (1) overall and local diversity, and (2) the faunal change between individual stations, different habitats, and areas within the islands. The influence of sampling depth, bottom depth, and time of day as important ecological factors on species composition, distribution, and diversity are also discussed.
Material and methods
Six pelagic trawling surveys with the vessel B/E ‘La Bocaina’ were carried out within the Canarian Archipelago between 1997 and 2002 (month and year follow the acronym for each cruise): ‘La Bocaina 11/97’ (B), ‘ECOS 04/99’ (C), ‘Mesopelagic 05/99’ (D), ‘Pelagic 01/00’ (E), ‘Pelagic 11/00’ (F), and ‘Bocaina 03/02’ (G). The trawl tows were conducted horizontally during day and night, at different depths (epipelagic: 8–219 m; mesopelagic: 215–1009 m) and above different bottom depths from coastal (min. bottom depth 40 m) to oceanic (max. bottom depth 2525 m) waters. Exact data concerning duration, location, trawling and bottom depth, of all surveys and tows are given in . The maps of the surveys and the single sampling stations, indicating the type (coastal, epipelagic or mesopelagic), the length, and the direction of the trawls are shown in and . Trawl tows overlapping the 200 m depth line which demarcates the epi- and mesopelagic realm were considered as epipelagic. Trawl tows overlapping the 200 m bottom depth line demarcating the coastal and oceanic realm were considered as coastal if the range of the bottom depth during the haul was less than 200 m and as epipelagic if the range was more than 200 m.
Figure 1. Map of survey (a) ‘La Bocaina 11/97’ (B1–B18), and (b) ECOS 04/99’ (C1–C23) and ‘Mesopelagic 05/99’ (D1–D6) (legend as in a). Station numbers refer to . Insert in a: the Canarian Archipelago, situated in the Eastern Central Atlantic.
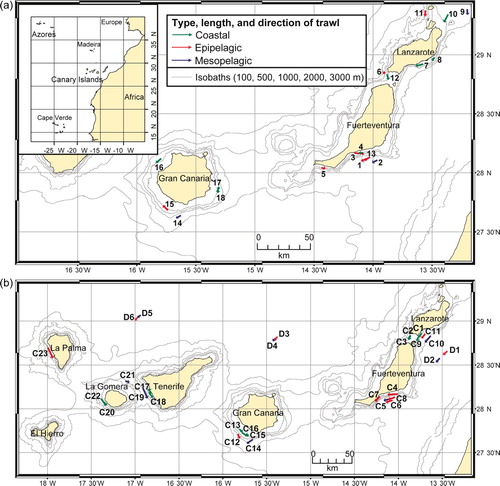
Figure 2. Map of survey (a) ‘Pelagic 01/00’ (E1–E11) and ‘Pelagic 11/00’ (F1–F19) (legend as in b) and (b) ‘Bocaina 03/02’ (G1–G27). Station numbers refer to .
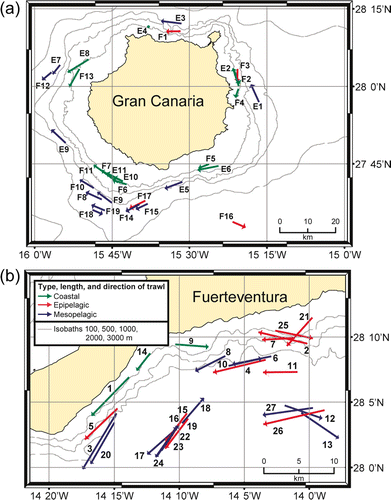
Table I. Characteristics of trawl tow stations of all surveys. Epipelagic stations in normal, coastal stations in italics, and mesopelagic stations in bold letters. Stations B4, B7, B10, C1, C3, C11, C20, E4, F4, G9, and G11 were without result. See text for further explanations.
A commercial trawl with 80 m maximum horizontal opening was used (average vertical opening during tows 6.6 to 16.5 m). Cod-end mesh size was 2 mm and the opening of the cod-end was strengthened by a steel ring of 1.5 m diameter. Samples were scaled to a trawling time of 1 h. Characteristics of the vessel and the net, as well as a description of the fishing operations are given in Bordes et al. (Citation1999) and Wienerroither (Citation2003). The fishes were fixed in 7% formalin and later transferred to 70% ethanol. The specimens of the surveys ‘La Bocaina 11/97’ and ‘ECOS 04/99’ have been included into the collections of the Zoological Museum of the University of Copenhagen (ZMUC), Denmark, and of the Instituto Canario de Ciencias Marinas (ICCM), Telde, Gran Canaria, Spain. Specimens of the remaining surveys are all deposited at ICCM. Species identification is based on the following literature: Badcock (Citation1982), Bigelow et al. (Citation1964), Hulley (Citation1981), Johnson (Citation1974), Krefft (Citation1970, Citation1971), Nafpaktitis et al. (Citation1977), Nakamura & Parin (Citation1993), Nelson (Citation2006), Nielsen & Smith (Citation1978), Nielsen & Bertelsen (Citation1985), Rofen (Citation1966), and Whitehead et al. (Citation1984–1986). The occurrence of species in Canarian waters and general distribution have been checked using Badcock (Citation1970), Badcock & Merrett (Citation1976), Bordes et al. (Citation1999), Brito et al. (Citation2002), Hureau & Monod (Citation1979), Kotthaus (Citation1972), Lloris et al. (Citation1991), Quéro et al. (Citation1990), Rodríguez (Citation2000), Rudyakov (Citation1979), Whitehead et al. (Citation1984–1986), and FishBase (http://www.fishbase.org).
