Abstract
We analyse a selection–mutation size-structured model with n ecotypes competing for common resources. Uniform persistence and robust uniform persistence are established, when the selection–mutation matrix Γ is irreducible, i.e. individuals of one ecotype may contribute directly or indirectly to individuals of other ecotypes. Similar results are also presented for a particular reducible form of Γ. In the case of pure selection in which the offspring of one ecotype belong to the same ecotype, i.e. Γ=I, the identity matrix, we prove that the boundary equilibrium that describes competitive exclusion, with the fittest being the winner ecotype, is globally asymptotically stable. We show that small perturbations of the pure selection matrix lead to the existence of globally asymptotically stable interior equilibria. For the case when the selection–mutation matrix is reducible, we present and discuss the outcome of a series of numerical simulations.
1. Introduction
There is a large and growing body of literature on continuous structured population models which assume that individuals are distinguished from one another by characteristics such as size, age or spatial position(e.g. Citation3 Citation7 Citation11 Citation13 Citation14 Citation16 Citation17 Citation18 Citation20 Citation22 Citation33). Continuous structured models have been successfully used to study many biological problems. For example, such models have been used to understand the dynamical behaviour of red coral (Corallium rubrum, L.) growth Citation20 and tumour growth Citation33. The authors in Citation7 derived a model that describes the dynamics of an amphibian population. In Citation17, age-size structured models have been developed to describe a cell population. A population model that incorporates age, size and spatial structure has been studied in Citation33.
Selection–mutation models have been extensively studied in the literature (e.g. Citation2 Citation9 Citation19). These are models that describe the dynamics of a population with respect to some discrete or continuous evolutionary trait. The pure selection process describes the faithful reproduction, i.e. individuals of one ecotype (having the same trait) reproduce individuals of the same ecotype, while mutation describes unfaithful reproduction.
Recently, several researchers have devoted attention to integrating selection–mutation models with continuous size-structured models. The resulting models are partial differential equation systems which assume that differences between individuals are not only due to differences in size and/or age, but also are due to individuals belonging to different ecotypes. The reproduction process in these models is assumed to be open (mutation occurs) or closed (pure selection). In Citation1 Citation4 Citation5 Citation6, nonlinear selection–mutation size-structured models that describe the dynamics of a population with n competing ecotypes are studied. In Citation4, a finite difference scheme to approximate the solution is developed and the convergence of the approximation to the unique bounded variation solution of the model is proved. In Citation5, the authors show, using a contraction mapping argument, that a unique solution of the model exists for all positive time. Conditions on the individual rates which guarantee competitive exclusion in the pure selection case are provided. In the case of an irreducible selection–mutation matrix where individuals of one subpopulation contribute offspring, either directly or indirectly, to all the other subpopulations, the authors proved that all ecotypes coexist.
Ackleh and Deng Citation1 formulate a quasi-linear PDE model for a population consisting of n ecotypes competing for common resources, but they reduce it to a 2n-dimensional ODE. Working with the ODE, the authors consider first the pure selection case, i.e. Γ=I. They prove that competitive exclusion among the ecotypes occurs and provide conditions which determine the fittest ecotype (the one that wins the competition). Second, they consider the case when the selection–mutation matrix is irreducible and prove coexistence of all ecotypes under a very restrictive condition on the model parameters.
Here, using theoretical tools from the persistence theory, we extend the results in Ackleh and Deng Citation1. The main novelties of our work are: Equation(1) when the selection–mutation matrix Γ is irreducible and the extinction equilibrium is unstable in the linear approximation, we obtain coexistence of all ecotypes in the form of uniform persistence. As a consequence, we get the existence of interior equilibria; Equation(2)
we obtain a similar persistence result for a special form of a reducible matrix Γ; Equation(3)
we prove that the persistence is uniform with respect to small changes in the parameters (i.e. the persistence is robust); Equation(4)
we show that irreducible perturbations of the pure selection matrix lead to existence of globally asymptotically stable interior equilibria.
