Abstract
This article introduces a two-strain spatially explicit SIS epidemic model with space-dependent transmission parameters. We define reproduction numbers of the two strains, and show that the disease-free equilibrium will be globally stable if both reproduction numbers are below one. We also introduce the invasion numbers of the two strains which determine the ability of each strain to invade the single-strain equilibrium of the other strain. The main question that we address is whether the presence of spatial structure would allow the two strains to coexist, as the corresponding spatially homogeneous model leads to competitive exclusion. We show analytically that if both invasion numbers are larger than one, then there is a coexistence equilibrium. We devise a finite element numerical method to numerically confirm the stability of the coexistence equilibrium and investigate various competition scenarios between the strains. Finally, we show that the numerical scheme preserves the positive cone and converges of first order in the time variable and second order in the space variables.
AMS Subject Classification:
1. Introduction
Most communicable diseases exhibit variation in epidemiological characteristics throughout space. For instance, different countries have different incidences of a given infectious disease. Key epidemiological parameters, such as transmission and often recovery, are intrinsically spatially dependent. Disease transmission through direct contact is always local, and its intensity can be linked to the spatial location. This suggests that when the classical epidemic models are recast to include diffusion and incorporate spatial heterogeneity explicitly, the transmission coefficients should be functions of the spatial location where the contact occurs. It is surprising to see that most classical epidemiological spatially heterogeneous models have a reaction part where rates are constant with respect to location. Notable exceptions exist. The spatial effects have been widely studied through patch models (e.g. Citation19) or by adding diffusion to an ordinary differential equation (ODE) model Citation25. The authors in Citation19 studied how banning travel of symptomatic or infected individuals from high to low prevalence patch will effect the spread of the disease by considering a multi-patch model where transmission and recovery are patch-dependent.
In three papers Fitzgibbon et al. study a number of directly, environmentally and vector-transmitted diseases where the transmission rate is spatially heterogeneous Citation15 Citation16 Citation17. The authors derive analytical results on well-posedness of the systems as well as various limiting properties of the solutions. Epidemiologically relevant threshold conditions, however, are derived in the case of constant coefficients Citation15.
Studying the classical epidemiological models with spatially heterogeneous coefficients presents significant difficulties. The first article that derives a reproduction number for a simple SIS models with spatially heterogeneous transmission and recovery appears to be the one by Allen et al. Citation2. The reproduction number defined there is not explicit in the sense that if the parameters were known the value may still be not easy to compute. Furthermore, this reproduction number does not seem to follow the next generation approach Citation12 but possesses the usual threshold property. The authors also derive useful epidemiological characteristics such as dependence of the reproduction number on the diffusion coefficients.
In this paper we consider a two-strain extension of the model presented in Citation2. The two strain model does not incorporate any explicit trade-off mechanisms, such as co-infection or super-infection Citation23 that may lead to coexistence of the strains even in the model’s ODE counterpart. The main question that we address is whether space alone could allow for coexistence. We expect and obtain a positive answer to our question. The continuous scenario that we investigate is analogous to a discrete multi-patch model where the disease cannot persist on any of the individual patches, but can persist in the system. The ability of the diseases to persist in a system of sink habitats, that is patches with basic reproduction number smaller than one, has been observed before in various multi-patch epidemiological models (e.g. Citation11).
We approach the problem both analytically and numerically. Analytical studies of multi-strain epidemic models with diffusion are scarce. Kim et al. Citation22 discusses a spatial version of an avian influenza multi-strain model with nonlocal transmission in humans, but the strains there do not compete as each one is established in a different species. Competition has more thoroughly been addressed in ecology in the context of Lotka–Volterra competition models Citation7, p. 224]. In an ecological context, the effect of diffusion on coexistence in competing populations has been studied in multiple scenarios. For instance, Pacala and Roughgarden [Citation25 consider a pair of coupled Lotka–Volterra competition equations with diffusion on one-dimensional spatial habitat. The only spatially heterogeneous parameter is the carrying capacity which is defined as a step function. Furthermore, the effects of spatial heterogeneity on the persistence of two competing species have been investigated in Citation8 where the authors studied the dynamics of the two interacting populations by considering a coupled reaction diffusion system having spatially dependent reaction terms. The impact of space on competing species in an ecological context has much in common with the question we study in this paper.
