Abstract
In this paper we consider a non-autonomous multi-strain SIS epidemic model with periodic coefficients. Reproduction numbers and invasion reproduction numbers are derived which agree well with their counterparts usually derived from autonomous epidemic models. With conditions on these reproduction numbers typical results are obtained, such as the local and global stability of the disease-free equilibrium. Existence and uniqueness of a single-strain periodic solution is established. Based on conditions on the invasion reproduction numbers, local stability of the single-strain periodic solution is shown. In a two-strain version of the model, conditions for uniform strong persistence are derived, and coexistence of the two strains is established. Coexistence, however, does not occur if the transmission rates of the different strains are linearly dependent.
1. Introduction
The incidence of many infectious diseases fluctuates over time and often exhibits periodic behaviour. One of the well-known examples are data on weekly measles notification in England and Wales for the period 1948–1968 (see [Citation5, Figure 6.3]). The number of measles cases per week oscillates with a period between two peaks of 2 years. Other childhood diseases such as mumps, chicken-pox, rubella, and pertussis have also been found to exhibit seasonal behaviour. The periodicity of 1 year in many childhood diseases is attributed to the periodic contact rate the children have during academic school years. Although not restricted to children, influenza also exhibits distinct seasonality, with a season in the US spanning the time between October and May. During the first 2-3 years after the first case, the number of human cases of avian influenza also seemed to follow seasonal pattern Citation33.
Numerous articles have been devoted to explaining the periodicity in the disease incidence rates (see Citation18 for a review). Various scenarios how periodicity may arise in epidemic models have been set forward. Those include
| 1. | the possibility that the presence of certain classes can destabilize the equilibrium of models with constant coefficients and lead to sustain oscillations, | ||||
| 2. | the possibility that the incidence is not bilinear but has more complex form, | ||||
| 3. | the possibility that the transmission rates are periodic, | ||||
| 4. | the presence of delay(s) in the model, | ||||
| 5. | the presence of host age-structure, | ||||
| 6. | the interaction of two strains, one of which mutates into the other Citation20. | ||||
One of the key difficulties in working with non-autonomous models is defining the reproduction number ℛ0. Many of the articles in the literature do not define a reproductive number but work with threshold conditions that fit best the analysis. Ma and Ma Citation26 consider the problem of defining adequate reproduction numbers for various non-autonomous SIR and SEIR models. The authors argue that the reproduction number should be defined as the reproduction number of a constant coefficient system obtained from the non-autonomous system by averaging the time-dependent coefficients. In all cases they are able to establish local stability of the disease-free equilibrium with the usual condition on the reproduction number . However, Ma and Ma show through simulation that the SEIR model can have a solution where the number of infected individuals goes to zero despite that the reproduction number is larger than one (see [26, ]). Thus, the condition
does not necessarily lead to persistence.
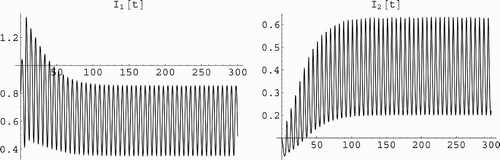
Figure 1. This figure gives an example of coexistence of the two strains in the population. The left figure illustrates that the number infected individuals with strain one I 1(t) persists and tends to an oscillatory solution. The right figure illustrates that the number infected individuals with strain two I 2(t) persists and tends to an oscillatory solution. The parameter values for both figures are: β1(t)=1.1(1+0.9 sin(t)), β2 (t)=1.5 (1+0.1 cos(t)), γ1=0.1, γ2=0.5, μ=0.5. The reproduction numbers of the two strains are ℛ1=1.83333, ℛ2=1.5.
The causative agents of many diseases are represented by multiple genetically distinct variants. Early autonomous multi-strain models suggested that competitive exclusion is the only possible outcome of the competition of many strains. Bremermann and Thieme Citation7 show that even if all reproduction numbers are larger than one, only the strain with the largest reproduction number persists while the remaining strains are eliminated. These results raised a question regarding the mechanisms that are responsible for the genetic diversity of pathogens in nature. Mechanisms such as super-infection Citation29 Citation32, coinfection Citation30, mutation Citation20, vertical transmission Citation22, and host age heterogeneity Citation28 were proposed as possible mechanisms that support diversity of pathogens. There is a significant literature devoted to the problem of competition of strains in autonomous multi-strain epidemic models which is reviewed in Citation21 Citation27 Citation42.
