Abstract
We propose a discrete-time, age-structured population model to study the impact of Allee effects and harvesting. It is assumed that survival probabilities from one age class to the next are constants and fertility rate is a function of weighted total population size. Global extinction is certain if the maximal growth rate of the population is less than one. The model can have multiple attractors and the asymptotic dynamics of the population depend on its initial distribution if the maximal growth rate is larger than one. An Allee threshold depending on the components of the unstable interior equilibrium is derived when only the last age class can reproduce. The population becomes extinct if its initial population distribution is below the threshold. Harvesting on any particular age class can decrease the magnitude of the possible stable interior equilibrium and increase the magnitude of the unstable interior equilibrium simultaneously.
1. Introduction
Allee effects Citation1, the decrease in population growth rates at small populations or densities, include an inability to find mates, reduced foraging efficiency in social animals, lessened defences against predators, reduced reproductive success in cooperative breeders, and reduced fertilization success in broadcast spawners Citation4. As a consequence, Allee effects play a crucial role in resource management and conservation. Moreover, Allee effects have also been observed in the context of biological control, both to the introduction of the control agent and also to the extirpation of the pest requiring control Citation10. Recently, there has also been a surge of interest and a need to investigate Allee effects for epidemic models Citation9 Citation11 Citation20. See Citation5 Citation7 Citation8 Citation12 Citation13 Citation14 Citation15 Citation16 Citation17 Citation23 Citation24 and references cited therein for population models of Allee effects.
In this work, we propose a general age-structured population model with Allee effects and harvesting to study the impact of these mechanisms on the population level. In Citation13 Citation15, the author studied Allee effects for a two-age classes population model and models of host-parasitoid interaction in which the host population is classified into two age classes and only the adult host can reproduce. Moreover, the fertility rates in Citation13 Citation15 depend only on the adult population size and also satisfy some monotonic assumptions which include for example the modified Beverton–Holt functions. Using this monotonic assumption, a global analysis for the single population model is obtained. In the present study, this monotonic assumption is relaxed and it is also assumed that individuals in each age class can reproduce with fertility rate depends on a weighted total population size. Moreover, the population is constantly being harvested with harvesting rates depending on age classes.
Our results are expressed in terms of a threshold, the maximal growth rate of the population. The population will become extinct, independent of initial population distributions, if this growth rate is less than one. When the maximal growth is larger than one, the system may have two stable equilibria and the fate of the population then depends on its initial distribution. An Allee threshold in terms of the unstable interior equilibrium is deduced when only the last age class can reproduce. The population will become extinct if its initial distribution is below the threshold. Bifurcation analysis is also performed when there is only two age classes. While harvesting on any particular age class can drive the maximal growth rate of the population to below one on one hand, it can also decrease the magnitude of the possible stable interior equilibrium and increase the magnitude of the unstable interior equilibrium on the other hand. The overall effect of harvesting is negative. There is no lessened intraspecific competition observed in the model due to harvesting.
In the following section, a general n-age classes model is formulated and analysed. Section 3 derives an Allee threshold of the model when only the last age class can reproduce. Specific examples and simulations are presented in Section 4. The final section provides a summary and discussion.
2. An age-structured model with Allee effects and harvesting
In this section, we derive a general discrete-time, age-structured population model with Allee effects and harvesting. The survival probabilities are independent of time and population density, whereas the fertility function depends on weighted total population size. We present conditions for global extinction and for the existence of non-extinct equilibrium. We also study the stability of the equilibria and equilibrium size relative to model parameters.
2.1. The model, threshold, and equilibrium
Consider a population with n age classes, n≥2. Let x
i
(t) denote the population size of age class i at time t, t=0, 1, …. The survival probability from age class i to age class i+1 is denoted by s
i
, 0<s
i
<1, . The fertility rate f is assumed to be a function of weighted total population size
due to intra-specific competition, where p
i
≥0 for 1≤i≤n. Since each age class in general will have different fertility rate, the fertility function f is scaled for each age class by F
i
, F
i
≥0 for 1≤i≤n. In particular, if F
i
=0, then the ith age class does not reproduce. We assume that the last age class is reproductive since we can simply ignore the last few age classes which do not contribute to reproduction. Therefore, we assume that F
n
>0 and p
n
>0. Let h
j
denote the age-specific harvesting rate, 0≤h
j
<1, 1≤j≤n. The model takes the following form:
Let
(H1) , f(0)≥0, β f(0)<1 and f(x)>0 for x>0. There exists m>0 such that
for 0≤x<m and
for x>m, and
.