Here, addressing our main question, we show analytically that a coexistence equilibrium exists. This is the case if the invasion numbers of the two strains, defined analogously to the reproduction numbers, are larger than one. To show stability of the coexistence equilibrium we develop a fully discrete finite element method. Finite element methods are a natural approach to diffusion problems including in epidemiology Citation21. The main challenge is to develop a finite element method that preserves the positivity of the solution. We use the finite element method to demonstrate through simulations the convergence to a coexistence equilibrium as well as to investigate other outcomes of the competition of the strains based on the reproduction numbers and invasion numbers. Since the reproduction numbers and invasion numbers for our examples cannot be computed exactly, even when the parameters of the model are known, we develop a novel numerical method that estimates the relation of these numbers to one.
This paper is structured in the following way: In Section 2 we introduce the two-strain model and the reproduction numbers of the strains. In Section 3 we investigate the local stability of the disease-free equilibrium (DFE) and the semi-trivial equilibria. We also introduce the invasion numbers and their relation to the corresponding principal eigenvalue. In Section 4, we use the monotonicity of the equilibrial system to show the existence of coexistence equilibrium in the case when both invasion numbers are larger than one. In Section 5 we introduce our finite element method. Furthermore, we establish its order of convergence and we show that it preserves the positivity of the solutions. Section 6 is devoted to the numerical computation of the reproduction numbers and invasion numbers. Section 7 shows numerical evidence of the convergence of our finite element method. Furthermore, we investigate through simulations various outcomes of the competition of the strains governed by the reproduction and invasion numbers. Section 8 contains a summary of our results.
2. A two-strain SIS model with diffusion
In this section we introduce a two-strain SIS epidemic model with diffusion. The total population size is divided into nonintersecting classes of susceptibles, infected with strain one and strain two. Let S(x, t) be the density of the susceptible individuals at position x at time t>0. Let I 1(x, t) and I 2(x, t) represent the densities of infected individuals with respect to strain one and two, respectively. We consider the following reaction diffusion system:
This model is a two-strain extension of the model in Citation2. It is clear from the standard theory of semilinear parabolic systems that a unique classical solution (S, I
1, I
2) of (Equation1) exists for all time Citation18. In this model, we assume the diffusion coefficients d
s
, d
1, and d
2 for the susceptible and infected populations to be positive coefficients. Reaction terms, β1(x)(S
I
1/(S+I
1+I
2)) and β2(x)(S
I
2/(S+I
1+I
2)) , give the incidence of the disease, that is, the number of newly infected susceptible individuals per unit of time who have contracted the disease and have become infected at a given position x∈Ω. Here, β1(x), β2(x) are the space-dependent rates of disease transmission by infectives with strain one and strain two, respectively, at position x∈Ω. Infected individuals with strain one and strain two recover and become susceptible with space-dependent rates of recovery γ1(x) and γ2(x) for position x∈Ω. We assume that β1(x), β2(x), γ1(x), and γ2(x) are positive Hölder continuous functions on
. Assume also that
We denote by the basic reproduction number which gives the number of secondary infections that one infected individual will produce in an entirely susceptible population. The basic reproduction number determines whether there is an epidemic or not. The reproduction number of strain i is given by
Then the basic reproduction number is defined as
. We show in the next section that if
, then both strains die out. It is expected that if
, then at least one of the strains persists.
3. Local stability of the DFE and semi-trivial equilibrium
We are interested in the equilibrium solutions of (Equation1), i.e. the time-independent solutions of the following system:
A DFE is a solution of (Equation2) in which both strains
and
vanish at every x∈Ω at equilibrium, i.e.
