Abstract
We consider a population model consisting of d species interacting in a p-periodic environment and modelled by a d-dimensional system of Leslie–Gower-type difference equations (coupled Beverton–Holt equations). It is shown that if the interspecific competition (coupling) is sufficiently small and the inherent growth rate of each species is such that in the absence of competition each species will grow to its (positive) individual carrying capacity, then there is a positive asymptotically stable p-periodic state that globally attracts all positive initial states. Three examples are studied numerically in which the competition is large and the principle of competitive exclusion is observed. The rate of decay to extinction is observed to be sensitive to the inherent growth rate of the dying species. The individual carrying capacities are seen to play a determining role in the case of equal and large competition and equal inherent growth rates.
1. Introduction
The study of competition models inevitably leads one to consider either the Lotka–Volterra model in the continuous case or the Leslie–Gower model Citation10 in the discrete case. The typical Leslie–Gower model consists of two Beverton–Holt equations with added coupling (interspecific competition). When the interspecific competition is strong, one species will be driven to extinction; the principle of competitive exclusion that is one of the important tenets in ecology (see Citation2 Citation3 Citation7 for results and many references to this phenomenon). In Citation11, global stability in a two-species model is considered using techniques of monotone systems Citation9. In Citation4, multi-species models are considered, taking into account harvesting and stocking.
See Citation6 for further results on stage-structured models for larvae, pupae and adults, the well-known ‘LPA’ model that is essentially a delay equation for the larvae and adults. In Citation5, non-equilibrium competitive coexistence for a two-species LPA model was explored and a boundary 2-cycle was established. See also Citation1 in which a two-species juvenile–adult model is studied with the assumption that there is no competition between juveniles and adults.
For the Beverton–Holt (scalar) equation, the issue of global asymptotic stability, even in the periodic case, has been settled in Citation8. Since the functions defining these equations, being fractional linear, form a semi-group ℬ under composition, the existence and global asymptotic stability of a periodic equation reduces to establishing a fixed point with the same property for a single (autonomous) equation. In fact, in Citation8, it was shown that ℬ is a sub-semi-group of the larger semi-group 𝒦 of continuous functions that are concave, increasing and cross the diagonal in
. In Citation12, the result was extended to C
3 functions that are either concave increasing or convex decreasing and have non-negative Schwarzian. These conditions are satisfied by certain rational functions with the roots of the numerator interlaced with the roots of the denominator.
In this paper, we consider d species interacting in a periodic environment modeled by a d-dimensional system of Leslie–Gower-type equations, or equivalently coupled Beverton–Holt equations. It is assumed that the inherent growth rate of each species is such that in the absence of competition each species will grow to its (positive) individual carrying capacity. It is shown that if the interspecific competition (coupling) is sufficiently small, then there is a positive asymptotically stable periodic state that globally attracts all positive initial states.
We then study numerically, in three four-dimensional examples, some cases in which the interspecific competition is large. In the first example, we see that large competition against just species number one, not surprisingly, drives that species to extinction. In the second example, we see that an increase in the inherent growth rate for species one by a factor of 1.77 must be countered by a 4.3-fold increase in the competition by all three competing species in order to achieve the same rate of decay to extinction. In the third example, we make all the competition large and equal and all the inherent growth rates equal and observe that the species with the smallest individual carrying capacity is driven to extinction.
2. Autonomous two-dimensional case
We begin with a discussion of this case in order to develop some notation that will make the d-dimensional case easier to formulate and discuss.
The two-species Leslie–Gower model is usually written in the following form:
We propose the following equivalent form of EquationEquation (1) that is a pair of coupled Beverton–Holt equations. In addition, we view a difference equation as a mapping
and thus focus our attention on the right-hand side f:
Our next goal will be to develop a notation and some operations that will make it straightforward to consider higher-dimensional maps. For those familiar with Matlab programming, these operations will not seem so strange.
Let a be a scalar and u and v be column vectors in ℝ d and C an d×d matrix. Define, in addition to the usual inner product and linearity rules,
To further simplify this and eliminate cumbersome notation, we define the parameters
This form is not specific to ℝ2, but can be interpreted in ℝ
d
as well. With C
0=0, EquationEquation (7) represents d independent Beverton–Holt equations.