The fertility function f increases first due to positive effect of increasing population size, reaches a maximum at population level m, and then declines due to intra-specific competition among individuals. A modified Beverton-Holt function for the fertility rate is analysed in Citation13 and a more general fertility function that satisfying for x>0 is studied in Citation15. Both of these models consist of only two age classes, juvenile, and adult. The assumptions of fertility function f in the present study do not have these restrictions.
It is clear that solutions of EquationEquation (1) remain nonnegative and EquationEquation (1)
always has a trivial steady state
where the population is extinct. If a component of a nontrivial steady state is positive, then all of the components of the steady state are positive. Therefore, we consider the existence of an interior steady state
. Let
Note if is a solution of EquationEquation (5)
, then a direct computation shows that the components x¯
i
, i=1, …, n, of the interior steady state are given by
Theorem 2.1
If β f(m)<1, then
is globally asymptotically stable for system
Equation(1)
.
Proof
Let be a solution of EquationEquation (1)
, where T denotes matrix transpose. Since
for all x≥0, we see that
for t≥0, where A
m
is given by
When β f(m)=1, the maximal growth rate of the population is one. System Equation(1) then has a unique interior steady state
with
. Since
, the Jacobin matrix of EquationEquation (1)
evaluated at E*, J(E*), is A
m
given in EquationEquation (8)
. Using the argument as in the proof of Theorem 2.1, we can verify that the dominant eigenvalue of J(E*) is now 1. Therefore, E* is nonhyperbolic, and its local stability cannot be determined using the linear theory.
Theorem 2.2
If β f(m)=1, then Equation
Equation(1)
has two steady states
and
where E
0
is locally asymptotically stable and E* is nonhyperbolic.
Suppose now β f(m)>1. Then EquationEquation (5) has two positive solutions X
i
* with
, and EquationEquation (1)
has two interior steady states
On the other hand, the Jacobian matrix of system Equation(1) evaluated at E
2*, J(E
2*), is given by
In the following, we derive a sufficient condition for which E
2* is locally asymptotically stable when either EquationEquation (12) or Equation(13)
is not satisfied. Towards this end, using the characteristic equation given by EquationEquation (14)
and applying Rouche's theorem Citation3, we let
and
. On the unit circle,
Theorem 2.3
Let β f(m)>1. Then system
Equation(1)
has three steady states E
0
and E
i
*, i=1, 2, where E
0
is locally asymptotically stable and E
1* unstable. If either Equations
Equation(12)
and
Equation(13)
are satisfied or Equation
Equation(15)
holds, then E
2* is locally asymptotically stable.
Note that if F
i
=F>0 and p
i
=1 for 1≤i≤n, i.e., if EquationEquation (13) holds, then EquationEquation (15)
becomes
2.2. Equilibrium size and parameters
In this subsection, we study the effects of parameters relative to the magnitude of the equilibrium. Since , β is a decreasing function of h
i
and an increasing function of both F
i
and s
i
for all i. From EquationEquation (5)
we have
From the above discussion, we see that the weighted total population size X 2* in the equilibrium E 2* increases with increasing survival probability s i and fertility rate F i but X 2* decreases with increasing harvesting rate h i . Therefore, harvesting can reduce weighted total population size in the equilibrium E 2*, which may be locally asymptotically stable.
3. Allee thresholds
It is well known that populations are more likely to go extinct if the populations are subject to Allee effects Citation4. In this section, we will derive Allee thresholds for model Equation(1) when β f(m)>1, i.e. the maximal growth rate of the population is greater than one. Since the extinction steady state
is locally asymptotically stable, there exists a basin of attraction for E
0. We only consider the case when F
i
=0 for
. System Equation(1)
then becomes
Since EquationEquation (24) is a special case of system Equation(1)
, we see that global extinction occurs if β f(m)<1. When β f(m)=1, then an increase of harvesting rate h
i
will drive the maximal growth rate to below one so that global extinction will happen. Moreover, since many life history parameters cannot be estimated precisely, the case of β f(m)=1 is hardly attained in the real world situation. We are more interested in the case when β f(m)>1, where system Equation(24)
has two interior steady states
, i=1, 2. Denoting the two positive solutions of EquationEquation (5)
by X
i
* as in the previous section, then
Suppose that initial conditions of EquationEquation (24) satisfy
Theorem 3.1
If solutions of Equation
Equation(24)
with initial conditions satisfy Equation
Equation(25)
, then the solutions converge to
.