. A strain-one equilibrium, I
1-equilibrium, is a solution of (Equation2
) in which only strain two,
vanishes at every x∈Ω at equilibrium. Similarly, a strain-two equilibrium, I
2-equilibrium, is a solution of (Equation2
) in which only
vanishes at every x∈Ω at equilibrium. An endemic equilibrium (EE) is a solution in which both strains coexist, i.e.
and
for some x∈Ω. To distinguish between these four types of equilibria, we use different notations for each. Throughout the paper, we denote DFE by
, I
1-equilibrium by
, I
2-equilibrium by
, and EE by
.
Theorem 3.1
There exists a unique DFE and it is given by
Proof
Since in DFE both , by (Equation2
) we have
. With boundary condition
,
implies that
is a constant in Ω. Clearly any constant is a solution, but note that since
and
and
is a constant,
, where |Ω| is the measure of the domain Ω.
Theorem 3.2
If
then DFE is locally stable, if
then it is unstable.
Proof
To study the stability of the equilibrium, let
Since the total population size is constant, we have
Note that eigenpairs of the scalar equations (Equation6) and (Equation7
) are also the solutions of the system (Equation5
)–(Equation7
), if we consider the solutions of the form (λ1, φ, ψ1, 0) or (λ2, φ, 0, ψ2). Clearly,
and
, whenever
, and
;
and
, whenever
, and
; and
and
, whenever
, and
. The perturbation system (Equation4
) is stable, iff
, and
, that is, iff
.
Theorem 3.3
If
then DFE is globally stable, if
then it is unstable.
The proof follows from the proof of Lemma 2.5 from Citation2.
Next, we consider the two semi-trivial equilibria. The I 1-equilibrium satisfies the following system:
To investigate local stability of semi-trivial equilibria, let S(x, t)=S*(x, t)+φ(x, t), , and I
2(x, t)=ψ2(x, t) and then linearizing (Equation1
) around
we obtain
Since (Equation12) decouples, we consider the following eigenvalue problem:
Let λ* denote the principal eigenvalue of (Equation13), which is given by the following variational formulation:
Theorem 3.4
Let λ*<0 and suppose that λ* is not an eigenvalue of (Equation15)–(
Equation16
), then the I
1
equilibrium
is unstable.
Proof
We use the principle of linearized stability theorem which states that the stability of the equilibrium is determined by the spectrum of the linearization at the equilibrium Citation24
Citation27. So, we consider the following eigenvalue problem:
Similar analysis is true for the I
2-equilibrium . For the I
2-equilibrium, we consider the following eigenvalue problem:
Theorem 3.5
Let
and suppose that
is not an eigenvalue of the following eigenvalue problem:
Proof
Similar to the proof of Theorem 3.4.
We denote the invasion numbers of strain one and strain two by , and
, respectively. The two invasion numbers are defined as follows:
Invasion number gives the ability of strain one to invade the I
2-equilibrium
measured as the number of secondary infections one strain one-infected individual can produce in a population where strain two is at equilibrium. Similarly, invasion number
gives the ability of strain two to invade the I
1-equilibrium
.
Lemma 3.1
The following statements are true for the invasion numbers
of strains one and two, respectively.
| i. |
| ||||
| ii. |
| ||||
| iii. |
| ||||
Proof
(i) The functions β1, β2, γ1, and γ2 are nonnegative on the domain Ω. Positivity of the invasion numbers ,
are satisfied by the existence of the positive single-strain equilibriums
and
. It is straightforward to show that the invasion numbers
,
are decreasing functions of d
1 and d
2, respectively.
(ii) Let φ(x) be the nonzero function in H
1(Ω) for which the is achieved in (Equation20
). Thus,
(iii) Proving this statement is similar to the analysis in (ii).
4. Coexistence equilibrium
Coexistence equilibrium, , satisfies the system (Equation2
) with Neuman boundary conditions (Equation3
). Since
and
it follows that for some positive constant κ
Lemma 4.1
We have
Proof is immediate and it is omitted. Lemma 4.1 implies that the operator G=(g 1, g 2) is quasi-monotone nondecreasing.