Multivariate community analysis
Epipelagic trawl stations conducted during the day (i.e. G15 and G16) as well as singletons with respect to the whole data set (station B8) were excluded a priori. Hierarchical agglomerative clustering with group-average linking, based on the Bray–Curtis Similarity measure, was used to delineate groups with distinct community structure (Field et al. Citation1982). The normal or q-type analysis uses all species and treats samples as individual observations, each being composed of a number of attributes (i.e. the various taxa collected per tow). The analysis was done based on root–root transformed densities using the PC-ORD software package. The transformation down-weights the more abundant species and is invariant to scale range (Field et al. Citation1982). The specification of the similarity level for cluster separation was chosen to be data-driven. To compensate for possible misinterpretations derived from the exclusive use of dendrograms in community studies (Field et al. Citation1982), non-metric multidimensional scaling (MDS) was adopted. This method produces an ordination of the stations or species in two dimensions. Indicator species values were calculated using the method of Dufrene & Legendre (Citation1997) to detect and describe the value of different species for the clusters derived by the classification and ordination method (McCune & Grace Citation2002). Information on the concentration of species abundance in a particular group is combined with the fidelity of occurrence of a species in a particular group, whereupon perfect indication means that the presence of a species points to a particular group without error, at least with the data set at hand. The statistical significance is evaluated by a randomization test (Monte Carlo Test, 1000 runs) which allows estimating exact significance without relying on the assumptions required for the standard asymptotic method.
Ecological diversity
The diversity and abundance of species was analysed using a representative set of indices (Magurran Citation2004) including total number of species and total number of individuals, species richness (Shannon–Wiener H’ and Simpsons D Index), evenness (Equitability J), and dominance (Berger–Parker). The hypotheses were tested that there are differences in diversity among (1) the principal clusters obtained from the multivariate analysis (result-driven data selection) and (2) areas within the Canarian archipelago (a-priori data selection). For comparison among areas epi- and mesopelagic tows south of Gran Canaria (epipelagic stations: B15, C12, F 17; mesopelagic stations: B14, C14, E5, F8, F9, F10, F14, F15, F18, F19) and south of Fuerteventura (epipelagic stations: B1, B3, B13, C4, C5, C6, C7, C8, G2, G4, G5, G7, G15, G16, G19, G21, G25, G26; mesopelagic stations: B2, G3, G6, G8, G10, G12, G13, G17, G18, G20, G22, G23, G24, G27), were grouped. Mean and confidence intervals for indices of clusters and areas were calculated and pairwise t-tests were used for additional statistical examination.
Beta diversity investigates the degree of association or similarity of sites or samples and is higher, the fewer species the different communities share (Magurran Citation2004). It was used to find possible differences (1) between the areas south of Gran Canaria and Fuerteventura and (2) variations among three selected groups of mesopelagic tows from survey 03/02 off SE Fuerteventura (b; group 1: G6, G8, G10; group 2: G22, G23, G24; group 3: G12, G13; G27) to study small-scale discontinuities. Two indices, recorded to be the best to determine the degree of turnover in species composition, Whittaker βW and Wilson & Shmida βT (Wilson & Shmida Citation1984; Magurran Citation2004), were used based on presence and absence data.
Results
From a total of 104 trawl tows (35 coastal, 32 epipelagic and 37 mesopelagic), 93 resulted in the capture of 65,815 adult or juvenile fishes (14,609 epipelagic, and 51,206 meso-, bathy-, benthopelagic, or bathydemersal fishes). These belong to 19 orders, 51 families, 123 genera, and 211 species. Thirteen species are new records for the area. An overview of all species, indicating number of individuals and the survey in which they were captured is provided in .
Table II. List of species encountered in the six surveys, with indication of number of specimens per survey. New records for the area are emphasized in bold, systematic order is according to Nelson (2006).
Multivariate community analysis
The dendrogram resulting from the Bray–Curtis analysis of the 90 trawl stations is shown in . Most of the stations are allocatable to four main clusters (1–4). One smaller cluster (cluster 5) is formed by four trawl stations. Two stations deviate in species composition from all others. The results of multidimensional scaling (MDS) using the same similarity matrix as above, delineating groups of stations from the dendrogram is shown in . This analysis gives essentially the same picture as the dendrogram. The principal factor of association was the trawling depth in combination with the bottom depth. Mesopelagic tows are found in cluster 2 and the smaller cluster 5, whereas coastal tows are represented in cluster 3 and 4. Epipelagic oceanic tows formed the cluster 1 and are distributed in all other clusters except cluster 5. On account of the many stations no fixed similarity level for cluster separation was defined, therefore clusters vary from 45.8% (cluster 2) to 14.2% (cluster 4) similarity. illustrates epipelagic and coastal tows around Gran Canaria and Fuerteventura and their affiliation to the clusters, showing also the spatial overlap between cluster 3 and 4. Below is a detailed characterization of each of the five clusters.