Our paper is organized as follows. In Section 2, we give a brief presentation of the model in Ackleh and Deng Citation1. Our main results are in Section 2. In particular, in Section 3, we give conditions for the coexistence of all ecotypes in the form of uniform persistence when Γ is irreducible; in the case Γ is reducible of a particular form, we provide conditions under which some ecotypes coexist. In Section 3.2, the uniform persistence is extended to robust uniform persistence. In Section 3.3, we show that, in the pure selection case, the boundary equilibrium which describes the survival of the fittest is globally asymptotically stable. This leads to globally asymptotically stable interior equilibria, by perturbing the pure selection matrix. In Section 4, we perform numerical simulations for a model with reducible selection–mutation matrices. Finally, we discuss our conclusions and future work in Section 5. Additional mathematical tools, such as definitions, notation and results that we use throughout the paper can be found in the Appendix.
2. The model
Here, we present the model considered in Citation1, for the dynamics of n ecotypes competing for common resources. We reproduce this model below, but slightly change the notation (the total population size at time t, which is denoted by P(t) in Citation1, is now N(t)) in order to avoid confusion with other notation that we use in this paper.
As in Citation1 Citation11 Citation22, we assume the following submodels for the growth and reproduction rates for each subpopulation
For this choice of growth, reproduction and mortality rates, integrating the first equation of Equation(1) (with respect to x∈[0, 1]), we obtain the differential equation for
and then multiplying the first equation of Equation(1)
by x, and integrating it (again, with respect to x), we obtain the differential equation for
. Thus, we have the following system of coupled ordinary differential equations
3. Main results
All the analysis in this section is with respect to EquationEquation (2). Notation and other results used in the proofs are contained in Section 3. In order to be consistent with the biological interpretations of P
i
s and Q
i
s, we consider our state space to be
(i.e.
). Notice that
whenever
, hence X is positively invariant. We make the following assumption.
-
(H1) f i is a continuously differentiable non-increasing function, f i (0)>0, and m i is a continuously differentiable increasing function, i=1, …, n. We also assume that equation
has a (unique) positive root for some i, and for each j≠i either it has a positive root, orfor all y≥0.
3.1. Coexistence of ecotypes
By setting , the model Equation(2)
can be written in the form
Lemma 3.1
If Γ is irreducible then so is A.
Proof
Suppose that A is reducible. Then there exists a partition J∪K of such that for all j∈J and k∈K, a
jk
=0. Note that
for 1≤i≤n, so n+i and i are either both in J or both in K. Suppose, without loss of generality, that
and
, where m<n. So a
jk
=0 for all
and
. But this contradicts the assumption that Γ is irreducible. Hence, A is irreducible. ▪
The next result establishes dissipativity for our model.
Lemma 3.2
Assume that (H1) holds. Then there exists R>0 such that
is positively invariant and attracts all solutions of Equation
Equation(2)
. Hence Equation
Equation(2)
is dissipative.
Proof
Since Q i ≤P i , we find by changing the order of summation that
As dissipativity plays an important role in our subsequent results, we will just assume that (H1) holds hereafter.
Theorem 3.3
If Γ is irreducible and s(A)>0, then there exists
such that, for all non-zero solutions of Equation
Equation(2)
, there holds
Proof
Let and
. Suppose there exists
. Then for such an x, let x(t) be the solution with x(0)=x. Considering the form Equation(5)
of EquationEquation (2)
and using that A(x(t)) is quasi-positive and irreducible, for all t≥0, from [Theorem 1.1]Citation29 we have that x(t)≫0, for all t>0. Hence X
0={0}. Next, we want to apply Theorem A.2 to conclude EquationEquation (7)
. Lemma 3.2 shows that EquationEquation (2)
is dissipative, which implies that it has a global attractor of points (the result that can be found in [Chapter 1]Citation27). Conditions in 2(a) and 2(b) in Theorem A.2 are clearly satisfied by taking S=X
0={0}, while conditions in 2(c) and 2(d) follow if we show that {0} is a uniformly weak repeller (see Appendix). For this, we make use of Lemma A.4, where we take M={0} and regard {0} as a periodic orbit of period T=1. P(t, 0), the fundamental matrix of solutions for u′(t)=A u(t) (see Appendix), is e
t A
. Hence
, which is a primitive matrix, because A is irreducible (in fact, e
A
has all entries positive, as mentioned in the proof of Theorem A.12.(i) of Citation29). The spectral radius of e
A
is e
s(A), hence it is greater than one, according to our hypothesis that s(A)>0. Then, from Lemma A.4, we obtain that {0} is a uniformly weak repeller, which concludes our proof. ▪
Corollary 3.4
There is an equilibrium point with all coordinates positive.