Proof
Let be a solution of EquationEquation (24)
with initial condition satisfying EquationEquation (25)
. We first claim that
Theorem 3.1 deduces the following forward invariant rectangular region
Recall from EquationEquation (18) we arrived at
for all j. Therefore, an increase of fertility F
n
can make the region Γ smaller and reduce the possibility for population extinction. On the other hand, from EquationEquation (19)
we see that an increase of h
j
will increase x
11* for any j=1, …, n. As a consequence, an elevated harvesting on any age classes will enlarge Γ in the x
1-direction. Since EquationEquation (20)
also holds, the overall effect of harvesting on the Allee threshold is complicated and is not easy to determine if the number of age classes of the population is large. However, the total weighted population level X
1* will always increase when harvesting is implemented on any age classes.
4. Examples and bifurcations
In this section, we use specific examples for the fertility functions to discuss possible bifurcations. We consider two age-classes. The model then takes the following form:
We now use the weights of total population size in the fertility function as a bifurcation parameter. Specifically, we let
On the other hand, condition Equation(36) is equivalent to
Proposition 4.1
System
Equation(34)
does not undergo a +1 bifurcation when E
2* loses its stability. If
then E
2* as an interior steady state of Equation
Equation(34)
does not loss its stability via a discrete Hopf bifurcation. In addition, if
then E
2* is locally asymptotically stable for Equation
Equation(34)
.
If inequalities Equation(39) and Equation(41)
are satisfied, then a simple calculation yields
Proposition 4.2
Let inequalities
Equation(39)
and
Equation(41)
hold. Then E
2* is always unstable for θ≥0 if
and E
2* is locally asymptotically stable for
if
.
In the following, we use specific fertility functions to investigate the system Equation(34). All the simulations below are performed using MatLab. Since Beverton–Holt type functions appear frequently in the fishery literature, we first consider a Beverton–Holt type function with Allee effects. Specifically, we let
We let λ=a=1 in our numerical simulations with the following parameter values:
Figure 1. This figure provides simulation results for system Equation(34) with f given by EquationEquation (44)
. (a) and (b) are bifurcation diagrams with h
1=0.1 and h
1=0.5, respectively. The rest of the plots are basins of attraction of (0, 0) for different parameter values. In particular, F
1=10 and p
1=0 in (c), F
1=0 and p
1=0 in (d), F
1=10 and p
1=1.5p
2 in (e), and F
1=0 and p
1=1.5p
2 in (f).
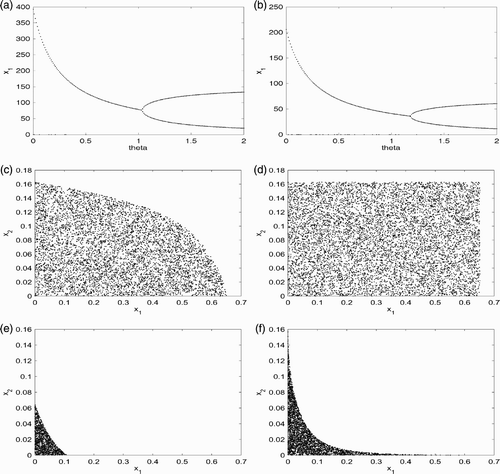
A period-doubling bifurcation occurs and the system has a period-two solution when θ is just beyond 1 which agrees with the earlier calculation of . We did not obtain more complicated dynamical behaviour by varying parameter values and increasing θ further. See for the same parameter values with h
1=0.1 been increased to h
1=0.5. In this case
. Recall from EquationEquation (22)
that an increase of h
1 will decrease x
2j
* for j=1, 2, and our simulations in and confirm this analytical result. Comparing and , we also see that an increase of h
1 with every other parameter value being fixed can increase the θ value for which the period-doubling bifurcation occurs. From this point of view, increasing harvesting rate of the young can help to stabilize the population. Similar results are obtained if we increase the number of age classes from 2 to 3.