From the work of Nevai et al. Citation2 we know that there are unique single-strain solutions and
with
and
. We assume that Theorems 3.4 and 3.5 hold and therefore both of the semi-trivial equilibria are linearly unstable.
For vectors , we say that x≤
K
y if and only if x
1≤y
1 and x
2≥y
2. We say that
is a super-solution for (Equation26
)–(Equation28
) if
We have the following result Citation26 Citation27.
Lemma 4.2
Let
I
=(I
1
,
I
2) be a sub-solution and
be a super-solution for (Equation26
)–(
Equation28
). If
in Ω, then (Equation26
)–(
Equation28
) has a solution
with
.
Lemma 4.3
System (Equation26)–(Equation28
) has a positive sub-solution
I
and super-solution
.
Proof
We claim that for all sufficiently small ε>0, the following choices work:
Similar arguments show that is a sub-solution of (Equation26
)–(Equation28
) for all sufficiently small ε>0.
Lemma 4.4
Let (I*1
, 0) and
be linearly unstable single-strain solutions of (Equation26
)–(Equation28
). Then (Equation26
)–(Equation28
) has a positive solution
. Moreover, if we set
Proof
Let . Then,
5. Numerical method
In this section we approximate the solution of the model problem (Equation1). We derive the approximate solution of (Equation1
) in two steps. In the first step we approximate the solution by means of functions which for each fixed time belong to a finite dimensional space. These functions will be the solution of h-dependent finite system of ODEs in time and referred to as spatially discrete solutions. In the second step, we discretize this system in the time variable to obtain a fully discrete approximation of (Equation1
) by a finite difference method. For the notational convenience, we rewrite (Equation1
) in the vector form:
We adopt the standard notation for the Hilbert Spaces Citation1 Citation6 Citation20;
In general, we denote by H m (Ω) the Hilbert space of all L 2 functions with m derivatives (m≥0) in L 2(Ω). These Hilbert spaces are endowed with the norms
A variational formulation of (Equation39) is derived by first multiplying both sides of (Equation39
) by functions v in H
1(Ω). We, then obtain the weak formulation of (Equation39
) by Green’s formula. We look for a function u(x, t)∈H
1(Ω)×H
1(Ω)×H
1(Ω) (that is, for every t>0 the function
is in H
1(Ω)×H
1(Ω)×H
1(Ω)), such that
Find u
h
(x, t)∈χ
h
×χ
h
×χ
h
(that is, for every t>0 the function is in χ
h
), such that
We define χ
h
first by partitioning the domain Ω into triangles. Let denote the partition of Ω into triangles τ, such that no vertex of any triangle lies on the interior of a side of another triangle. Let h denote the mesh size, which is the maximal length of sides of the triangles in triangulation
. The parameter h is a measure of how refined the mesh is, the smaller the h, the finer the mesh. We assume that
is a quasi-uniform mesh in the sense that the triangles in
are essentially the same size. We also assume that all the angles in the triangulation
are acute, which means that all the angles are less than π/2. We denote the union of triangles in
by Ω
h
which is a convex polygonal domain. Clearly, Ω
h
is an approximation of bounded domain Ω with smooth boundary, and the error in approximating the domain Ω−Ω
h
does not change the rate of convergence Citation9. We thus assume Ω
h
=Ω.
We define χ
h
to be the space of continuous functions on Ω which are linear polynomials in each triangle of .
We now, discretize (Equation42) with respect to the time variable to obtain fully discrete approximation of (Equation1
). Let T denote the maximum time, k denote the time step, and J be a fixed nonnegative integer, then
We linearize the system by using discretization ,
for the approximation of nonlinear terms S
I
1/(S+I
1+I
2), S
I
2/ (S+I
1+I
2), respectively. At each time step, we solve three linear systems of algebraic equations simultaneously.