Diseases that exhibit seasonality may also be represented by pathogens that experience mutation and exist as multiple strains. The most notable example of this scenario is influenza. Several intrinsic mechanisms such as quarantine Citation34 or host age structure Citation8, leading to periodicity in autonomous epidemic models, have been suggested as possible mechanisms for the influenza seasonality. Other possible mechanisms such as crowding during cooler months (increased contact rate), weakening of the host immune system (increased susceptibility), and ability of the viral particles to persist in cooler temperatures Citation25 could be modelled with periodic transmission rates. This would require the development and analysis of multi-strain non-autonomous models. Multi-strain non-autonomous epidemic models do not seem to have been discussed so far.
In this paper we consider a multi-strain SIS model with periodic coefficients. Since we assume the total population size constant and equal to one, we eliminate the variable for the susceptible individuals. The resulting system is a variant of the classical Lotka–Volterra n-species competition system. In contrast to multi-strain non-autonomous epidemic models, Lotka–Volterra competition systems have been studied extensively not only in the constant coefficient case but also in the case of time dependent (periodic) coefficients. The pioneering work on non-autonomous Lotka–Volterra competition systems is due to Jim Cushing, whose contribution to science in general, and to mathematical biology in particular, we are celebrating today on occasion of his 65th birthday. In the late 1970s and early 1980s, Cushing authored a number of articles devoted to the non-autonomous versions of many classical ecological models: Lotka–Volterra predator–prey models Citation10 Citation13, Lotka–Volterra 2-species competition models Citation11 Citation14, n-species predator-prey/competition models Citation9, and the more-general Kolmogorov systems with periodic coefficients Citation12. In Citation11 Cushing showed that competing species may coexist in a competitive systems with periodic coefficients even if they will competitively exclude each other in a system with constant coefficients equal to the time averages of the periodic coefficients. Cushing's early results were extended in Citation31. Cushing's work motivated a number of articles on competitive non-autonomous Lotka–Volterra systems (e.g. Citation1 Citation3 Citation6 Citation16 Citation19 Citation23 Citation37). Even today the questions on global persistence and extinction of the species in such systems is still being investigated Citation2 Citation4 Citation24.
This paper is structured as follows. Section 2 introduces the model, its re-formulations, the reproduction numbers and results on extinction of the disease. Section 3 establishes the existence of a single-strain periodic solution corresponding to each strain. Section 4 derives conditions for the local stability of the single-strain periodic solutions. Section 5 considers a two-strain version of the model and establishes the existence and convergence to a periodic coexistence solution. Section 6 discusses the special case when the transmission rate is a product of a periodic contact rate and a constant probability of transmission, and establishes competitive exclusion in that case. Section 7 summarizes the results and draws conclusions.
2. A non-autonomous SIS model
In this section we introduce a non-autonomous SIS epidemic model with multiple strains. We study a model with recruitment and deaths. We assume all newly recruited individuals are susceptible. The recruitment and mortality rate are assumed equal and denoted by μ(t). The function μ(t) is assumed non-negative and periodic. In nature the recruitment rate needs to be periodic if it models seasonality in births. Many species breed in strictly defined breeding seasons and the birth/recruitment rates may be periodic. The transmission rate of strain i is denoted by β i (t) for i=1, …, n. All transmission rates are also assumed non-negative and periodic. The transmission rate may be periodic because of migratory patterns of host species. In human epidemic models the contact rate can vary with the time in the year as is the case with students academic years. Finally, we denote by γ i (t) the recovery rate from strain i. All recovery rates are also assumed non-negative and periodic. All periodic functions have period ω. The SIS model with multiple strains takes the form:
Proposition 2.1
The disease-free equilibrium
of the system
Equation(1)
is locally asymptotically stable if
for all j=1, …, n. If there exists
then the disease-free equilibrium ℰ0
is unstable.