It was shown in Theorem 3.1 in terms of the present content that if the initial population distribution lies inside the rectangular region , then the population goes extinct. In Theorem 3.1, it was assumed that only the last age class can reproduce. In this two-age classes model with F
1=0, we have
,
,
, and
and
when θ=0. We plot those initial conditions for which the solutions converge to (0, 0). In we used the same parameter values as in with p
1=0. provides the region with the addition that F
1=0. Note that in this situation of
, the basin of attraction of (0, 0) has been studied in Citation13
Citation16 which is given precisely by Γ. From these two plots, we see that the reproduction of smaller age class can have a smaller region of basin of attraction for the extinction steady state (0, 0). This observation was derived in EquationEquation (18)
along with Theorem 3.1.
We repeat the same simulations with p
1=1.5p
2 where the model has an asymptotically stable 2-cycle. and plot basin of attraction for (0, 0) when F
1=10 and F
1=0, respectively. When F
1=0, we see that and
and the region
lies inside the numerically simulated basin of attraction of (0, 0). Since the region in is smaller than the region in , we arrive at the same conclusion that reproduction of the young can help the population to avoid extinction. This conclusion was analytically deduced in EquationEquation (18)
and our numerical simulations confirm this earlier analytical finding.
We next consider a Ricker type function with Allee effects for the fertility rate. The Ricker functions have also been used frequently in the fishery modelling. Let
Recall from EquationEquation (43) that
if and only if
. When f is given by EquationEquation (45)
, this latter inequality is equivalent to
Let
. Then
if and only if h(X
2*)<0. Note that h is a concave down parabola with positive vertical intercept and h(K)≤0 is equivalent to r(K−c)≥4. Moreover, it can be verified that X
2*>K if β>1. Hence, we have
when β>1 and r(K−c)≥4.
Let
Figure 2. This figure provides simulation results for system Equation(34) with f given by EquationEquation (45)
and parameters are being chosen so that θ
H
=0.00561<θ
P
=0.00600. (a) is the time evolution of the x
1 component of a solution when θ=0.0059. (b) and (c) are the time evolutions of the x
1 component of a solution when θ=0.00600001, while θ=0.006006 in (d) and θ=0.00601 in (e). Plot (f) is the bifurcation diagram for θ in [0.0057, 0.0078].
![Figure 2. This figure provides simulation results for system Equation(34) with f given by EquationEquation (45) and parameters are being chosen so that θ H =0.00561<θ P =0.00600. (a) is the time evolution of the x 1 component of a solution when θ=0.0059. (b) and (c) are the time evolutions of the x 1 component of a solution when θ=0.00600001, while θ=0.006006 in (d) and θ=0.00601 in (e). Plot (f) is the bifurcation diagram for θ in [0.0057, 0.0078].](/cms/asset/2381f2c6-df02-45fd-8da0-2f494ec4c556/tjbd_a_439086_o_f0002g.gif)
We now decrease θ to 0.00562 and simulate solutions with randomly generated initial conditions that are very close to E
2*. As stated in Proposition 4.2, since E
2* is locally asymptotically stable, such solutions converge to E
2. Simulations confirm this result and a particular solution is plotted in . When we decrease θ further to 0.0056009, we obtain an invariant closed curve solution in the x
1
x
2- plane by truncating the first 30,000 iterations and recording the next 1000 iterations. An invariant closed curve is plotted in . Therefore, a discrete Hopf bifurcation does occur at . We plot bifurcation diagrams for the model using θ as the bifurcation parameter. The θ value is set to be 0.0059 initially and decreases by 0.00001 for each step until
for the diagram provided in . We see that the model is chaotic when θ decreases to below θ
H
from above. presents another bifurcation diagram with θ ranging from 0.0001 to 0.0059. The population goes extinct for all small θ values as can be seen from and also for large θ values as demonstrated in .
Figure 3. This figure provides simulation results for system Equation(34) with f given by EquationEquation (45)
. The parameter values are the same as those in except θ. (a) plots a solution when θ=0.00562. In (b), there is an invariant closed curve solution when θ=0.0056009. (c) and (d) are bifurcation diagrams of EquationEquation (34)
with θ∈[0.0049, 0.0059] and θ∈[0.0001, 0.0059], respectively.