3. Interior fixed point, autonomous case: p=1
We consider the positive cone
Theorem 3.1
Assume μ
i
>1 for all i=1, 2, …, d and the coupling terms C
0
are sufficiently small. Then there exists a unique fixed point
that reduces to
when C
0=0.
The theorem applies equally well to any of the invariant coordinate ‘faces’
In order to study stability, we note the following lemma.
Lemma 3.2
The function f is bounded in
. More precisely, each component function
We next define K min and K max to be the minimum and maximum of the K i and b=K min/2. We then have the following lemma.
Lemma 3.3
Assume each row c
i
of the matrix C
0
in Equation
Equation(7)
satisfies
The proof will follow by setting v=x in the next more general lemma needed later.
Lemma 3.4
Define
as follows. Let
be arbitrary and define (cf. Equation
Equation(7)
)
Proof
Let denote the inner product of vectors p and q. For
, the ith component of [fcirc] satisfies
3.1. Dynamic reduction
In the technique of dynamic reduction introduced in Citation14, one defines a class of p-periodic sequences of column vectors v n :
Theorem 3.5
In addition to
assume that the rows of C
0
satisfy
Proof
Each component function [fcirc]
i
in EquationEquation (18) is fractional linear, concave increasing and from EquationEquation (19)
has a slope at the origin that is greater than one. Thus, either by Citation8 or Citation12, one obtains an exponentially asymptotically stable solution (fixed point) w
i
and from Lemma 3.4,
. This establishes EquationEquation (20)
. Note that since w satisfies EquationEquation (18)
,
4. Periodic case: p>1
The periodic version of EquationEquation (7) is
, where
Figure 1. Orbit in skew-product flow.
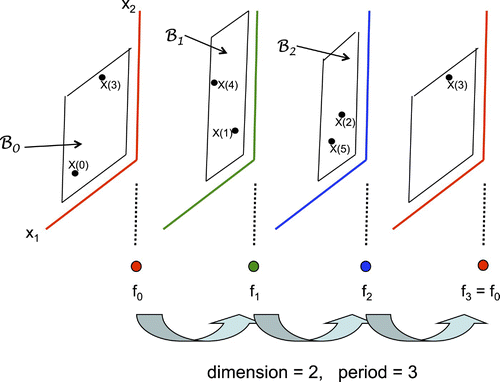
Before proceeding, however, let us prove the periodic version of Lemmas 3.2 and 3.4 in the general case.
Lemma 4.1
The functions f
n
are bounded in
. More precisely, each component function satisfies
Lemma 4.2
For
define
with
Note: The set on the right is labelled with ‘j+1’ since it lies in the domain of f j+1 with subscripts taken mod p ().
Proof
Let denote the inner product of vectors p and q. For
, the ith component of [fcirc] satisfies
4.1. Dynamic reduction: periodic case
To apply dynamic reduction, we first separate out the coupling terms in EquationEquation (21) to obtain
The difference equations on the right-hand sides of EquationEquation (29) are uncoupled and each such equation is a three periodic concave increasing function mapping
. By Citation8
Citation12, we obtain an exponentially asymptotically stable periodic solution,
Figure 2. Mapping 𝒯.
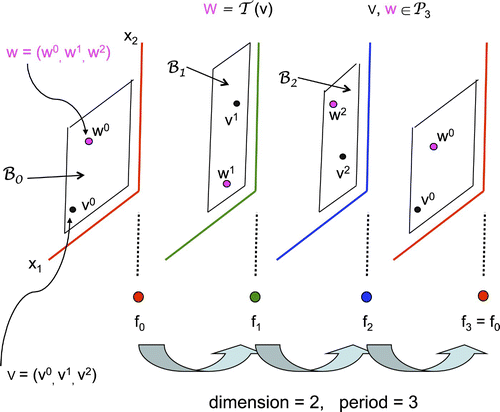
We next introduce the following notation: For and a periodic vector valued sequence of functions
with
we define
. For a fixed
with
, and
with
define
. For F=F(x, g), we will use ∂1
F and ∂2
F to mean differentiation with respect to the first and second arguments.