Proof
Let B be the set guaranteed by Lemma 3.2. Then B is convex (i.e. B contains the entire line segment connecting any two points in B) and positively invariant. From Theorem 3.3, there exists an
such that EquationEquation (7)
holds. Thus, if we take
we have that [Btilde] is compact and convex and, without loss of generality, we can assume it to be positively invariant. Then, from [Theorem 4.8]Citation32, [Btilde] contains an equilibrium. ▪
In our persistence result presented in Theorem 3.3 above, we took advantage of the fact that the set where the persistence function evaluated along an orbit is always zero consisted only of the trivial state {0}. This simplification allowed us to formulate an explicit sufficient condition for uniform persistence of all ecotypes, namely for the extinction equilibrium to be unstable in the linear approximation. A similar situation occurs when the fertility matrix Γ is reducible, but of a particular form, case that we present below. Consider matrix Γ of the form
Let
Theorem 3.5
If Γ1
is irreducible and s(A
1(0))>0, then there exists
such that, for all non-zero solutions of Equation
Equation(2)
, there holds
Proof
Let ,
and x=(x
1, x
2). Let
. Let
be a solution to EquationEquation (2)
. First we show that
Our next goal is to study the case of small mutations of the pure selection model which has been addressed by many researchers to help understand some aspects of adaptive dynamics Citation8 Citation9 Citation10 Citation12 Citation15 Citation21 Citation23. The authors in Citation8 Citation9 show that the non-trivial equilibria of selection–mutation models tend to concentrate at the evolutionary stable strategy (ESS) when the mutation rate or the size of the mutation is small. Furthermore, the study of small mutations has proved to be relevant in the treatment of diseases, such as HIV and influenza, where mutations have a profound impact on therapy Citation21 Citation23.
To achieve our goal and investigate the dynamics of the model with small mutation, we take advantage of the perturbation result developed in Citation28. In order to apply this perturbation result, we will first need to establish (in Section 3.2) robust persistence, and then prove (in Section 3.3) that the steady state of the pure selection model (unperturbed system) is globally asymptotically stable.
3.2. Robust uniform persistence
In this section, we address the question of robust uniform persistence, that is, persistence that is uniform with respect to small changes in the parameters involved in the model.
Let be a compact set, for some positive integer k. We regard Λ as the parameter space and assume that the right-hand side of EquationEquation (2)
is continuous in
. Let
be the semiflow generated by EquationEquation (2)
and A
λ be A=A(0), both corresponding to the parameter
. Hence the map
is continuous. We often write
for
.
For simplicity, we present here only the robust persistence result for an irreducible fertility matrix corresponding to parameter λ0, but a similar result for the case when
is of the form Equation(8)
can be obtained in a completely analogous fashion.
Theorem 3.6
Let
and assume that the corresponding matrix
is irreducible and
. Then
Proof
From Lemma 3.2, it follows that for every there is a
such that
-
(C)
3.3. Globally asymptotically stable interior equilibria
In this section, we consider to be a fixed parameter, for which the corresponding
is the identity matrix. For convenience, we re-order the equations in Equation(2)
such that the equations for P
1′ and Q
1′ are the first two equations, the equations for P
2′ and Q
2′ the next two, and so on. When Γ=I, the system of differential Equationequations (2)
becomes
-
(H2)
Thus, the population corresponding to i=1 turns out to be the ‘fittest’ ecotype (i.e. the ecotype that wins the competition), as shown in [Theorem 5, Theorem 6]Citation1, but under slightly stronger assumptions than (H2). However, with minor modifications in the argument used in Citation1, one can show, under hypothesis (H2), that E
1 is an equilibrium that attracts all solutions of EquationEquation (2) starting with P
1(0)>0. Moreover, the next lemma shows that E
1 is globally asymptotically stable in
–result that will be used later to prove that small perturbations of the fertility matrix
lead to globally asymptotically stable interior equilibria.