We use the vertex quadrature rule to approximate the L
2 inner product in χ
h
. Let be the vertices of the triangle τ, then the vertex quadrature rule, q
τ, h
(f), is defined as follows Citation30:
The system (Equation44) can be rewritten as:
5.1. Error analysis
We assume that the family of finite dimensional sub-spaces, χ h , of H 1(Ω) possesses the following approximation property Citation30: Let u∈H 2(Ω) and v∈χ h , then
Before considering the error estimates for the fully discrete approximation, we prove the following lemma.
Lemma 5.1
Let u=(S, I
1
, I
2) be the solution of (Equation39), and let u
h
=(u
s
, u
1
, u
2)>0 be the solution of (Equation41
), then
| i. |
| ||||
| ii. |
| ||||
| iii. |
| ||||
where
and
.
Proof
Here we demonstrate the proof of (ii), since the proofs of (i) and (iii) are similar. Adding and subtracting terms β1(S u 1/(u s +u 1+u 2)) and β1(S u 1/(S+I 1+I 2)) we obtain
We now perform error analysis for the fully discrete scheme.
Theorem 5.1
Let u=(S, I
1
, I
2
) and u
h
=(u
s
, u
1
, u
2
) be the solution of (
Equation39
) and (
Equation44
), respectively, then
Proof
Setting , in which we use the fact that the elliptic projection commutes with the finite difference operator, we obtain
Applying a similar procedure for , in (Equation44
) we obtain
Next, we estimate the residual terms. Estimation of the first terms in the residuals, are standard, and it is described in [Citation30, Theorem 1.5].
Using Lemma 4.1 for the remaining terms in we obtain
Thus, applying discrete Gronwall’s inequality we finally obtain,
5.2. Positivity of solutions
In this section we study the positivity of the solution to the problem (Equation45). We want to know whether any positive initial condition will lead to a positive solution of the system (Equation45
). Let
denote the time-dependent coefficients of the approximate solution (u
s
(x, t), u
1(x, t), u
2(x, t)) determined by the basis functions
. Using, α
n
=α(t
n
), in matrix notation (Equation45
) becomes
The initial condition α0 is the components of given initial approximation of u
0h
. Note that α0>0, since by (Equation43) α
s, j
(t
0)=u
s
(η
j
, t
0)>0.
The above system of equations may be written as
Definition 5.1 ([Citation5])
A real n×n matrix A=a i, j is said to have a Z n sign pattern if a i, j ≤0 for all i≠j.
We are interested in special subclass of matrices with Z n sign pattern, called M-matrices.
Definition 5.2 [([Citation31, p. 91])]
Let A have a Z n sign pattern, then it is an M matrix, if A is nonsingular and A −1 ≥0.
Note that if a matrix A has Z n sign pattern, then it can be expresses as A=s I−B, where B≥0. The matrix A is an M-matrix, if s≥ρ(B), the spectral radius of B.
Note that in one dimension, the stiffness matrix a is an M-matrix. In two dimensions, if all the angles in the triangulation are not greater than π/2, which is called an acute type triangulation, then the matrix is a is an M-matrix Citation10
Citation32. If all the angles of the triangulation
are acute and a vertex quadrature rule is used in the computation of the mass matrix m, then there exists a discrete-maximum principle. Clearly, this is not the case for the standard Galerkin method Citation30.
Lemma 5.2
We have the following properties for the matrices M and A:
| i. | The matrix M is an M-matrix. | ||||
| ii. | The matrix A is an M-matrix. | ||||
Definition 5.3 [([Citation5,Citation14])]
The directed graph, G(A), of an n×n matrix A consists of n vertices, P 1 , P 2 , …, P n where an edge leads from P i to P j if and only if a ij ≠0. The directed graph is called strongly connected if there is a path from each vertex to every other vertex.
Theorem 5.2 [Citation5,Citation14]
A matrix A is irreducible if and only if G(A) is strongly connected.
Proof
See [Citation14, Theorem 3.6] or [Citation5, Theorem 2.7].