![Figure 3. This figure provides simulation results for system Equation(34) with f given by EquationEquation (45). The parameter values are the same as those in Figure 2 except θ. (a) plots a solution when θ=0.00562. In (b), there is an invariant closed curve solution when θ=0.0056009. (c) and (d) are bifurcation diagrams of EquationEquation (34) with θ∈[0.0049, 0.0059] and θ∈[0.0001, 0.0059], respectively.](/cms/asset/90da0a30-d480-44ca-9adb-69c967e38eaa/tjbd_a_439086_o_f0003g.gif)
5. Discussion
The interest of studying Allee effects on population and epidemic models has been increased in recent years due to fragmentation of habitats, invasion of new species, and biological control of pest, etc Citation4. In this study, we propose an age-structured model of a single population with Allee effects and harvesting to investigate the impact of these mechanisms on the population level. The dynamics of the population depend on the lumped parameter, β f(m), which can be interpreted as the maximum growth rate of the population. It is the maximum average number of offspring that an individual can reproduce during its life span. The population will definitely go extinct if the maximal growth rate is less than one (cf. Theorem 2.1). The population may survive if the maximal growth rate is larger than one. In this situation, the model has two interior steady states E
1* and E
2*, where E
1* is always unstable. The other interior steady state may be locally asymptotically stable under some conditions (cf. EquationEquations (12) and Equation(13)
, or EquationEquation (15)
). Therefore, the model may exhibit multiple attractors and the fate of the population depends on its initial distribution.
Since the extinction steady state E
0 is always locally asymptotically stable, the basin of attraction provides a threshold for the population below which the population becomes extinct. If we assume that only the last age class can reproduce with fertility rate depends on the weighted total population size, then the region Γ can be systematically deduced (cf. Theorem 3.1) and is given by EquationEquation (33). The population will become extinct if its initial population distribution lies inside of Γ which depends on the components of E
1*. Since β is a decreasing function of the harvesting rate h
i
, an increase of h
i
will increase X
1* and decrease X
2*. As a consequence, the weighted threshold
in E
1* is larger, and the system has a smaller weighted population size
in the equilibrium E
2*. Therefore, the population may stabilize in a smaller equilibrium when E
2* is locally asymptotically stable and harvesting is in effect.
On the other hand, the weighted population size in the equilibrium X
2* increases with increasing fertility rate F
i
and survival probability s
i
by EquationEquation (17). Also EquationEquation (18)
implies that an increase of F
i
can reduce x
1j
* for all i and j. As a result, increasing fertility rate F
i
has an overall positive effect on the population. However, the impact of h
j
on x
1i
* is complicated and cannot be determined explicitly as demonstrated at the end of Section 2. Furthermore, an increase of harvesting rate h
i
may decrease the maximal growth rate β f(m) to below one so that global extinction can occur.
We also investigate a general two-age classes model, system Equation(34), and with two specific fertility functions, EquationEquations (44)
and Equation(45)
. Using θ as our bifurcation parameter, where
, we conclude that the only possible bifurcation is either a period-doubling or a discrete Hopf bifurcation when the interior steady state E
2* loses its stability for general system Equation(34)
(cf. Proposition 4.1). Moreover, E
2* is unstable for all θ≥0 if
and E
2* is locally asymptotically stable for
if
(cf. Proposition 4.2). However, a discrete Hopf bifurcation cannot occur if a modified Beverton–Holt function such as EquationEquation (44)
is used as the fertility function (cf. Proposition 4.1). We numerically simulate this particular model. We also simulate system Equation(34)
with f given by EquationEquation (45)
. From this numerical study, we see that an early reproduction (that is, F
1>0) can help the population to avoid extinction for both fertility functions. When a modified Ricker type function Equation(45)
is used as the fertility function with two age classes, we simulate the model with parameter values such that
, and thus E
2* is locally asymptotically stable when
. A period-doubling bifurcation occurs when
and we do not obtain more complicated dynamical behaviour as we increase θ. Moreover,the system does exhibit an invariant closed curve solution when θ just passes θ
H
from above. The system is chaotic for some θ values as we decrease θ further.