Referring to EquationEquation (28), let us temporarily suppress the dependency of the F
j
on g and define
Thus, from EquationEquation (31), and recalling that we are taking the period p=3 for simplicity of presentation,
Thus has a bounded inverse and from EquationEquation (32)
,
The global asymptotic stability follows by the argument given in Citation14. Thus, we have established the following theorem.
Theorem 4.3
Consider the p
-periodic, d
-dimensional system
Equation(21)
which we repeat:
where
5. Large inter-specific competition: numerical examples
In the previous sections, we considered the result of small inter-specific competition, i.e. small coupling C
0 in EquationEquation (7) or EquationEquation (34)
in the periodic case. No species was driven to extinction and a coexistence state was established that attracted all initial states starting in the positive cone 𝒞0 defined in EquationEquation (8)
.
We now consider, in dimension 4, the effect of strong competition against species number one, x 1, with inherent growth rates approximately equal in Section 5.1. Then, in Section 5.2, we increase the inherent growth rate for x 1 and note the rather large increase in competition against x 1 needed to achieve the same decay rate to extinction as in case 1. In Section 5.3, we consider the case of equal competition and equal inherent growth rates and see that the individual carrying capacities determine the species that goes extinct.
As a point of reference, let us consider an example of small competition with
Figure 3. Coexistence.
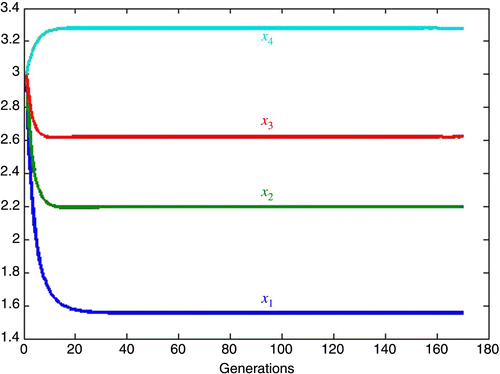
5.1. Inherent growth rates approximately equal
We now consider the example in EquationEquations (35) and Equation(36)
in which the inherent growth rates of all species are approximately equal and increase the competition against species one to
. The attractive fixed point is now (),
Figure 4. Large competition against species 1.
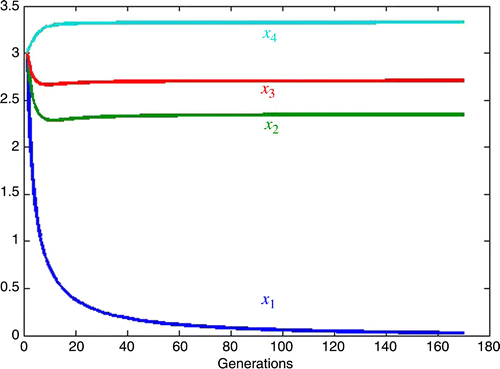
Specifically,
5.2. One dominant inherent growth rate
We now consider the example in EquationEquations (35) and Equation(36)
except that now we take the inherent growth rate of species 1 to be 1.77 times that given in EquationEquation (36)
. Thus, let
For the attractive fixed point is
By increasing the competition against species one by approximately 5.3% to the attractive fixed point remains unchanged to the accuracy shown, but species x
1 approaches extinction much faster, viz.
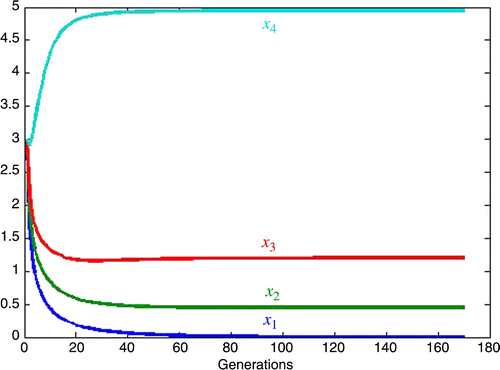
5.3. Equal but large competition
Here we consider the case in which all the competition is equal and the inherent growth rates are equal. As a point of reference, consider the small competition case where in EquationEquation (3),
Figure 5. Extinction determined by carrying capacities.