-
Cluster 1, similarity level 41.1%, 13 stations, mean water depth 1427 m, habitat epipelagic–oceanic: it comprises only epipelagic tows, characterized by a high variability in abundance of vertically migrating mesopelagic species and by three epipelagic species (Sardinella aurita, Scomber colias, Trachurus picturatus). Indicator species for this group are myctophids (), known for their intense daily vertical migration behaviour. All tows are restricted to the oceanic realm ().
-
Cluster 2, similarity level 45.8%, 33 stations, mean water depth 1388 m, habitat mesopelagic: with the exception of one far-offshore epipelagic trawl station (D6), cluster 2 is composed of mesopelagic tows exclusively. The relatively high similarity (32.3%) of clusters 1 and 2 indicates a close relationship among the stations of these groups. In fact they share many vertically migrating species, but not the non- or little migratory species (found only in the mesopelagic tows), which delineate the two clusters. MDS () plots the stations of clusters 1 and 2 very closely, but also indicates the high similarity of the stations within and between the clusters. Non-migrating species like Serrivomer beanii and Sigmops elongatus, among others, are the best indicators for this group ().
-
Cluster 3, similarity level 17.0%, 12 stations, mean water depth 327 m, habitat coastal: this cluster comprises six coastal and six epipelagic samples, composed of several epipelagic species, mainly in low numbers, and vertically migrating myctophids, partly rich in individuals. Diaphus dumerilii is the best indicator species. The mesopelagic species in this cluster obviously perform an intense horizontal migration from the oceanic realm towards the islands ().
-
Cluster 4, similarity level 14.2%, 26 stations, mean water depth 205 m, habitat coastal: the cluster includes most of the tows taken above the continental shelf (coastal) plus eight oceanic epipelagic tows. It is characterized by the highest number of epipelagic species and individuals. As opposed to cluster 3, mesopelagic fishes (in terms of species and individuals) were less frequent in these trawl stations. Among the epipelagic species caught during the six surveys, only Scomber colias and Trachurus picturatus showed up regularly and are indicator species.
-
Cluster 5, similarity level 44.3%, 4 stations, mean water depth 1294 m, habitat mesopelagic: the mesopelagic tows in this small cluster have a strong affiliation to cluster 2, the stations are partly overlapping (), but differ in a slightly lower number of species.
Figure 3. Dendrogram showing classification of the 90 trawl stations, four main clusters (1–4) can be distinguished. Abundances of species were root–root transformed before comparing stations using the Bray–Curtis measure, and the dendrogram formed by group-average sorting. The distance axis indicates the distance between groups, not expressed as a simple distance measure, but as Wishart's (Citation1969) Objective Function (which is a measure of information loss as agglomeration proceeds): Cluster 1 = 6.95, Cluster 2 = 6.40, Cluster 3 = 9.80, Cluster 4 = 10.13. The second axis is based on the same function, but is converted to a percentage of information remaining: Cluster 1 = 41.1%, Cluster 2 = 45.8%, Cluster 3 = 17.0%, Cluster 4 = 14.2%.
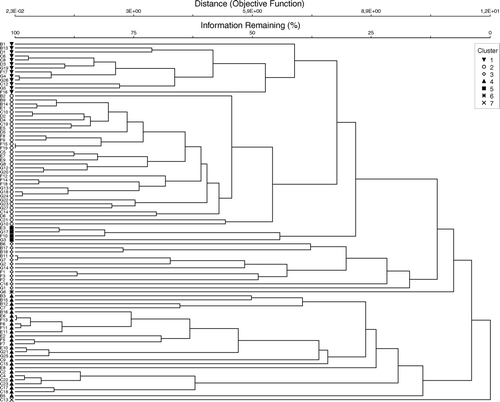
Figure 4. Ordination of the 90 trawl stations in two dimensions using multidimensional scaling on the same similarity matrix as , final stress = 14.99250.
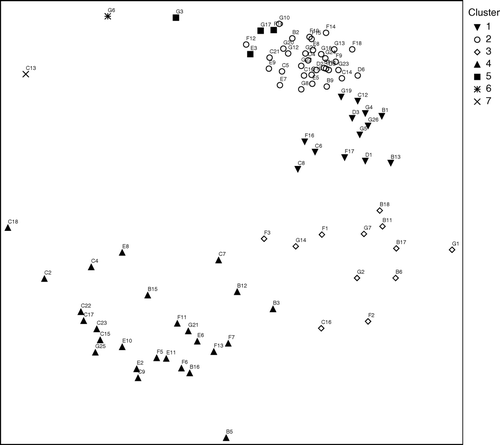
Figure 5. Epipelagic and oceanic tows around Gran Canaria and Fuerteventura, indicating their affiliation to clusters in classification () and ordination ().