In fact a stronger result is possible. The following proposition gives a sufficient condition for the eradication of strain j. We will see in the next section that if that condition fails to hold, then the corresponding strain can survive in the absence of competition.
Proposition 2.2
If
for some j, then
To see this result, notice that since S(t)≤1 we have
Proposition 2.2 implies that if for all j=1, …, n, then the disease-free equilibrium ℰ0 is globally asymptotically stable.
3. Existence of single-strain periodic solution
In this section we show that if for some j and
for all k=1, …, n and k≠j, then strain j may survive, while the rest will be eliminated. Without loss of generality we may assume
, and the remaining basic reproduction numbers are smaller than one. As a first step we assume I
k
(0)=0 for k=2, …, n. This implies that I
k
(t)=0 for all t. We will show that if
, then the equation
We are considering EquationEquation (4) on the domain
To show uniqueness, assume there exist two fixed points I p1 and I p2 which are distinct. Without loss of generality we may assume I p1<I p2. We have
Summarizing the results in this section we have the following theorem.
Theorem 3.1
For each i for which
there exist a unique, positive periodic function ξ
i
(t) such that
is a solution to the system
Equation(2)
. Furthermore, if
is a solution of the system
Equation(2)
starting from I
i
(0)>0, we have
To complete the proof of the Theorem 3.1 we have to establish the convergence to the periodic solution. We again assume that , and we consider the solutions of EquationEquation (4)
. Let I(t) be an arbitrary solution, starting from the initial condition I
0. We recall that I
p
is the initial condition for the periodic solution. We assume that I
0≠I
p
. We have two choices
or
. Assume
. The other case can be addressed in a similar way. Since the Poicarè map is increasing, we have
. Consequently, the sequence
is decreasing, and since it is bounded, it must have a limit:
4. Local stability of the single-strain periodic solutions
The single-strain periodic solutions play the role of the dominance equilibria in models with constant coefficients. Just as in the constant coefficient case, the stability properties of these single-strain periodic solutions determine the outcome of the competition of the strains. Determining the global stability of the single-strain periodic solutions is only possible in some cases but it is difficult in the general case. We, however, can consider the local stability. To determine the local stability of each single-strain periodic solution, one linearizes the system Equation(2)
around the single-strain periodic solution. Without loss of generality we may consider the single-strain solution corresponding to the periodic solution for strain one
. The other cases are symmetrical. Let
and
, where x
j
(t) with j=1, …, n are the perturbations. Then the linearized system takes the form:
From the Floquet exponents we can define the invasion reproduction numbers of strain j at the equilibrium of strain one:
The invasion reproduction numbers and the Floquet exponents are related: if and only if
. The results of this section can be summarized in the following theorem.
Theorem 4.1
The single-strain periodic solution
is locally asymptotically stable if
In the remainder of this section we establish global stability of the single-strain periodic solution under given conditions. These conditions are sufficient but not necessary. We again derive the result for the single-strain periodic solution of strain one. Analogous results hold for the other single-strain periodic solutions.
Theorem 4.2
If
and
for j=2, …, n, then the single-strain periodic solution
is globally stable.
To see this result we use Proposition 2.2. Let ε>0 be small so that . Since Proposition 2.2 implies that I
j
(t)→0 for j=2, …, n as t→∞. Then there exists T such that
5. Coexistence
In this section we show that coexistence in an oscillatory regime is possible. Although we believe that the result below can be extended to n strains, we consider a two-strain version of the model Equation(2).
Theorem 5.1
Let the sequence
be bounded. Then the sequence is converging, that is there exists
I
p
such that
This theorem says that if is an arbitrary solution of system Equation(15)
starting from an arbitrary initial condition
, then there exists a periodic solution of the system Equation(15)
, denoted by
such that the solution
converges to that periodic solution. The theorem does not say whether
and
. Thus, in theory the limit periodic solution could be one of the single-strain periodic solutions
or
whose existence was established in the previous sections.
