Abstract
We show that a discrete-time, two-species competition model with Ricker (exponential) nonlinearities can exhibit multiple mixed-type attractors. By this is meant dynamic scenarios in which there are simultaneously present both coexistence attractors (in which both species are present) and exclusion attractors (in which one species is absent). Recent studies have investigated the inclusion of life-cycle stages in competition models as a casual mechanism for the existence of these kinds of multiple attractors. In this paper we investigate the role of nonlinearities in competition models without life-cycle stages.
1. Introduction
In Citation1 the authors utilize a competition model to explain an unusual coexistence result observed and studied by T. Park and his collaborators in a series of classic experiments involving two species of insects (from the genus Tribolium) Citation2–4. The explanation offered in Citation1 is based on a single species model (called the LPA model) designed explicitly to account for the dynamics of the species involved. The LPA model has an impressive track record, spanning several decades, of describing and predicting the dynamics of Tribolium populations, under a variety of circumstances in controlled laboratory experiments—dynamics that range from equilibrium and periodic cycles to quasi-periodic and chaotic attractors Citation5, Citation6. This history of success adds credence to the two-species competition model used in Citation1 (called the competition LPA model) and significant weight to the explanation given for the observed case of coexistence. The explanation entails, however, some unusual aspects with regard to classic competition theory, including non-equilibrium dynamics, coexistence under increased intensity of inter-specific competition, and the occurrence of multiple mixed-type attractors. By multiple mixed-type attractors we mean a scenario that includes at least one coexistence attractor and at least one exclusion attractor. A coexistence attractor is one in which both species are present. An exclusion attractor is one in which at least one species is absent and at least one species is present. Park observed the coexistence case in an experimental treatment that also included cases of competitive exclusion, that is to say, he observed a case of what we have termed to be multiple mixed-type attractors.
Competition theory is primarily an equilibrium theory that is exemplified, for example, by the classic Lotka–Volterra model and its limited number of asymptotic outcomes: a globally attracting coexistence equilibrium; a globally attracting exclusion equilibrium; or two attracting exclusion equilibria. (In this context, globally attracting means within the positive cone of state space.) These three equilibration alternatives are illustrated by the Leslie–Gower model Citation7 (the discrete analog of the famous Lotka–Volterra differential equation model)
The competition LPA model, although applied specifically to species of Tribolium in Citation7, is none the less a rather general model that, unlike the Leslie–Gower model (or a Lotka–Volterra type model in general), accounts for life-cycle stages in the competing species. Therefore, the LPA model serves to illustrate that in general (when more biological details are included) competition theory is likely to be considerably more complicated and varied than that represented by classic Lotka–Volterra types of models. The competition LPA model is, like the Leslie–Gower model Equation(1), a discrete-time (difference equation) model. It differs from the Leslie–Gower model, however, in two basic ways: the state variables of the LPA model account for three life-cycle stages for each species (which mathematically introduces time delays and makes the model higher dimensional) and it utilizes ‘stronger’ (overcompensatory) nonlinearities. A natural question to ask is which of these two mechanisms most accounts for non-Lotka–Volterra dynamic scenarios and, in particular, for the occurrence of multiple mixed-type attractors? With regard to the first mechanism, it is shown in Citation13 that a result of introducing only a single life-cycle stage (specifically, a juvenile stage) in just one species in a Leslie–Gower model Equation(1)
can indeed result in multiple mixed-type attractors—specifically, the occurrence of exclusion equilibria in the presence of coexistence 2-cycles (provided inter-specific competition is sufficiently strong). A more robust occurrence of multiple attractors (equilibrium and cycles) of mixed type occurs if both species are given a juvenile stage Citation14.
Our goal here is investigate the second mechanism, namely the role of the nonlinearity in the occurrence of multiple mixed-type attractors. We do this by introducing a Ricker-type nonlinearity into the Leslie–Gower model Equation(1):
2. Equilibria
We can assume without loss in generality (by scaling the units of x and y) that c
ii
=1 in the Ricker competition model Equation(2). Therefore, we will consider, after relabeling c
12 as c
1 and c
21 as c
2, the competition model
Lemma 2.1
Assume (s
1, s
2)∈S. Let (x
t
, y
t
) denote the solution of the Ricker competition model
Equation(3)
with an initial condition (x
0, y
0) lying in the closure
of
. If n
1<1 then
. If n
2<1 then
.