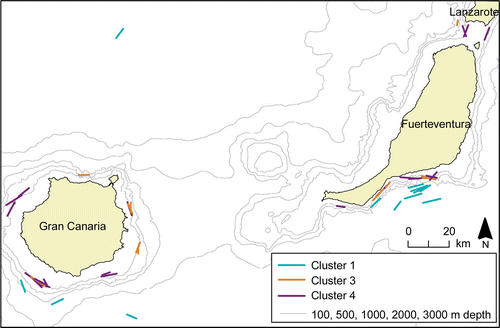
Table III. Indicator species for the four clusters (I–IV) derived by the classification and ordination method. Relative abundance (RA) of the species in the group of stations over the average abundance of that species in all stations (in % of perfect indication). Relative frequency (RF) of the species in the trawl stations of the group (in % of perfect indication). Indicator values (IV) range from 0 (no indication) to 100 (perfect indication) and are based on a combination of RA and RF. Monte Carlo test of significance of observed indicator value for the species in the particular group. Only species with an indicator value of more than 50 are listed.
Ecological diversity
Alpha diversity
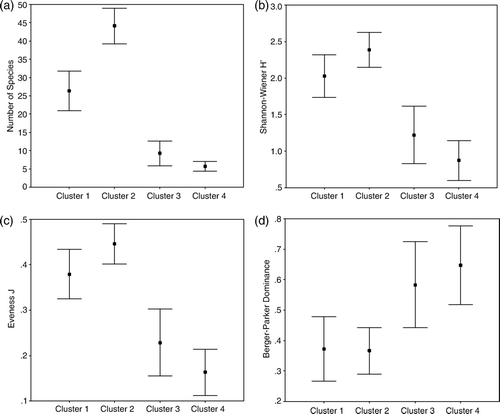
Only the four main clusters (1–4) derived by classification and ordination methods are considered in the diversity analysis (Figure 6). Clusters formed by stations close to the shelf (3 and 4) tend to be different from clustered stations from oceanic waters (1 and 2). Cluster 2 (mesopelagic tows) has the highest diversity and equitability and the lowest dominance. This is in contrast to cluster 4, which is much less diverse but shows a high level of dominance (with cluster 3).
The number of species (a) is significantly different between all clusters (F = 153.36, p<0.001) except between clusters 3 and 4, whereas there are no significant differences in number of individuals between the clusters. Significant differences in diversity indices (H’ between clusters 1–3, 1–4, 2–3, and 2–4, F = 56.12, p<0.001; Simpson D between clusters 1–4, 2–3, and 2–4, F = 17.02, p<0.005), equitability (Evenness J between clusters 1–3, 1–4, 2–3, and 2–4, F = 56.14, p<0.001) and dominance (Berger–Parker between clusters 1–4, 2–3, and 2–4, F = 14.38, p<0.005) among the clusters were revealed by pairwise t-tests, which coincide with non-overlapping 99% confidence intervals (b–d).
Figure 6. Mean and 99% confidence intervals of diversity and evenness indices (a: Number of species, b: Shannon–Wiener H’, c: Evenness J, d: Berger–Parker Dominance) of the principal clusters (cf. classification and ordination).
Examinations based on a-priori data selection to detect differences between richness, diversity, equitability, and dominance between the patches of trawl stations to the south of Gran Canaria and Fuerteventura, epi- and mesopelagic tows were treated separately. Pairwise t-tests revealed no significant differences.
Beta diversity
To the south of Fuerteventura beta diversity is considerably higher compared to the south of Gran Canaria (). The two habitats to the south of Gran Canaria have approximately similar beta diversity, whereas there is a remarkable difference between the epi- and mesopelagic realms south of Fuerteventura. Groups 2 and 3 have similar beta diversity, whereas group 1 (closer to the coast and the upwelling region off Gran Tarajal) is higher in βW and βT ().
Table IV. Beta diversity, Whittaker βW (=S/α) and Wilson & Shmida β T (=[g(H) + l(H)]/2S j ), of two different areas, habitats, and groups of mesopelagic trawl stations in the south of Fuerteventura. (S=total number of species recorded, α=average sample diversity, g(H) = number of species gained, l(H) = number of species lost, S j =species richness of sample j).
Discussion
Multivariate community analysis
The dense ordination of mesopelagic trawl stations (cluster 2) reveals that these have the most closely related species composition. Although several tows contain very rare and unique species, the tows are primarily unified by the dominant non- or little migrating fishes (e.g. Serrivomer spp., Cyclothone spp.). The far offshore position and the deep sounding of the only epipelagic station within this cluster might contribute to the species composition resembling the mesopelagic tows.
Clusters 1 and 2 showed the closest relationships. Their tows were all made in the oceanic realm and contain a high diversity of myctophids, which is their unifying factor. Lepidophanes gaussi is the main indicator species for cluster 1 and all the other indicator species are lanternfishes too (e.g. Benthosema suborbitale, Ceratoscopelus warminigii, Diogenichthys atlanticus). These inhabit the epipelagic realm during night for foraging reasons and are often encountered at higher densities there than in mesopelagic tows. Possible explanations for this phenomenon might be the limited space in near-surface habitats leading to a denser aggregation of specimens and/or biotic or abiotic factors with effects on spatial preferences. The most abundant species in these clusters are all mesopelagic (mainly myctophids), only the epipelagic Scomber colias was encountered in comparable abundances. This confirms the unusual horizontal migration into oceanic waters of this normally shelf-dwelling species, as indicated by Bordes et al. (Citation1999).