In what follows we show that under certain conditions the solution of the system Equation(15) is persistent. We say that strain i is weakly uniformly persistent, if there exists δ>0, independent of the initial conditions, such that
Theorem 5.2
Assume
and that the invasion reproduction number of strain i is larger than one,
. Then, strain i is uniformly strongly persistent.
To see this result assume without loss of generality and
. The first condition guarantees that strain one in the absence of strain two has a non-trivial periodic solution. The second condition says that strain two can invade the periodic solution of strain one. A consequence of the second condition is that
. We will establish uniform strong persistence of strain two. The other case is established in the same way. First we establish the uniform weak persistence. Assume the contrary, that is assume that for every ε we have
. Then for a fixed ε>0 there exists T such that for all t>T, we have
.
From the first equation in system Equation(15) we obtain the following inequality:
The above result leads to the following corollary
Corollary 5.3
Assume
and
and that both invasion reproduction numbers are larger than one, that is
and
. Then, every solution converges to a coexistence periodic solution
where
and
.
To see this corollary, we notice that under the above conditions Theorem 5.2 implies that both I 1(t) and I 2(t) are uniformly strongly persistent, that is there exists δ>0 such that
We illustrate the periodic coexistence in . The results in Theorem 5.2 and Corollary 5.3 presumably can be obtained from Citation31 but the rescaling there makes the mathematical conditions imposed difficult to interpret biologically.
6. Is periodicity in the contact rate sufficient for coexistence?
We motivated the consideration of multi-strain epidemic models with periodic coefficients by the observed periodicity in childhood diseases and influenza. In the case of the childhood diseases it has been suggested that periodicity stems from the school-year system which makes the contact rate among children periodic. Is periodic contact rate sufficient to sustain pathogen diversity? We address this question in this section. We recall that in the constant coefficient case the transmission rate of strain i is a product of the contact rate of the population c (assuming that all classes have the same contact rate) and the probability of transmission of strain i – p i :
Surprisingly, coexistence does not occur, and competitive exclusion is the only outcome of the multi-strain competition in periodic environments, even if the remaining parameters μ(t) and γ i (t) are still assumed periodic (see Citation43). As before, the strain with the largest reproductive number eliminates the rest. This result is established in the following theorem, where we have assumed without loss of generality that strain one has the largest reproduction number.
Theorem 6.1
Assume
for i=1, …, n. Assume, in addition, that
for j=2, …, n. Then,
To see the claim in this theorem, for a fixed j>1 consider the quantity
Then, similarly to EquationEquation (4) one can show that strain one persists, and in fact the solutions of the system Equation(2)
converge to
.
7. Summary of results
In this paper we introduce an n-strain SIS model with periodic coefficients. The birth and death rates are assumed to be the same so that the total population is constant and equal to one. That is, the number of individuals in each class is in fact the proportion of individuals in the various classes. For this model we succeed in defining reproduction numbers for each strain ℛ
i
which agree with the corresponding reproduction numbers of the autonomous system obtained from system Equation(1) by assuming all coefficients constant and equal to the averages of time dependent coefficients. We establish the following results:
| 1. | There is a unique disease-free equilibrium which is locally and globally stable if all reproduction numbers are smaller than one. The disease-free equilibrium is unstable if one of the reproduction number is greater than one. These observations agree with similar observations made for multi-strain autonomous models. | ||||
| 2. | Moreover, we establish that if | ||||
| 3. | We establish that to each strain there corresponds a unique single-strain periodic solution, if its reproduction number is greater than one. This periodic solution is globally stable in the hyper-plane where all other strains are extinct. | ||||
| 4. | The single-strain periodic solution is locally stable if the invasion numbers of all other strains at the equilibrium of the resident strain are less than one. The single-strain periodic solution is unstable if there is an invasion number grater than one. | ||||
| 5. | For the model with two strains we establish the existence of at least one periodic coexistence solution. As a part of this result, we derive conditions on the reproduction numbers and invasion reproduction numbers that guarantee uniform strong persistence of a strain. | ||||
| 6. | Finally, for the n-strain model we establish that if the transmission rate is periodic because the contact rate is periodic, but the probability of transmission given a contact is constant, | ||||
Acknowledgements
This article was inspired by some modelling of avian influenza that my student Joseph Lucchetti did. The author thanks Horst R. Thieme for his valuable comments. The author also thanks the referees for their constructive remarks. The author acknowledges partial support from the NSF under grant DMS-0817789.