The impact of harvesting can be drastic for populations that are also subject to Allee effects. Population extinction is certain if harvesting decreases the maximal growth rate of the population to below one even if the population distribution seems large from observation. In addition to the possibility of global extinction, harvesting plays a role on the magnitude of X
1*. Increasing h
i
will increase X
1* so that the weighted size in Γ is larger. However, this deleterious effect of harvesting can be balanced out if the population can reproduce at a younger age. Since life history parameters are not easy to estimate, harvesting without regulations may impose risk for population extinction especially when there is a mechanism of Allee effects operating in the population. Any resource management plan should take Allee effects into consideration before it is implemented.
Related Research Data
References
- Allee , W. C. 1938 . The Social Life of Animals , New Hampshire, USA : William Heinemann .
- Allen , L. 2006 . An Introduction to Mathematical Biology , New Jersey, USA : Prentice-Hall .
- Conway , J. B. 1978 . Functions of One Complex Variable , 2 , New York : Springer .
- Courchamp , F. , Berec , L. and Gascoigne , J. 2008 . Allee Effects in Ecology and Conservation , New York : Oxford University Press .
- Cushing , J. M. 1988 . The Allee effect in age-structured population dynamics, in Mathematical Ecology , Edited by: Hallam , T. , Gross , L. and Levin , S. 479 – 505 . New Jersey, USA : World Scientific .
- Cushing , J. M. 1998 . An Introduction to Structured Population Models , Vol. 71 , Philadelphia : SIAM . Conference Series in Applied Mathematics
- Dennis , B. 1989 . Allee effects, population growth, critical density, and the chance of extinction . Nat. Resour. Model. , 3 : 481 – 538 .
- Dennis , B. 2002 . Allee effects in stochastic populations . Oikos , 96 : 389 – 401 .
- Deredec , A. and Courchamp , F. 2006 . Combined impacts of Allee effects and parasitism . Oikos , 112 : 67 – 679 .
- Grevstad , F. S. 1999 . Experimental invasions using biological control introductions: The influence of release size on the change of population establishment . Biol. Invasions , 1 : 313 – 323 .
- Hilker , F. M. , Langlais , M. , Petrovskii , S. V. and Malcho , H. 2007 . A diffusive SI model with Allee effect and application to FIV . Math. Biosci. , 206 : 61 – 80 .
- Jang , S. R.-J. 2006 . Allee effects in a discrete-time host-parasitoid model . J. Difference Equ. Appl. , 12 : 165 – 181 .
- Jang , S. R.-J. 2007 . Allee effects in a discrete-time host-parasitoid model with stage structure in the host . Discrete Contin. Dyn. Syst. Ser. B , 8 : 145 – 159 .
- Jang , S. R.-J. Discrete-time host-parasitoid models with Allee effects: density dependence versus parasitism . J. Difference Equ. Appl. , (to appear)
- Jang , S. R.-J. Discrete host-parasitoid models with Allee effects and age structure in the host . Math. Biosci. Eng. , (to appear)
- Jang , S. R.-J. and Diamond , S. L. 2007 . A host-parasitoid interaction with Allee effects on the host . Comput. Math. Appl. , 53 : 89 – 103 .
- Morozov , A. , Petrovskii , S. and Li , B.-L. 2004 . Bifurcations and chaos in a predator-prey system with the Allee effect . Proc. R. Soc. Ser. B , 271 : 1407 – 1414 .
- Neimark , Y. 1959 . On some cases of periodic motions depending on parameters . Dokl. Akad. Nauk. SSSR , 129 : 736 – 739 .
- Sacker , R. 1965 . A new approach to the perturbation theory of invariant surfaces . Comm. Pure Appl. Math. , 18 : 717 – 732 .
- Thieme , H. R. , Dhirasakdanon , T. , Han , Z. and Trevino , R. 2009 . Species decline and extinction: synergy of infectious diseases and Allee effect? . J. Biol. Dyn. , 3 : 305 – 323 .
- Wang , X. 2004 . A simple proof of Descartes's rule of signs . Amer. Math. Monthly , 111 : 525 – 526 .
- Wiggins , S. 2003 . Introduction to Applied Nonlinear Dynamical Systems and Chaos , 2 , New York : Springer .
- Zhou , S. and Wang , G. 2004 . Allee-like effects in metapopulation dynamics . Math. Biosci. , 189 : 103 – 113 .
- Zhou , S. , Liu , Y. and Wang , G. 2005 . The stability of predator-prey systems subject to the Allee effects . Theo. Pop. Biol. , 67 : 23 – 31 .