6. Conclusions
We have studied d species interacting in a p-periodic environment and modeled by a d-dimensional system of Leslie–Gower-type equations (coupled Beverton–Holt equations). It is shown that if the interspecific competition (coupling) is sufficiently small and the inherent growth rate of each species is such that in the absence of competition each species will grow to its (positive) individual carrying capacity, then there is a positive asymptotically stable p-periodic state that globally attracts all positive initial states, i.e. coexistence.
We also study three cases of large competition, all of which lead to competitive exclusion with species one, x 1 going extinct. In the first, we let the competition be unbalanced and discriminating against x 1 with inherent growth rates approximately equal. In case 2, we increase the inherent growth rate for x 1 by a factor of 1.77 and see that the competition against x 1 must be increased by a factor of 4.3 in order to obtain the same rate of decay to extinction as in case 1. In case 3, we set all the competition equal and large and all the inherent growth rates equal and see that the ordering of the size of the species at equilibrium is the same as that of the individual carrying capacities, with the species having the least carrying capacity, x 1, going extinct.
Acknowledgements
Robert J. Sacker is supported by the University of Southern California, Letters Arts and Sciences Faculty Development Grant. The author thanks the referee for their helpful suggestions.
References
- Ackleh , A. S. and Chiquet , R. A. Competitive exclusion in a discrete juvenile–adult model with continuous and seasonal reproduction . preprint (2010), to appear in J. Difference Equ. Appl. doi:10.1080/10236190903460420
- Ackleh , A. S. , Dib , Y. M. and Jang , S. R.J. 2005 . “ A discrete-time Beverton-Holt competition model ” . In Difference Equations and Discrete Dynamical Systems , Edited by: Allen , L. J.S. , Aulbach , B. , Elaydi , S. and Sacker , R. 1 – 9 . London : World Scientific .
- Ackleh , A. S. , Dib , Y. M. and Jang , S. R.J. 2007 . Competitive exclusion and coexistence in a nonlinear refuge-mediated selection model . Discrete Contin. Dyn. Syst. Ser. B , 7 ( 4 ) : 683 – 698 .
- AlSharawi , Z. and Rhouma , M. 2009 . Coexistence and extinction in a competitive exclusion Leslie/Gower model with harvesting and stocking . J. Difference Equ. Appl. , 15 ( 11 ) : 1031 – 1053 .
- Cushing , J. M. , Henson , S. M. and Roeger , L.-I. 2007 . Coexistence of competing juvenile–adult structured populations . J. Biol. Dyn. , 1 ( 2 ) : 201 – 231 .
- Cushing , J. M. and LeVarge , S. 2005 . “ Some discrete competition models and the principle of competitive exclusion ” . In Difference Equations and Discrete Dynamical Systems , Edited by: Allen , L. J.S. , Aulbach , B. , Elaydi , S. and Sacker , R. 283 – 301 . London : World Scientific .
- Cushing , J. M. , Levarge , S. , Chitnis , N. and Henson , S. M. 2004 . Some discrete competition models and the competitive exclusion pronciple . J. Difference Equ. Appl. , 10 ( 13–15 ) : 1139 – 1151 .
- Elaydi , S. and Sacker , R. J. 2005 . Global stability of periodic orbits of nonautonomous difference equations . J. Differential Equations , 208 ( 11 ) : 258 – 273 .
- Hirsch , M. W. and Smith , H. 2005 . Monotone maps: A review . J. Difference Equ. Appl. , 11 ( 4–5 ) : 379 – 398 .
- Leslie , P. H. and Gower , J. C. 1958 . The properties of a stochastic model for two competing species . Biometrika , 45 : 316 – 330 .
- Liu , P. and Elaydi , S. N. 2001 . Discrete competitive and cooperative models of Lotka-Volterra type . J. Comput. Anal. Appl. , 3 ( 1 ) : 53 – 73 .
- Sacker , R. J. 2010 . Semigroups of maps and periodic difference equations . J. Difference Equ. Appl. , 16 ( 1 ) : 1 – 13 .
- Sacker , R. J. and Sell , G. R. 1977 . Lifting properties in skew-product flows with applications to differential equations . Mem. Amer. Math. Soc. , 11 ( 190 )
- Sacker , R. J. and von Bremen , H. F. 2007 . Dynamic reduction with applications to mathematical biology and other areas . J. Biol. Dyn. , 1 ( 4 ) : 437 – 453 .