Lemma 3.7
If (H2) holds, then
is globally asymptotically stable in
.
Proof
As mentioned above, the proof for E
1 attracting all solutions in is analogous to the proofs in [Theorems 5 and 6]Citation1 and we will omit it here. Next we show that E
1 is asymptotically stable. Evaluating the Jacobian of EquationEquation (25)
at E
1, we find that E
1 is asymptotically stable in the linear approximation if and only if matrices
The next result, used in the proof of Theorem 3.9 below, shows that each neighbourhood of E 1 attracts all initial conditions, uniformly in λ ‘close’ to λ0.
Lemma 3.8
For each
a neighbourhood of E
1
there exists Λ0
a neighbourhood of λ0
such that
Proof
We argue by contradiction: suppose that there is a neighbourhood of E
1 such that, for every neighbourhood
of λ0, there exists
and
with the property that for each T>0,
for some t≥T. Consequently, there exist sequences
and
with
as
such that
Theorem 3.9
There exists
an ϵ-neighbourhood of λ0
in Λ such that for every λ in
there is an equilibrium point E
λ
corresponding to
that is globally asymptotically stable in
. In particular, λ can be chosen so that E
λ
has all components positive.
Proof
Let . Let
be a bounded neighbourhood of E
1 and let Λ0 be a neighbourhood of λ0 granted by Lemma 3.8. We restrict our parameter space to Λ0 (that is, take
). Then, applying [Corollary 2.3]Citation28 in connection to Lemmas 3.7 and 3.8, we obtain that here exists
an ϵ-neighbourhood of λ0 in Λ such that for every λ in
there is an equilibrium point E
λ corresponding to
, such that
, for all x in X
1. We can assume that ϵ is sufficiently small such that, by using that
has all eigenvalues with negative real parts (see Lemma 3.7), to have E
λ be asymptotically stable, hence it is globally asymptotically stable in X
1. Now each eigenvalue of the Jacobian of EquationEquation (2)
evaluated at zero corresponding to λ0 (i.e. each eigenvalue of
) is also an eigenvalue of
4. Numerical simulations
In the previous section, the dynamical behaviour of the system when Γ is irreducible is well studied, but the behaviour of the model in the case Γ is reducible is not completely clear yet. So, in this section, we present numerical simulations of system Equation(2) when Γ is reducible. In Citation2, the authors considered a selection–mutation model (without any size-structure) with constant birth rate and linear death rate functions. They proved that if Γ is completely reducible, competitive exclusion occurs while in the case Γ is reducible but not completely reducible, competitive exclusion and coexistence are possible outcomes.
Our main objective in this section is to numerically investigate the dynamical behaviour of model Equation(2) when Γ is reducible and with birth rates not all being constants and death rates not all being linear (cf. Citation2). To this end, we consider a population with four different ecotypes (phenotypes). We denote by P
i
the number of individuals carrying phenotype i, i=1, 2, 3, 4, at time t. We choose the following values of parameters in model Equation(2)
:
and
. As for the birth and death functions, we let
and
. The above parameters are merely chosen to illustrate the dynamical outcome of the model when Γ is reducible.
We focus on the following two cases of reducible Γ (cf. Citation2): Equation(1) completely reducible Equation(2)
reducible but not completely reducible. We fix all the above chosen parameters and vary c in model Equation(2)
to investigate the influence of c on the dynamics of P
1, P
2, P
3 and P
4. Note that, as c increases, the mortality of individuals carrying phenotype 3 increases and thus the fitness of these individuals decreases.
We begin with the first case. Assume that Group A consists of individuals of ecotypes 1 and 2 (i.e. carrying phenotypes 1 and 2) and that the offspring of these two ecotypes belong to Group A. Similarly assume that Group B consists of individuals of ecotype 3 and 4 and that offspring of Group B belong to Group B. We describe such a scenario using the following completely reducible selection–mutation matrix:
Figure 1. The dynamical behaviour of P 1, P 2, P 3 and P 4 with respect to time and parameter c, when Γ is completely reducible.