Lemma 5.3
We have the following properties for the matrix X:
| i. | X has Z n sign pattern. | ||||
| ii. | X is irreducible. | ||||
Proof
| i. | The matrix M is a diagonal matrix with positive diagonal entries. The matrices A and −F(α n−1) have Z n sign patterns. So, X has a Z n sign pattern. | ||||
| ii. | By Theorem 5.2, X is irreducible if and only if G(X) is strongly connected. Clearly, X i, i =M i, i +k A i, i −k F i, i ≠0, and X i, i−1=k A i, i−1≠0, also X i, i+1=k A i, i+1≠0, for i=1, …, 3N h . Note that the elements of each component matrix a given by a i, j =(∇ϕ j , ∇ϕ i )≠0 if and only if i and j are the neighbouring nodes. Hence, the matrix a is a symmetric matrix. Also, note that if X i, j ≠0, then X j, i ≠0 as well. So, G(X) is strongly connected. | ||||
There are many equivalent conditions for M-matrices, among them we mention the following.
Theorem 5.3
Let A be an n×n irreducible matrix with a Z n sign pattern, then the following statements are equivalent:
| (i) | There exist a vector x>0 such that Ax>0. | ||||
| (i) | A −1 >0. | ||||
| (i) | A is an M-matrix. | ||||
Proof
See [Citation14, Theorem 5.12].
Theorem 5.4
Let α 0 >0, then α n >0 for n=1, 2, …, J.
Proof
To show that X is an M-matrix, we apply Theorem 5.3. Take a vector with all components equal to 1. Then,
for k small enough, where
and
, and
Hence y
X>0, therefore X
T
y
T
>0. By Theorem 5.3, X
T
is an M-matrix, and (X
T
)−1>0. Hence, X
−1>0 and X is an M-matrix.
6. Numerical estimation of the reproduction numbers
6.1. Numerical computation of basic reproduction numbers
In this section we develop a numerical method that will allow us to estimate whether the reproduction numbers , and the invasion numbers
, are each larger or smaller than one. We first focus on estimation of this threshold property for the reproduction numbers, since similar analysis is true for the invasion numbers. Recall that the expression for the reproduction number for each strain is implicit. Even if all the parameters are known, the computation of an approximate value is not an easy task. On the other hand, we only need to estimate if
or
for i=1, 2 to decide what type of dynamical outcome we can expect for each combination of parameters.
We demonstrate the numerical computation of reproduction number for , since the computation of reproduction number
is analogous to
.
It is discussed in Citation2 that the reproduction number for strain I
1 is a positive and monotone decreasing function of d
1>0. The relationship between
and the principal eigenvalue of the following eigenvalue problem can be proved by following a similar argument as in Citation2
Let λ1 be the principal eigenvalues of (Equation51), then
when λ1<0,
when λ1=0, and
when λ1>0.
The weak formulation of (Equation51) is, find
, and ψ∈H
1(Ω), ψ≠0 such that
It is also well known that
With and v
h
=ϕ
i
for i=1, …, n, we obtain the following generalized matrix eigenvalue problem:
06.2. Numerical computation of invasion numbers
We demonstrate the numerical computation of invasion number of strain one, since similar computation is true for In Lemma 3.1, the relationship between
and the principal eigenvalue of the following problem is proved:
The discrete weak formulation of (Equation54) within the finite dimensional space χ
h
is as follows: Find ψ
h
∈χ
h
such that
7. Numerical experiments
7.1. Verifying finite element method convergence rate
In this first example, we verify the convergence rates proved in Theorem 5.1.
Example 1
For the numerical computations, we choose d
s
=0.5, d
1=1, and d
2=1.5, use a uniform mesh on the unit square Ω=[0, 1]×[0, 1], set the final time to T=250 and solve the system (Equation1) with the following choices of transmission and recovery rates for strains one and two; β1=5−5x, γ1=1, β2=5x, γ2=1. Thus as it will be shown in Example 6, these choices of transmission and recovery rates will give coexistence equilibrium. In the error computations in , exact solution is taken to be the approximate solution at refinement step j=8, that is the approximate solution is computed with n=131, 072 elements, and m=66, 049 nodes. The time step is fixed in all refinements; it is set to k=1. Error is measured with ℓ2(0, T;L
2(Ω)) norm. Thus,
Table 1. Observed convergence rates, j denotes the refinement step, h denotes mesh size, n denotes the number of elements, and m denotes the number of nodes.