Myctophids are also the most influential group in cluster 3. Although Diaphus dumerilii is the best indicator species, other lanternfish species occur in high numbers in trawl tows of this cluster too (e.g. Hygophum hygomii, Lobianchia dofleini, Notoscopelus resplendens). But the latter are also very abundant in other tows and therefore less appropriate as indicators. Diaphus dumerilii shows its highest abundances in rather shallow depths indicating a certain degree of land association. Cluster 3 combines trawls in and close to the coastal zone. Uiblein & Bordes (Citation1999) ascribe the occurrence of mesopelagic species in such shallow waters to the abrupt changeover of depths around islands of volcanic origin. Nevertheless, the findings of such high numbers (e.g. 1561 D. dumerilii in survey 11/00 tow 3, 43–90 m trawling depth; 888 H. hygomii in survey 11/00 tow 1, 55–90 m trawling depth), differ from the observations by Hulley (Citation1986) and Hulley & Prosch (Citation1987), who recorded in the southern Benguela upwelling region the stock of only one myctophid species above a bottom depth of less than 300 m. Several species of Diaphus have been found to show spatial affinities to steep slope bottoms and hence characterized as ‘pseudoceanic’ (Merrett Citation1985; Hulley & Lutjeharms Citation1989).
Cluster 4 shows the highest heterogeneity, deriving from several epipelagic species, especially the two indicator species Scomber colias and Trachurus picturatus. Surprising is the presence of the myctophid D. dumerilii in this cluster comprising mostly coastal tows. In tows 5, 6, and 7 of survey 11/00, the abundance of the lanternfish was comparatively high (54, 128, and 1075 individuals) and also Boops boops (393 and 271 individuals in tow 5 and 7) and S. colias (387 individuals in tow 5; all numbers extrapolated) showed relatively high numbers. These three trawl stations were typically coastal, although this habitat is naturally very narrow around islands of volcanic origin. As proposed by Uiblein & Bordes (Citation1999), close interactions among coastal, oceanic, benthic, and benthopelagic organisms are induced by the abrupt topography with possible predation of epipelagic species on myctophids like D. dumerilii. Impingement of pseudoceanic species may lead to intense spatial and trophic interactions with slope community (Reid et al. Citation1991; Gordon et al. Citation1995; Uiblein & Bordes 1999). The more dispersed ordination of the stations of group IV compared to the other large clusters () reflects irregular occurrences of many species.
Clusters 1 and 2 are clearly defined by habitat (oceanic–epipelagic and mesopelagic), whereas there are spatial overlaps between tows affiliated to clusters 3 and 4 (). Seasonal effects can be excluded, as coastal tows within surveys show affiliation to different clusters. This suggests that occurrence and abundance of species is locally and temporally limited to a rather small scale instead of showing a constant pattern throughout the study area. The indicator species are not necessarily restricted to a single cluster (). Specifically D. dumerilii and S. colias were found in stations of all clusters (except S. colias in cluster 2). Although numbers of individuals vary somewhat significantly, this clearly shows an intensified use of coastal habitats by mesopelagic fishes as well as of oceanic habitats by coastal species, respectively.
Several authors used a similar approach to investigate mesopelagic nekton assemblages. Hulley & Lutjeharms (Citation1989), for instance, found that in the southern Benguela region the depth of the water column was the reason for a change in the lanternfish fauna and distinguish between oceanic and pseudoceanic species. Depth is also the deciding factor in cephalopod assemblages along the Spanish Mediterranean coast (González & Sánchez Citation2002). Fock et al. (Citation2004) indicate that habitat structure in conjunction with physical and biological features is an important determinant of community structure. Benoit-Bird & Au (Citation2006) found myctophids as the only animals in shallow waters of Hawaiian Islands, migrating actively with a striking horizontal component. Furthermore, these myctophids were of larger size than those found in farther offshore, deeper layers, which they ascribed to differences in swimming speed.
Ecological diversity
Alpha diversity
The diversity indices used confirm the often-stated characteristics of the mesopelagic realm, reported to be the most diverse and densely inhabited area of the open oceans (National Research Council Citation1995). The high species richness and evenness, and the low dominance values obtained in the current study agree with this statement. Quite contrasting are the mainly coastal tows of cluster 4. These stations in shallow bottom depths are characterised by low species richness, an often high number of individuals (and biomass) and one or two dominating epipelagic species. Clusters 1 and 3 are positioned in between. Cluster 1 (epipelagic tows) is in its diversity closer to cluster 2, with high richness and evenness but low dominance. This can be ascribed to the vertical migrants represented in these tows by high numbers (in species and individuals) and the lack of the partly rare non-migrators. Cluster 3 resembles cluster 4, although these tows are a special case: made close to the coast (some of them even coastal), with mesopelagic fishes, remarkably high in individuals but low in species.