Proof
If n
1<1 then all solutions of the linear equation satisfy
From the inequality
and u
0=x
0, an induction shows
for all t=0, 1, 2, …. A similar argument proves the assertion when n
2<1. ▪
We assume throughout the rest of the paper that both inherent net reproductive numbers satisfy n
i
>1. In this case, all solutions of Equation(3) are bounded and at least one species does not go extinct, as the following dissipativity and persistence theorem shows. The proof appears in the Appendix.
Theorem 2.1
Assume (s
1, s
2)∈S and both n
i
>1 in
Equation(3). There exist positive constants
such that all solutions with
satisfy
Theorem 2.2
Assume (s
1, s
2)∈S, that one of the inequalities
Equation(7)
holds, and that E
3
is a coexistence equilibrium. Then E
3
is unstable.
Proof
If one of the inequalities Equation(7) holds and if E
3 is a coexistence equilibrium, then the formula Equation(4)
for E
3 implies 1−c
1
c
2<0. A calculation shows
It follows from Theorem 2.2 that if at least one exclusion equilibrium is (hyperbolic and) stable, then either E
3 is not a coexistence equilibrium or, if it is, it is unstable. Consequently, with regard to equilibria, a mixed-type multiple attractor scenario is impossible for the competition model Equation(3). Thus, the Ricker competition model Equation(3)
and the classic Lotka–Volterra competition model have in common the impossibility of multiple mixed-type equilibrium attractors. In the next section we show, on the other hand, that it is possible for the Ricker model Equation(3)
to have multiple mixed-type non-equilibrium attractors.
3. Multiple mixed-type attractors
We want to investigate the possible occurrence of mixed-type non-equilibrium attractors in the Ricker model Equation(3) under symmetrically high inter-specific competition (as has been observed in more complicated models that include juvenile life-cycle stages Citation1, Citation8, Citation11, Citation13, Citation14). To carry out this investigation by means of a single parameter problem, we introduce the notation
,
and re-write the competition model Equation(3)
as
In the absence of species x t the dynamics of species y t are governed by the Ricker model equation
The 2-cycle Equation(10) yields an exclusion 2-cycle
By the linearization principle, the exclusion 2-cycle Equation(13) is (locally asymptotically) stable if the spectral radius of the matrix
is less than one. A calculation shows this matrix is triangular and its eigenvalues are
Theorem 3.1
Assume (s
1, s
2)∈S and that
is such that
Equation(9)
has a stable 2-cycle. Let c* denote the unique positive root of the equation
The loss of stability of the exclusion 2-cycle Equation(13) described in Theorem 3.1 suggests the occurrence of a bifurcation of planar 2-cycles from the exclusion 2-cycle Equation(13)
. 2-Cycles of the map defined by Equation(8)
correspond to fixed points of the composite map. The point
, corresponding to the exclusion 2-cycle Equation(13)
, is a fixed point of the composite for all values of c. On the other hand, a positive fixed point
of the composite corresponds to a coexistence 2-cycle. Positive fixed points of the composite satisfy the equations (obtained from the composite equations after x and y are cancelled)
Theorem 3.2
Assume (s
1, s
2)∈S and b
1∈I. If
, then a branch of coexistence 2-cycles bifurcates from the exclusion 2-cycle
Equation(13)
at c=c*.
By Theorem 3.1, the exclusion 2-cycle Equation(13) loses stability as c decreases through c*. By the exchange of stability principle (Citation16, p. 26) the bifurcating coexistence 2-cycles guaranteed by Theorem 3.2 are (locally asymptotically) stable if they exist for
(and unstable if they exist for
). Accordingly, our next goal is to determine the conditions under which the bifurcating coexistence 2-cycles occur for
. That is to say, we want to determine when
for the solution branch
of equations Equation(15)
. We can utilize the Lyapunov–Schmidt expansions used in the Appendix to establish Theorem 3.2 to calculate an expansion of c
′(0) for b
2 near
, the lowest order terms of which determine the sign of c
′(0). Details appear in the Appendix. To describe the results of this analysis, we need some further notation.