Related Research Data
References
- Ahmad , S. 1993 . On the nonautonomous Volterra–Lotka competition equations . Proc. Amer. Math. Soc. , 117 : 199 – 204 .
- Ahmad , S. and Lazer , A. C. 2005 . Average growth and extinction in a competitive Lotka–Volterra system . Nonlinear Anal. , 62 : 545 – 557 .
- Ahmad , S. and Montes de Oca , F. 1998 . Extinction in nonautonomous T-periodic competitive Lotka–Volterra system . Appl. Math. Comput. , 90 : 155 – 166 .
- Ahmad , S. and Stamova , I. 2008 . Survival and extinction in competitive systems . Nonlinear Anal. Real World Appl. , 9 ( 2 ) : 708 – 717 .
- Anderson , R. M. and May , R. M. 1991 . Infectious Diseases of Humans: Dynamics and Control , Oxford : Oxford University Press .
- Battaaz , A. and Zanolin , F. 1998 . Coexistence states for periodic competitive Kolmogorov systems . J. Math. Anal. Appl. , 219 : 179 – 199 .
- Bremermann , H. J. and Thieme , H. R. 1989 . A competitive exclusion principle for pathogen virulence . J. Math. Biol. , 27 : 179 – 190 .
- Castillo-Chavez , C. , Hethcote , H. W. , Andreasen , V. , Levin , S. A. and Liu , W. M. 1989 . Epidemiological models with age structure, proportionate mixing, and cross-immunity . J. Math. Biol. , 27 ( 3 ) : 233 – 258 .
- Cushing , J. M. 1976 . Stable limit cycles of time dependent multi-species interactions . Math. Biosci. , 31 : 259 – 273 .
- Cushing , J. M. 1977 . Periodic solutions of time dependent predator-prey systems . SIAM J. Appl. Math. , 32 ( 1 ) : 82 – 95 .
- Cushing , J. M. 1980 . Two species competition in a periodic environment . J. Math. Biol. , 10 : 385 – 400 .
- Cushing , J. M. 1982 . Periodic Kolmogorov systems . SIAM J. Math. Anal. , 13 ( 5 ) : 811 – 827 .
- Cushing , J. M. 1984 . Periodic two-predator, one-prey interactions and the time sharing of a resource niche . SIAM J. Appl. Math. , 44 : 392 – 410 .
- Cushing , J. M. 1986 . Periodic Lotka–Volterra competition equations . J. Math. Biol. , 24 : 381 – 403 .
- Dietz , K. 1976 . “ The incidence of infectious diseases under the influence of seasonal fluctuations ” . In Mathematical Models in Medicine , Edited by: Berger , Jürgen . 1 – 15 . New York : Springer-Verlag . Lecture Notes in Biomathematics Vol. 11
- Golpalsamy , K. 1985 . Global asymptotic stability in a periodic Lotka–Volterra system . J. Austral. Math. Soc. Ser. B , 27 : 66 – 72 .
- Hethcote , H. W. 1973 . Asymptotic behavior in a deterministic epidemic model . Bull. Math. Biol. , 36 : 607 – 614 .
- H.W. Hethcote and S.A. Levin, Periodicity in epidemiological models, in Applied Mathematical Ecology (Trieste, 1986), S.A. Levin, T.G. Hallam, and L.J. Gross, eds., Biomathematics Vol. 18, Springer, Berlin, 1989, pp. 193–211
- Korman , P. 1992 . Some new results on the periodic competition model . J. Math. Anal. Appl. , 171 : 131 – 138 .
- Li , J. , Zhou , Y. , Ma , Zh. and Hyman , J. M. 2004 . Epidemiological models for mutating pathogens . SIAM J. Appl. Math. , 65 : 1 – 23 .
- Lipsitch , M. and O'Hagan , J. J. 2007 . Patterns of antigenic diversity and the mechanisms that maintain them . J. R. Soc. Interface , 4 : 787 – 802 .