Beta diversity
Depths of more than 2000 m are encountered close to the investigation area of Gran Canaria, whereas this depth zone is farther away from that of Fuerteventura. The differences in diversity might reflect the prevailing depth zones in the respective areas or general differences in spatial heterogeneity. The higher oceanic influence and the eddy south of Gran Canaria (Arístegui et al. Citation1994, Citation1997; Bordes et al. Citation1999) may have an effect on species distribution. Arístegui et al. (Citation1989) found high primary production and large populations of mesozooplankton in coastal waters south of Gran Canaria compared to oceanic waters. Barange et al. (Citation1998) emphasized the intense influences of hydrological features on a pelagic community in general. Even more remarkable is the distinctness between the epi- and mesopelagic habitat off south Fuerteventura. It emphasizes the complex connection of hydrological features and species distribution. Uiblein et al. (Citation1996, Citation1998) discussed the existence of a seasonally limited local upwelling off Gran Tarajal, southeast Fuerteventura. Within the sub-area groupings of mesopelagic tows the two groups farther away from the coast have similar beta diversity, that clearly differs from group 1 indicating a heterogeneous species distribution at the scale of a few kilometres distance only. The mesopelagic boundary community of Hawaiian Islands shows similar regional differences in taxonomic composition, abundance and diversity (Reid et al. Citation1991; Benoit-Bird et al. Citation2001; Lammers et al. Citation2004). Benoit-Bird & Au (Citation2006) refer the fine scale separation of habitat use to a reduction of competition between individuals in these dense aggregations relative to night time. Heterogeneity in species distribution depending on topography and hydrography seems to be a common feature, as it was also detected between adjacent deep-sea canyons off the Georges Bank, NW Atlantic (Uiblein et al. unpublished data).
This study has resulted in the collection of 13 new pelagic fish species for the Canary Islands. Not all islands and habitats have been covered by the surveys. We have presented here clear evidence of considerable variation in local species composition reflecting the influences of topography and hydrography and the interactions between these two variables. Both epipelagic and mesopelagic fishes seem to respond actively to physical heterogeneity at the scale of a few kilometres only, thus inducing patchiness and turnover among close-by areas. The high number of species encountered, the still ongoing discovery of hitherto unknown species, and the finding of distinct, taxon-specific distribution patterns strongly suggest that pelagic fishes are an important component for the assessment and functional understanding of biodiversity around oceanic islands.
Editorial responsibility: Peter R. Møller
Acknowledgements
All surveys were carried out in projects supported by the Viceconsejería de Pesca del Gobierno de Canarias. Rupert Wienerroither got financial support from the ‘Stiftungs- und Förderungsgesellschaft der Paris-Lodron Universität Salzburg’. Sincere thanks to all colleagues at the Zoological Museum Copenhagen (ZMUC) and the University of Salzburg, especially A. Lametschwandtner and J. Strobl.
Notes
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
References
- Arístegui , J , Hernández-León , S , Gómez , M , Medina , L , Ojeda , A and Torres , S. 1989 . Influence of the north trade winds on the biomass and production of neritic plankton around Gran Canaria island. Topics in Marine Biology . Scientia Marina , 53 : 223 – 9 .
- Arístegui , J , Sangrá , P , Hernández-León , S , Cantòn , M , Hernández-Guerra , A and Kerling , LJ. 1994 . Island-induced eddies in the Canary Islands . Deep-Sea Research I , 41 : 1509 – 25 .
- Arístegui , J , Tett , P , Hernández-Guerra , A , Basterretxea , G , Montero , MF Wild , K . 1997 . The influence of island-generated eddies on chlorophyll distribution: A study of mesoscale variation around Gran Canaria . Deep-Sea Research I , 44 : 71 – 96 .
- Auster , PJ , Griswold , CA , Youngbluth , MJ and Bailey , TG. 1992 . Aggregations of myctophid fishes with other pelagic fauna . Environmental Biology of Fishes , 35 : 133 – 9 .
- Badcock , J. 1970 . The vertical distribution of mesopelagic fishes collected on the SOND cruise . Journal of the Marine Biological Association of the UK , 50 : 1001 – 44 .
- Badcock , J. 1982 . A new species of the deep-sea fish genus Cyclothone Goode & Bean (Stomiatoidei, Gonostomatidae) from the tropical Atlantic . Journal of Fish Biology , 20 : 197 – 211 .
- Badcock , J and Merrett , NR. 1976 . Midwater fishes in the eastern North Atlantic – I. Vertical distribution and associated biology in 30°N, 23°W, with developmental notes on certain myctophids . Progress in Oceanography , 7 : 3 – 58 .
- Barange , M , Pakhomov , EA , Perissinotto , R , Froneman , PW , Verheye , HM Taunton-Clark , J . 1998 . Pelagic community structure of the subtropical convergence region south of Africa and in the mid-Atlantic Ocean . Deep-Sea Research I , 45 : 1663 – 87 .
- Barton , ED , Arístegui , J , Tett , P , Cantón , M , García-Braun , J Hernández-León , S . 1998 . The transition zone of the Canary Current upwelling region . Progress in Oceanography , 41 : 455 – 504 .
- Benoit-Bird , KJ and Au , WWL. 2003 . Spatial dynamics of a nearshore, micronekton sound-scattering layer . ICES Journal of Marine Science , 60 : 899 – 913 .
- Benoit-Bird , KJ and Au , WWL. 2006 . Extreme diel horizontal migrations by a tropical nearshore resident micronekton community . Marine Ecology Progress Series , 319 : 1 – 14 .
- Benoit-Bird , KJ , Au , WWL , Brainard , RE and Lammers , MO. 2001 . Diel horizontal migration of the Hawaiian mesopelagic boundary community observerd acoustically . Marine Ecology Progress Series , 217 : 1 – 14 .