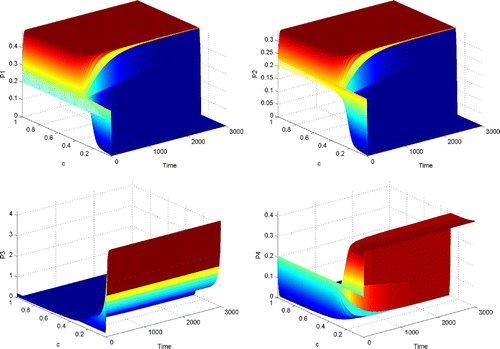
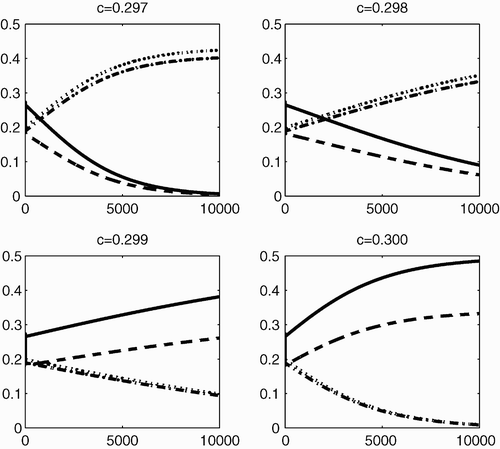
Figure 2. The dynamical behaviour of P 1 (solid line), P 2 (dashed line), P 3 (dotted line) and P 4 (dashed-dotted line) with respect to time, for different values of c (0.297, 0.298, 0.299 and 0.300), when Γ is completely reducible.
Next we will investigate the dynamical behaviour of model Equation(2) when Γ is reducible but not completely reducible. Still consider the above example of a population with individuals belonging to one of four different ecotypes (carrying one of four phenotypes). Suppose again that ecotypes 1 and 2 are in Group A and ecotypes 3 and 4 are in Group B. As before, offspring of individuals in Group A belong to Group A. On the other hand, assume that individuals of Group B have a probability of producing offspring in Groups A and B. We describe such a scenario using the following reducible selection–mutation matrix:
From and , we can see that when Γ is reducible but not completely reducible, competitive exclusion and coexistence are possible outcomes of the dynamical behaviour of system Equation(2). When the value of c is small (less than 0.3), coexistence among P
1, P
2, P
3 and P
4 occurs. As the value of c increases above 0.3, the populations of P
3 and P
4 go extinct while P
1 and P
2 survive. Thus competitive exclusion occurs.
Figure 3. The dynamical behaviour of P 1, P 2, P 3 and P 4 with respect to time and parameter c, when Γ is reducible but not completely reducible.
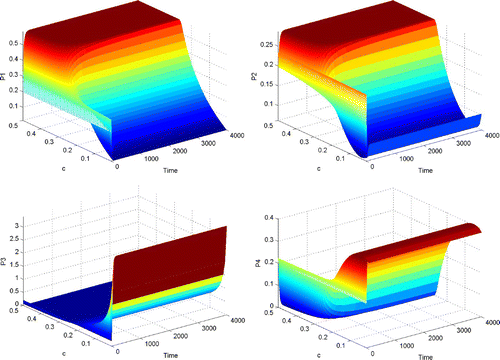
Figure 4. The dynamical behaviour of P 1 (solid line), P 2 (dashed line), P 3 (dotted line) and P 4 (dashed-dotted line) with respect to time, for different values of c (0.10, 0.15, 0.20, 0.25, 0.30 and 0.35), when Γ is reducible but not completely reducible.
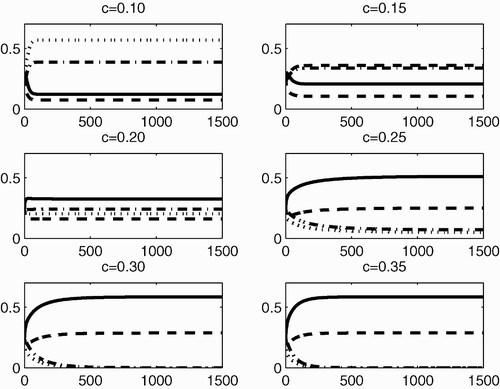
To interpret this biologically, again note that the smaller the value of c the more fit ecotype 3 is, leading to a higher fitness in Group B. Thus, Group B survives and since individuals of Group B produce individuals of Group A, this leads to Group A surviving and hence coexistence occurs. On the other hand, when c becomes large, the fitness of Group A exceeds that of Group B. Thus, Group A survives and Group B goes extinct (since individuals of the more fit Group A do not produce individuals of Group B).