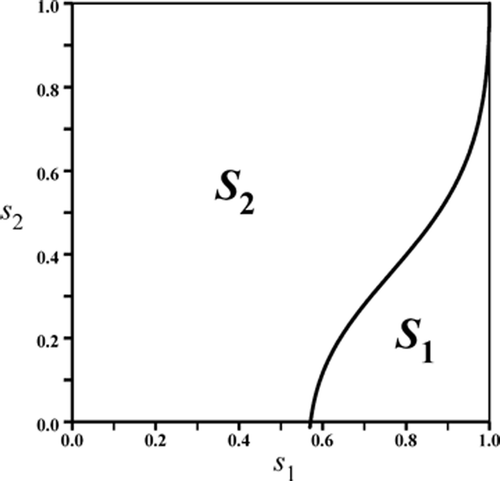
We partition the unit square into the union ,
where S
1 is the set of points (s
1, s
2)∈S that satisfy either
Figure 1. The unit square S for the survivorship parameters s
1 and s
2 in the competition model Equation(8) is partitioned into to sub-regions S
1 and S
2 corresponding to the two case in Theorems 3.3.
Theorem 3.3
Assume b
1∈I. For
, and
the bifurcating coexistence 2-cycles of the competition model
Equation(8)
(guaranteed by
Theorem 3.2
) are stable in either of the following cases.
-
(1)
and either
-
(a)
-
(b)
-
(c)
-
-
(2)
The subinterval in Theorem 3.4 is centered on the value
According to Equation(7), the exclusion equilibrium E
1 is stable if
In order for both the coexistence 2-cycles and the exclusion equilibrium to be stable in the cases (1b,c) and Equation(2) of Theorem 3.3, it is required that
. Necessary for this requirement is
This inequality is characterized in Lemma A.6 of the Appendix. These results, together with Theorem 3.3, lead to the following theorem.
Theorem 3.4
Assume b
1∈I. For
, and
the exclusion equilibrium E
1
and the bifurcating coexistence 2-cycles of the competition model
Equation(8)
are both stable if
and one of the following cases holds:
-
(1)
-
(2)
-
(3)
This theorem provides conditions on the parameters in the competition model Equation(8) under which there are multiple mixed-type attractors (specifically, a 2-cycle and an equilibrium). It follows from Lemma A.6 of the Appendix that in the cases not covered in Theorem 3.4 (namely when
or when
and b
1 is near the endpoints 1−s
1 and
of the interval I) either the 2-cycle is unstable or the equilibrium E
1 is unstable.
4. Discussion
The Ricker competition model Equation(8) can possess multiple mixed-type attractors. Theorem 3.4 provides some conditions under which there exist both a stable exclusion equilibrium and a stable coexistence 2-cycle. That theorem deals with values of b
2 greater than (but near) the critical period doubling bifurcation value
, values of b
1 less than the critical value
, and with the inter-specific competition coefficient c near a specified critical value c*. The theorem also requires that the survivorships (s
1, s
2) lie in the region S
1 of . This latter assumption means that the survivorship s
1 of species x is larger than the survivorship s
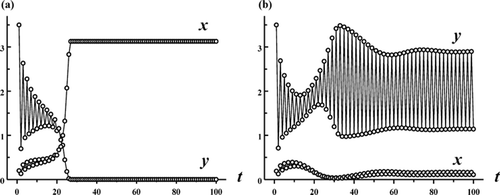
2 of species y. Therefore, Theorem 3.4 requires that there be an asymmetry between the two species in the sense that one species has a high reproductive rate and low survivorship in contrast to the other species, which has a low reproductive rate and a high survivorship. illustrates the existence of multiple mixed-type attractors under these conditions.
Figure 2. Each plot shows a solution of the Ricker competition model Equation(8) with b
1=8, b
2=10, s
1=0.65, s
2=0, r=1.1 and c=1.9. In plot (a) the initial conditions (x
0, y
0)=(0.2, 3.5) lead to competitive exclusion. In (b) the initial conditions (x
0, y
0)=(0.19, 3.5) lead to a competitive coexistence 2-cycle. See .