- Bigelow HB , Cohen DM , Dick MM , Gibbs Jr. RH , Grey M , Morrow Jr. JE , et al. . 1964 . Fishes of the Western North Atlantic. Part IV. Soft-rayed Bony Fishes . New Haven, CT : Sears Foundation of Marine Research, Yale University .
- Bordes , F , Uiblein , F , Castillo , R , Barrera , A , Castro , JJ Coca , J . 1999 . Epi- and mesopelagic fishes, acoustic data, and SST images collected off Lanzarote, Fuerteventura and Gran Canaria, Canary Islands, during cruise ‘La Bocaina 04-97’ . Informes Técnicos del Instituto Canario de Ciencias Marinas , 5 : 1 – 45 .
- Brito , A , Pascual , PJ , Falcon , JM , Sancho , A and Gonzalez , G. 2002 . Pesces de las Islas Canarias. Catálogo comentado e ilustrado , Tenerife : Francisco Lemus .
- Dufrene , M and Legendre , P. 1997 . Species assemblages and indicator species: The need for a flexible asymmetrical approach . Ecological Monographs , 67 : 345 – 66 .
- Field , JG , Clarke , KR and Warwick , RM. 1982 . A practical strategy for analysing multispecies distribution patterns . Marine Ecology Progress Series , 8 : 37 – 52 .
- Fock , HO , Pusch , C and Ehrich , S. 2004 . Structure of deep-sea pelagic fish assemblages in relation to the Mid-Atlantic Ridge (45°–50°N) . Deep-Sea Research I , 51 : 953 – 78 .
- González , M and Sánchez , P. 2002 . Cephalopod assemblages caught by trawling along the Iberian Peninsula Mediterranean coast . Scientia Marina , 66 : 199 – 208 .
- Gordon , JDM , Merrett , NR and Haedrich , RL. 1995 . “ Environmental and biological aspects of slope dwelling fishes ” . In Deep-water Fisheries of the North Atlantic Oceanic Slope , Edited by: Hopper , AG . 1 – 30 . Dordrecht : Kluwer Academic Publishers .
- Hulley , PA. 1981 . Results of the research cruises of FRV ‘Walther Herwig’ to South America. LVIII. Family Myctophidae (Osteichthyes, Myctophiformes) . Archiv für Fischereiwissenschaften , 31 : 1 – 300 .
- Hulley , PA. 1986 . Lanternfishes of the southern Benguela region, Part 1, Faunal complexity and distribution . Annals of the South African Museum , 97 : 227 – 49 .
- Hulley , PA. 1992 . Upper-slope distribution of oceanic lanternfishes (family: Myctophidae) . Marine Biology , 114 : 365 – 83 .
- Hulley , PA and Lutjeharms , JRE. 1989 . Lanternfishes of the southern Bengula region. Part 3. The pseudoceanic–oceanic interface . Annals of the South African Museum , 98 : 409 – 35 .
- Hulley , PA and Prosch , RM. 1987 . Mesopelagic fish derivatives in the southern Benguela upwelling region. The Benguela and comparable ecosystems . South African Journal of Marine Science , 5 : 597 – 611 .
- Hureau JC , Monod T . 1979 . Check-list of the fishes of the North-eastern Atlantic and of the Mediterranean/Catalogue des poissons de l'Atlantique du nord-est et de la Méditerranée (CLOFNAM) . 2nd edition . Paris : UNESCO .
- Johnson , RK. 1974 . Five new species and a new genus of alepisauroid fishes of the Scopelarchidae (Pisces: Myctophiformes) . Copeia , 1974 : 449 – 57 .
- Koleff , P and Gaston , KJ. 2002 . The relationship between local and regional species richness and spatial turnover . Global Ecology and Biogeography , 11 : 363 – 75 .
- Kotthaus , A. 1972 . Die meso- und bathypelagischen Fische der “Meteor”-Roßbreiten-Expedition 1970 (2. und 3. Fahrabschnitt) . Meteor Forschungsergebnisse Reihe D Biologie , 11 : 1 – 28 .
- Krefft , G. 1970 . Ergebnisse der Forschungsreisen des FFS “Walther Herwig” nach Südamerika. XII. Barbantus elongatus spec. nov. (Pisces, Alepocephaloidei), ein weiterer neuer Searside aus dem tropischen Atlantik . Archiv für Fischereiwissenschaften , 1970 : 22 – 7 .
- Krefft , G. 1971 . Ergebnisse der Forschungsreisen des FFS “Walther Herwig“ nach Südamerika. XVIII. Pseudosopelus scutatus spec. nov. (Pisces, Perciformes, Trachinoidei, Chiasmodontidae), ein neuer Kreuzzahnbarsch aus dem tropischen Atlantik . Archiv für Fischereiwisssenschften , 1971 : 165 – 74 .
- Lammers , MO , Brainard , RE and Au , WWL. 2004 . Diurnal trends in the mid-water biomass community of the Northwestern Hawaiian Islands observed acoustically . Journal of the Acoustical Society of America , 116 : 2488 – 9 .
- Lloris , D , Rucabado , J and Figueroa , H. 1991 . Biogeography of the Macaronesian Ichthyofauna (The Azores, Madeira, the Canary Islands, Cape Verde and the African Enclave) . Boletim do Museo Municipal do Funchal , 43 : 191 – 241 .
- Longhurst , A. 1998 . Ecological geography of the sea , San Diego, CA : Academic Press .