5. Conclusions and future work
In this paper, we analysed a selection–mutation size-structured model where the population consists of n ecotypes. We showed that, in the case of an irreducible selection–mutation matrix Γ (i.e. when individuals of one subpopulation contribute directly or indirectly to individuals of another subpopulation), the origin is a uniformly weak repeller (see Definition A.3) and all ecotypes coexist in the form of robust uniform persistence. In the case of the pure selection matrix Γ=I (the offspring of one ecotype belong to the same ecotype) the authors in Citation1 proved that competitive exclusion occurs: the fittest subpopulation survives and all other subpopulations die out. We extended this result and showed, under less restrictive assumptions, that the boundary globally attracting equilibrium corresponding to the fittest ecotype is globally asymptotically stable. Moreover, for Γ irreducible, but ‘close’ to the identity matrix, the model Equation(2) has a globally asymptotically stable interior equilibrium.
Our numerical simulations suggest that, in the case when Γ is completely reducible, competitive exclusion is the most probable outcome, while in the case when Γ is reducible, but not completely reducible, either competitive exclusion, or coexistence are possible outcomes. Our future efforts will focus on theoretically studying the dynamical behaviour in the reducible case. In particular, we are interested in finding conditions that determine which ecotypes persist and which go extinct.
Furthermore, our numerical results suggest that even in the case of reducible or completely reducible matrix Γ, the total population persists (i.e. at least one ecotype survives). Therefore, we conjecture that Theorem 3.3 can be established by dropping the assumption on irreducibility. In fact, we discussed this case, but only for a particular form of Γ, as given in EquationEquation (8). As we already mentioned, more general cases can be treated by using similar ideas, but then explicit conditions for uniform persistence are improbable to obtain, as one needs information about the dynamics on the boundary corresponding to the extinction states, and this dynamics could be quite complicated.
In our model Equation(2), k
i
∈(0, 1) is a fraction of ingested food that is channelled to growth and maintenance and (1−k
i
) is the fraction channelled to reproduction. Throughout our discussion, we assumed that each
, is constant. But many populations reproduce seasonally and thus in many cases, k
i
are better represented by periodic functions of t. In our future work, we plan to study this periodic version of system Equation(2)
.
Acknowledgements
The authors would like to the thank an anonymous referee for his valuable comments and in particular for suggesting Theorem 3.5. The research of A.S. Ackleh and Baoling Ma is supported in part by the National Science Foundation under grant # DMS-0718465.
References
- Ackleh , A. S. and Deng , K. 2004 . Survival of the fittest in a quasilinear size-structured population model . Natur. Resource Modeling , 17 : 213 – 228 .
- Ackleh , A. S. and Hu , S. 2007 . Comparison between stochastic and deterministic selection–mutation models . Math. Biosci. Eng. , 4 : 133 – 157 .
- Ackleh , A. S. and Ito , K. 1997 . An implicit finite difference scheme for the nonlinear size-structured population model . Numer. Funct. Anal. Optim. , 18 : 865 – 884 .
- Ackleh , A. S. , Banks , H. T. and Keng Deng . 2002 . A finite difference approximation for a coupled system of nonlinear size-structured populations . Nonlinear Anal. , 50 : 727 – 748 .
- Ackleh , A. S. , Deng , K. and Wang , X. 2004 . Competitive exclusion and coexistence for a quasilinear size-structured population model . Math. Biosci. , 192 : 177 – 192 .
- Ackleh , A. S. , Banks , H. T. , Deng , K. and Hu , S. 2005 . Parameter estimation in a coupled system of nonlinear size-structured populations . Math. Biosci. Eng. , 2 : 289 – 315 .
- Ackleh , A. S. , Deng , K. and Huang , Q. 2010 . Existence-uniqueness results and difference approximations for an amphibian juvenile-adult model . Contemp. Math. , 513 : 1 – 23 .