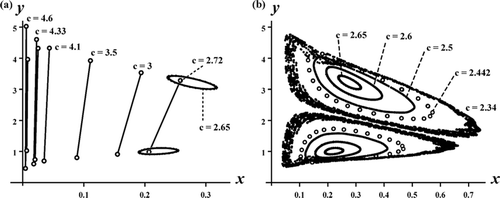
Theorem 3.4 implies the local bifurcation of stable coexistence 2-cycle only for c sufficiently large, namely, near the critical point c*. An interesting question concerns the global extent of this bifurcating branch of 2-cycles. What is the ‘spectrum’ of c values for which these coexistence 2-cycles occur? Numerous numerical explorations have shown that the bifurcation sequence displayed in is typical. As c decreases, and the coexistence 2-cycles bifurcate from the exclusion 2-cycle on the y-axis at c=c*, there exists a second critical value of c at which the coexistence 2-cycles lose stability because of an invariant loop (Sacker/Neimark or discrete Hopf ) bifurcation. The resulting coexistence (double) invariant loops persist until c reaches a third critical value at which the loops disappear in a global heteroclinic bifurcation. See and .
Figure 3. A sequence of phase plane plots shows the bifurcation of stable coexistence 2-cycles from the exclusion 2-cycles on the y-axis in the Ricker competition model Equation(8) as the competition coefficient c decreases through the critical value c*≈2.35. Model parameters are b
1=8, b
2=10, s
1=0.65, s
2=0, and r=1.1. Plot (a) shows a sequence of stable 2-cycles (open circles with connecting lines) that eventually destabilize and give rise to stable, double invariant loops as shown in plot (b). In plot (c) the double invariant loops eventually collide, under further decreases in c, and undergo a global, heteroclinic bifurcation involving the (saddle) coexistence equilibrium, the exclusion (saddle) equilibrium, the exclusion (saddle) 2-cycle located and their stable and unstable manifolds. For the parameter values in these plots, the exclusion equilibrium E
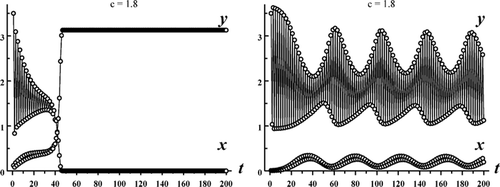
1:(x, y)≈(22.86, 0) is also stable and hence these plots contain multiple mixed-type attractors.
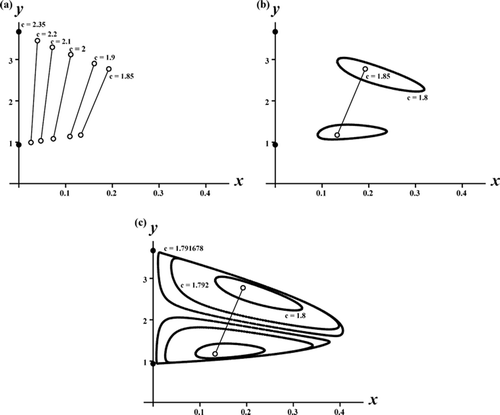
Figure 4. Each graph shows a solution of the Ricker competition model Equation(8) with b
1=8, b
2=10, s
1=0.65, s
2=0, r=1.1 and c=1.8. In plot (a) the initial conditions (x
0, y
0)=(0.12, 3.5) lead to competitive exclusion. In plot (b) the initial conditions (x
0, y
0)=(0.01, 3.5) lead to a competitive coexistence quasi-periodic oscillation (see , c)).
In this paper we have shown that the Ricker competition model Equation(8) cannot display a multiple mixed-type attractor scenario with only equilibria. On the other hand, Theorem 3.4 shows that multiple mixed-type attractor scenarios are possible with non-equilibrium attractors, specifically, with stable competitive exclusion equilibria and stable coexistence 2-cycles. Multiple mixed-type attractors scenarios are also possible for model Equation(8)
that involve other combinations of higher period cycles, quasi-periodic (as in ) and even chaotic attractors. shows one example. The analysis of such multiple attractor cases remains an open problem.