- Lipsitch , M. , Siller , S. and Nowak , M. A. 1996 . The evolution of virulence in pathogens with vertical and horizontal transmission . Evolution , 50 : 1729 – 1741 .
- Lisena , B. 2004 . Global stability in periodic competitive system . Nonlinear Anal. Real World Appl. , 5 : 613 – 627 .
- Lisena , B. 2006 . Competitive exclusion in a periodic Lotka–Volterra system . Appl. Math. Comput. , 177 : 761 – 768 .
- Lofgren , E. , Fefferman , N. H. , Naumov , Y. N. , Gorski , J. and Naumova , E. N. 2007 . Influenza seasonality: underlying causes and modeling theories . J. Virol. , 81 : 5429 – 5436 .
- Ma , J. and Ma , Z. 2006 . Epidemic threshold conditions for seasonally forced SEIR models . Math. Biosci. Eng. , 3 : 161 – 172 .
- Martcheva , M. , Bolker , B. M. and Holt , R. D. 2008 . Vaccine-induced pathogen strain replacement: what are the mechanisms? . J. R. Soc. Interface , 5 : 3 – 13 .
- Martcheva , M. , Pilyugin , S. S. and Holt , R. D. 2007 . Subthreshold and superthreshold coexistence of pathogen variants: the impact of host age-structure . Math. Biosci. , 207 : 58 – 77 .
- May , R. and Nowak , M. 1994 . Superinfection, metapopulation dynamics, and the evolution of diversity . J. Theor. Biol. , 170 : 95 – 114 .
- May , R. and Nowak , M. 1995 . Coinfection and the evolution of parasite virulence . Proc. Roy. Soc. London B , 261 : 209 – 215 .
- de Mottoni , P. and Schiaffino , A. 1981 . Competition systems with periodic coefficients: a geometric approach . J. Math. Biol. , 11 : 319 – 335 .
- Nowak , M. and May , R. 1994 . Superinfection and the evolution of parasite virulence . Proc. Royal Soc. London B , 255 : 81 – 89 .
- Normile , D. and Enserink , M. 2007 . With change in the seasons, bird flu returns . Science , 315 : 448
- Nuño , M. , Feng , Z. , Martcheva , M. and Castillo-Chavez , C. 2005 . Dynamics of two-strain influenza with isolation and partial cross-immunity . SIAM J. Appl. Math. , 65 : 964 – 982 .
- I.B. Schwartz, Nonlinear dynamics of seasonally driven epidemic models, in Biomedical Systems Modelling and Simulation (Paris, 1988), J. Eisenfeld and D.S. Levine, eds., IMACS Ann. Comput. Appl. Math. Vol. 5, Baltzer, Basel, 1989, pp. 201–204
- Smith , H. L. 1983 . Multiple stable subharmonics for a periodic epidemic model . J. Math. Biol. , 17 : 179 – 190 .
- Smith , H. L. 1986 . Periodic solutions of periodic competitive and cooperative systems . SIAM J. Math. Anal. , 17 : 1289 – 1318 .
- Smith , H. L. 1995 . Monotone Dynamical Systems: an Introduction to the Competitive and Cooperative Systems , Providence, RI : AMS .
- Smith , H. L. and Waltman , P. 1995 . The Theory of the Chemostat , Cambridge : Cambridge University Press .
- Thieme , H. R. 2000 . Uniform persistence and permanence for non-autonomous semiflows in population biology . Math. Biosci. , 166 : 173 – 201 .
- Thieme , H. R. 2003 . Mathematics in Population Biology , Princeton, NJ : Princeton University Press .
- Thieme , H. R. 2007 . “ Pathogen competition and coexistence and the evolution of virulence ” . In Mathematics for Life Science and Medicine , Edited by: Takeuchi , Y. , Iwasa , Y. and Sato , K. 123 – 153 . Berlin, Heidelberg : Springer-Verlag .
- Zhang , T. and Teng , Z. 2007 . On a nonautonomous SEIR model in epidemiology . Bull. Math. Biol. , 69 : 2537 – 2560 .