- Calsina , À. and Cuadrado , S. 2004 . Small mutation rate and evolutionarily stable strategies in infinite dimensional adaptive dynamics . J. Math. Biol. , 48 : 135 – 159 .
- Calsina , À. and Cuadrado , S. 2005 . Stationary solutions of a selection mutation model: The pure mutation case . Math. Models Methods Appl. Sci. , 15 : 1091 – 1117 .
- Calsina , À. and Cuadrado , S. 2007 . Asymptotic stability of equilibria of selection–mutation equations . J. Math. Biol. , 54 : 489 – 511 .
- Calsina , À and Saldaña , J. 1995 . A model of physiologically structured population dynamics with a nonlinear individual growth rate . J. Math. Biol. , 33 : 335 – 364 .
- Cuadrado , S. 2009 . Stability of equilibria of a predator–prey model of phenotype evolution . Math. Biosci. Eng. , 6 : 701 – 718 .
- Cushing , J. M. 1990 . Some competition models for size-structured populations . Rocky Mountain J. Math. , 20 : 879 – 897 .
- Cushing , J. M. 1998 . An Introduction to Structured Population Dynamics , Philadelphia : SIAM .
- Diekmann , O. , Jabin , P. E. , Mischler , S. and Perthame , B. 2005 . The dynamics of adaptation: An illuminating example and a Hamilton–Jacobi approach . Theoret. Popul. Biol. , 67 : 257 – 271 .
- Farkas , J. Z. and Hagen , T. 2009 . Asymptotic analysis of a size-structured cannibalism model with infinite dimensional environmental feedback . Commun. Pure Appl. Anal. , 8 : 1825 – 1839 .
- Heijmans , H. 1986 . “ The dynamical behavior of the age-size distribution of a cell population ” . In The Dynamics of Physiologically Structured Populations , Edited by: Metz , J. A.J. and Diekmann , O. 185 – 202 . Berlin : Springer . Lecture Notes in Biomathematics Vol. 68
- Henson , S. M. and Hallam , T. G. 1994 . Survival of the fittest: Asymptotic competitive exclusion in structured population and community models . Nonlinear World , 1 : 385 – 402 .
- Hofbauer , J. and Sigmund , K. 2003 . Evolutionary game dynamics . Bull. Am. Math. Soc. , 40 : 479 – 519 .
- Jabin , P.-E. , Lemesle , V. and Aurelle , D. 2008 . A continuous size-structured red coral growth model . Math. Models Methods Appl. Sci. , 18 : 1927 – 1944 .
- Leenheer , P. De. and Pilyugin , S. S. 2008 . Multistrain virus dynamics with mutations: A global analysis . Math. Med. Biol. , 25 : 285 – 322 .
- Metz , J. A.J. and Diekmann , O. 1986 . The Dynamics of Physiologically Structured Populations , Berlin : Springer . Lecture Notes in Biomathematics Vol. 68
- Rong , L. , Feng , Z. and Perelson , A. S. 2007 . Emergence of HIV-1 drug resistance during antiretroviral treatment . Bull. Math. Biol. , 69 : 2027 – 2060 .
- Salceanu , P. L. 2010 . “ Lyapunov exponents and persistence in dynamical systems with applications to some discrete time models ” . Proquest Dissertations and Theses 2010, 110 pages, Ph.D. diss., Arizona State University, Arizona, USA, 2009. Publication Number: AAT 3371233. Source: DAI-B 70/08, February
- Salceanu , P. L. and Smith , H. L. 2009 . Lyapunov exponents and persistence in discrete dynamical systems . Discrete Contin. Dyn. Syst. B , 12 : 187 – 203 .
- Salceanu , P. L. and Smith , H. L. Lyapunov exponents and uniform weak normally repelling invariant sets . Proceedings of the third Multidisciplinary International Symposium on Positive Systems: Theory and Applications (POSTA 2009) . September , Valencia, Spain. pp. 17 – 27 . Lecture Notes in Control and Informational Sciences
- Smith , H. L. and Thieme , H. 2011 . Dynamical Systems and Population Persistence , Providence, R.I : American Mathematical Society .