Figure 5. A sequence of phase plane plots shows the bifurcation of stable coexistence 4-cycles from the exclusion 4-cycles on the y-axis in the Ricker competition model Equation(8) as c decreases from the critical value c*≈4.77. Model parameters are b
1=8, b
2=14, s
1=0.8, s
2=0, r=0.8 and c=1.9. Plot (a) shows a sequence of 4-cycles the undergoes a period-halving bifurcation to 2-cycles which ultimately destabilize and give rise to stable, double invariant loops. As c decreases further, plot (b) shows the double invariant loops, which occasionally period lock, eventually giving rise to chaotic attractors. The chaotic attractors suddenly disappears when an ‘interior crisis’ occurs at a critical value of c. For the parameter values in these plots, the exclusion equilibrium E
1:(x, y)≈(3.69, 0) is also stable and hence these plots contain multiple mixed-type attractors.
Acknowledgements
We thank R.F. Constantino, J. Edmunds, S.L. Robertson and S. Arpin for helpful discussions. J.M. Cushing and S.M. Henson were supported in part by NSF grant DMS-0414142.
Notes
†Both eigenvalues of a 2 × 2 matrix A have absolute value less than 1 if and only if, −1< det A<1 and−(1+ det A)<trA<1+ det A. At least one eigenvalue has absolute value greater 1 if and only if one of the inequalities is reversed.
Related Research Data
References
- Edmunds , J. , Cushing , J. M. , Costantino , R. F. , Henson , S. M. , Dennis , B. and Desharnais , R. A. 2003 . Park's Tribolium competition experiments: a non-equilibrium species coexistence hypothesis . Journal of Animal Ecology , 72 : 703 – 712 .
- Park , T. 1957 . Experimental studies of interspecies competition. III, Relation of initial species proportion to the competitive outcome in populations of . Tribolium. Physiological Zoology , 30 : 22 – 40 .
- Park , T. , Leslie , P. H. and Mertz , D. B. 1964 . Genetic strains and competition in populations of Tribolium . Physiological Zoology , 37 : 97 – 162 .
- Leslie , P. H. , Park , T. and Mertz , D. M. 1968 . The effect of varying the initial numbers on the outcome of competition between two Tribolium species . Journal of Animal Ecology , 37 : 9 – 23 .
- Costantino , R. F. , Desharnais , R. A. , Cushing , J. M. , Dennis , B. , Henson , S. M. and King , A. A. 2005 . The flour beetle Tribolium as an effective tool of discovery . Advances in Ecological Research , 37 : 101 – 141 .
- Cushing , J. M. , Costantino , R. F. , Dennis , B. , Desharnais , R. A. and Henson , S. M. 2002 . Chaos in Ecology: Experimental Nonlinear Dynamics , New York : Academic Press .
- Leslie , P. H. and Gower , J. C. 1958 . The properties of a stochastic model for two competing species . Biometrika , 45 : 316 – 330 .
- Cushing , J. M. , LeVarge , S. , Chitnis , N. and Henson , S. M. 2004 . Some discrete competition models and the competitive exclusion principle . Journal of Difference Equations and Applications , 10 : 1139 – 1151 .
- Kulenović , M. and Merino , O. 2006 . Competitive-exclusion versus competitive-coexistence for systems in the plane . Discrete and Continuous Dynamical Systems Series B , 6 : 1141 – 1156 .
- Smith , H. L. 1998 . Planar competitive and cooperative difference equations . Journal of Difference Equations and Applications , 3 : 335 – 357 .
- Edmunds , J. 2001 . “ A study of a stage-structured model of two competing species ” . The University of Arizona . Ph.D. dissertation
- Edmunds , J. Multiple attractors in a discrete model of two competing species . Theoretical Population Biology , to appear
- Cushing , J. M. and LeVarge , S. Some discrete competition models and the principle of competitive exclusion . Proceedings of the Ninth International Conference (mcSingapore: . Difference Equations and Discrete Dynamical Systems , World Scientific .
- Cushing , J. M. , Henson , S. M. , Roeger and Lih-Ing . 2007 . A competition model for species with juvenile-adult life cycle stages . Journal of Biological Dynamics , 1 : 201 – 231 .
- Elaydi , S. N. 2005 . An Introduction to Difference Equations, , 3 , New York : Springer-Verlag .
- Kielhofer , H. 2004 . Bifurcation Theory: An Introduction with Applications to PDEs , Vol. 156 , 26 Berlin : Springer . Series in Applied Mathematical Sciences
Appendix
The proof of Theorem 2.1 utilizes the following lemma.