- Magurran , AE. 2004 . Measuring Biological Diversity , Oxford/Carlton : Blackwell Science Ltd .
- McCune , B and Grace , JB. 2002 . Analysis of Ecological Communities , Gleneden Beach, OR : MjM Software Design .
- Merrett NR . 1985 . Biogeography and the oceanic rim: A poorly known zone of ichthyofaunal interaction . In : Pelagic biogeography. UNESCO technical papers in marine science 49 : 201 – 9 .
- Mittelstaedt , E. 1983 . The upwelling area off northwest Africa – a description of phenomena related to coastal upwelling . Progress in Oceanography , 12 : 307 – 31 .
- Nafpaktitis BG , Backus RH , Craddock JE , Haedrich RL , Robison BH , Karnella C . 1977 . Family Myctophidae . In : Fishes of the Western North Atlantic. Part VII. Order Iniomi (Myctophiformes) . New Haven, CT : Sears Foundation for Marine Research, Yale University , p. 13 – 265 .
- Nakamura I , Parin NV . 1993 . FAO Species Catalogue. Vol. 15, Snake mackarels and cutlessfishes of the world (Families Gempylidae and Trichiuridae) . Rome : FAO Fisheries Synopsis .
- National Research Council . 1995 . Understanding Marine Biodiversity . Washington, DC : National Academic Press .
- Nelson JS . 2006 . Fishes of the World . 4th edition . New York : John Wiley & Sons Inc .
- Nielsen , JG and Bertelsen , E. 1985 . The gulper-eel family Saccopharyngidae (Pisces, Anguilliformes) . Steenstrupia , 11 : 157 – 206 .
- Nielsen , JG and Smith , DG. 1978 . The eel family Nemichthyidae (Pisces, Anguilliformes) . Dana-Report , 88 : 1 – 71 .
- Porteiro FM , Sutton T . 2007 . Midwater fish assemblages and seamounts . In: Pitcher TJ , Morato T , Hart PJB , Clark MR , Haggan N , Santos RS Seamounts: Ecology, Fisheries and Conservation . Fish and Aquatic Resources Series 12 : 101 – 16 .
- Pusch , C , Beckmann , A , Porteiro , FM and von Westernhagen , H. 2004b . The influence of seamounts on mesopelagic fish communities . Archive of Fishery and Marine Research , 51 : 165 – 86 .
- Pusch , C , Hulley , PA and Kock , K-H. 2004a . Community structure and feeding ecology of mesopelagic fishes in the slope waters of King George Island (South Shetland Islands, Antarctica) . Deep-Sea Research I , 51 : 1685 – 708 .
- Quéro JC , Hureau JC , Karrer C , Post A , Saldana L 1990 . Checklist of the fishes of the eastern tropical Atlantic/Catalogue des poissons de l'Atlantique oriental tropical (CLOFETA) . Paris : UNESCO .
- Reid , SB , Hirota , J , Young , RE and Hallacher , LE. 1991 . Mesopelagic-boundary community in Hawaii: Micronekton at the interface between neritic and oceanic ecosystems . Marine Biology , 109 : 427 – 40 .
- Robison , BH. 2004 . Deep pelagic biology . Journal of Experimental Marine Biology and Ecology , 300 : 253 – 72 .
- Rodríguez , JM. 2000 . Fish larvae form the Canary region in autumn . Scientia Marina , 64 : 79 – 85 .
- Rofen RR . 1966 . Family Paralepididae . In : Fishes of the Western North Atlantic. Part V . New Haven, CT : Sears Foundation for Marine Research, Yale University , p. 205 – 561 .
- Rudyakov , YA. 1979 . Diurnal vertical migrations of pelagic animals in the Canary Islands area . Oceanology , 19 : 196 – 9 .
- Uiblein , F and Bordes , F. 1999 . Complex trophic interactions around ocean islands . Ocean Challenge , 9 : 15 – 16 .
- Uiblein , F , Bordes , F and Castillo , R. 1996 . Diversity, abundance and depth distribution of demersal deep-water fishes off Lanzarote and Fuerteventura, Canary Islands . Journal of Fish Biology , 49 : 75 – 90 .
- Uiblein , F , Bordes , F , Castillo , R and Ramos , AG. 1998 . Spatial distribution of shelf- and slope-dwelling fishes collected by bottom longline off Lanzarote and Fuerteventura, Canary Islands . Marine Ecology , 19 : 53 – 66 .
- Whitehead PJP , Bauchot ML , Hureau JC , Nielsen J , Tortenese E . 1984–1986 . Fishes of the North-eastern Atlantic and the Mediterranean/Poissons de l'Atlantique du Nord-Est et de la Méditerranée (FNAM) . Paris : UNESCO .
- Wienerroither , RM. 2003 . Species composition of mesopelagic fishes in the area of the Canary Islands, Eastern Central Atlantic . Informes Técnicos del Instituto Canario de Ciencias Marinas , 9 : 1 – 110 .
- Wilson , MV and Shmida , A. 1984 . Measuring beta diversity with presence/absence data . Journal of Ecology , 72 : 1055 – 64 .
- Wishart , D. 1969 . An algorithm for hierarchical classifications . Biometrics , 25 : 165 – 70 .