- Smith , H. L. and Waltman , P. 1999 . Perturbation of a globally stable steady state . Proc. Am. Math. Soc. , 127 : 447 – 453 .
- Smith , H. L. and Waltman , P. 2003 . The Theory of the Chemostat , New York : Cambridge University Press .
- Smith , H. L. and Zhao , X.-Q. 2001 . Robust persistence for semi-dynamical systems . Nonlinear Anal. , 47 : 6169 – 6179 .
- Thieme , R. H. 2003 . Mathematics in Population Biology , NJ : Princeton University Press .
- Verhulst , F. 1990 . Nonlinear Differential Equations and Dynamical Systems , New York : Springer .
- Webb , G. F. 2008 . “ Population models structured by age, size and spatial position ” . In Structured Population Models in Biology and Epidemiology , Edited by: Magal , P. and Ruan , S. 1 – 49 . Berlin, Heidelberg : Springer .
- Zhao , X.-Q. 2003 . Dynamical Systems in Population Biology , New York : Springer .
Appendix
Let be the non-negative cone in ℝ
m
. By abusing notation, we define the norm of
to be
, where |x
i
| represents the absolute value of x
i
. Let d be the distance induced by this norm, i.e.
. S¯ denotes the closure of
.
We call a matrix A non-negative (strictly positive), and write, A≥0 (A≫0), if each entry of A is a non-negative (positive) number. We call A positive, and write A>0, if A≥0, but A is not the zero matrix. Assume analogous definitions (and notation) for vectors.
Denote by s(A) and r(A) the stability modulus and the spectral radius of a matrix A, respectively. I denotes the identity matrix (whose dimension will be clear from the context).
Consider a differential equation
Let be continuous and not identically zero. Let
and
, for some set
, where ω(x) represents the omega limit set of x. A set S is said to be positively invariant (with respect to φ) if x∈S implies
,
.
Definition A.1
The system (A1) is called uniformly (strongly) ρ-persistent if there is an
such that
, whenever
.
Usually ρ is referred to as the ‘persistence function’. Also, one may omit the word ‘strongly’ and just say ‘uniformly persistent’.
The next theorem, which is a particular case of Theorem 3 in Citation30, is well known in persistence theory and is one of the main tools used for our persistence results. Detailed explanation of the terminology used in this theorem (e.g. invariant set, isolated set, etc.) can be found, for example, in Citation27 Citation30 Citation34.
Theorem A.2
Assume that the following hold:
-
(1) φ has a global attractor (of points).
-
(2) There exists a finite sequence
-
(a)
-
(b) S is acyclic;
-
(c) S i is isolated in
-
(d)
-
W
S
(S
i
) above denotes the stable manifold of the set S
i
, that is, , where
.
However, verifying conditions (c) and (d) in the above theorem can be quite a task, even when the sets S i are ‘simple’, such as equilibria or periodic orbits. If S i s are uniformly weak repellers (see definition below) then these conditions are satisfied, with the additional requirement (for c)) that S i be asymptotically stable in X 0. Characterization of such boundary attractors as uniformly weak repellers can be found in Citation24, or in Citation25 Citation26 (for discrete time).
Definition A.3
A non-empty set
is called a uniformly weak repeller if there exists
such that
.
A square non-negative matrix A is called: Equation(1)
reducible, if it can be set in the form
Assume that x(t) satisfying Equation Equation(A1) can be written as
, where
and A(x) is a continuous matrix function such that A(x
1, 0) is quasipositive (it has all off-diagonal entries non-negative). In this setup, x
1(t) could be an empty vector, in which case x
2(t)=x(t). Assume that the set
, as well as its complement, is positively invariant for Equation Equation(A1)
. Let M be a compact and positively invariant set contained in
. Let P(t, x) denote the fundamental matrix of solutions for
Given a periodic orbit 𝒫 of Equation Equation(A1), of period T>0, r(P(T, x)) has the same value for all
(see [Lemma 3.10]Citation24). Denote this common value by
. Next result, reproduced here (in a simplified form) from Citation24 ([Corollary 3.12]Citation24), is especially useful in the particular case when the boundary attractor consists of periodic orbits.
Lemma A.4
Assume that Ω(M) is a union of periodic orbits and the following hold:
-
(1)
-
(2)