Lemma A.1
Consider the difference equation
with z
0, b, k>0, s≥0 and b+s>1. There exist positive constants
(independent of z
0) such that the solution satisfies
Proof
The maximum of bzexp(−kz) for z≥0 is bk
−1
e
−1. The inequalities and an induction show
Proof of Theorem 2.1
If x
0=0 or y
0=0 the result follows from Lemma A.1. Suppose . Note that the maximum of the function bxexp(−x) for x≥0 is be
−1. The inequalities
and
, together with a straightforward induction, show that
and
where u
t
and v
t
satisfy the linear difference equations
and
with initial conditions u
0=x
0 and v
0=y
0. Because s
i
<1, we have that
and
. As a result
Proof of Theorem 3.2
Define and w=c−c* and re-write the composite, fixed point equations Equation(15)
as
Lemma A.2
Assume (s
1, s
2)∈S and
. The bifurcating stable 2-cycles
Equation(10)
of the Ricker equation
, y
0>0 have, for
, the representations
Proof
The point on a 2-cycle Equation(10)
of the Ricker equation is a fixed point of the composite map. The fixed point equation reduces, after the cancellation of y, to
Lemma A.3
Assume (s
1, s
2)∈S and
. The critical value c* (at which the exclusion 2-cycle
Equation(13)
loses stability) has, for
, the representation
Proof
The critical value c* is the unique root of the equation Equation(14). By means of the expansions from Lemma A.2 and this equation, we seek the coefficients in the expansion
of the root. Substitution of these ϵ -expansions into the left-hand side of Equation(14)
and expanding in ϵ results, to first order, in
Using the ϵ expansions provided by Lemmas A.2 and A.3, we can obtain ϵ expansions, and hence lower order approximations, of δ for for
. To do this, we need to calculate (with the aid of a symbolic computer program) the partial derivatives p
z
|(0, 0, 0), p
x
|(0, 0, 0), q
z
|(0, 0, 0), q
x
|(0, 0, 0) and p
w
|(0, 0, 0). These have complicated formulas, only one of which we display here:
Proof of Theorem 3.3
Our goal is to determine when . From the equations
we obtain by implicit differentiation that
where
Lemma A.4
Assume (s
1, s
2)∈S and
. Let c* denote the unique positive root of the equation
Equation(14)
. If Ω>0 then the bifurcating branch of coexistence 2-cycles occurs for
and the 2-cycles are (locally asymptotically) stable. If Ω<0 then the bifurcating branch of coexistence 2-cycles in Ω occurs for
and the 2-cycles are unstable.
The proof of Theorem 3.3 will be complete when we determine the parameter values s
1, s
2, b
1, and r for which Ω>0. Since Ω0>0 for (s
1, s
2)∈S, the sign of Ω in Equation(A9) depends on that of Ω1, which in turn is the sign of the factor m(ln n
1). The term m(ζ) is a quadratic polynomial in ξ. Restricting our attention to b
1∈I, we need only investigate the sign of m(ξ) for ξ on the interval
which we denote by I*. Note that
and at the endpoints of I* we find that
and
The maximum of the quadratic m(ξ) occurs at
Lemma A.5
Assume (s
1, s
2)∈S and b
1∈I. Also assume
, and
. Then Ω>0 in either of the following cases.
-
(1)
-
(a)
-
(b)
-
(c)
-
-
(2)
In order to satisfy r<r* (to obtain the stability of the bifurcating coexistence 2-cycles in cases (1b,c) and Equation(2) of Theorem 3.3) and also
(to obtain the stability of E
1), it is necessary that
. This inequality is equivalent to
where
. (Recall m(ln n
1)<0 in cases (1b) and Equation(2)
of Theorem 3.3). This inequality does not hold at the endpoints of the interval I*. This is because
and
. In case Equation(2)
the maximum of m(ξ) occurs at the right endpoint
and as a result the inequality
does not hold. In cases (1b,c) then inequality holds at and near the roots ξ± of m(ξ).
Lemma A.6
In case
Equation(2)
of Lemma A.5,
. In cases (1b, c) of Lemma A.5,
if and only if
or
.