Looking at , we see that for each of the columns 5, 6, and 7, the error drops at least 4 times, as the mesh size h drops 2 times, confirming our theoretical result that the convergence rate is of order h 2.
In , we verify the convergence rate for the time step k. In error computations in , the exact solution is taken to be the approximate solution at refinement step j=8. The approximate solution is computed for J=6400 time steps. Mesh size is kept fixed, and all computations are done with n=25 elements. In , we observe that the order of convergence for the time step k is approximately 1 as j increases.
Table 2. Observed convergence rates, j denotes the refinement step, k denotes the time step, n denotes the number of elements, m denotes the number of time steps,  is the error computed at the refinement step j, and gives the ratio of the error computed at two consecutive refinement steps.
is the error computed at the refinement step j, and gives the ratio of the error computed at two consecutive refinement steps.
7.2. Verifying numerically some asymptotic properties of system (Equation1)
The reproduction numbers and invasion numbers often (but not always) control the outcome from the competition of the strains. We summarize the expected outcomes based on the values of the reproduction numbers and invasion numbers in .
Table 3. List of conditions on reproduction numbers and invasion numbers and potential dynamical outcomes of the competition of the strains. Last column gives the example where the case is numerically illustrated. Case 6 could not be numerically illustrated, possibly because it does not occur in system (Equation1).
Before presenting the numerical examples of some asymptotic properties of the system (Equation1), we first prove the following Proposition, which partially established properties 2 and 3 of (Equation1
).
Proposition 7.1
The following statements are true:
| (i) |
If
| ||||
| (i) |
If
| ||||
Proof follows from results in Citation2.
Proving rigorously all properties in may not be possible but we check numerically whether they hold at least for arbitrarily chosen parameter values in the examples presented below. For all examples below we choose d s =0.5, d 1=1, d 2=1.5, the domain to be unit square Ω=[0, 1]×[0, 1], and the initial conditions to be as follows:
Figure 1. Initial values.
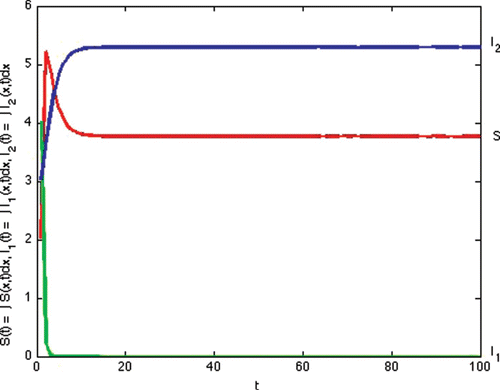
(a) Example 2. illustrates Case 2 in . In this example, we simulate the results of Proposition 7.1(i). Transmission and recovery rate of each strain is chosen as follows:
Figure 2. Total densities of susceptible individuals, infected individuals with strain one and two at time t.
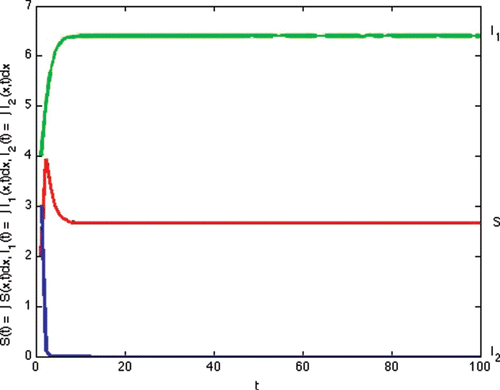
(b) Example 3. Example 3 illustrates Case 3 in . In this example, we also illustrate Proposition 7.1(ii). Transmission and recovery rate of each strain is chosen as follows:
Figure 3. Total densities of susceptible individuals, infected individuals with strain one and two at time t.
(c) Example 4. If both reproduction numbers are larger than one, as in the remaining examples, none of the strains is automatically eliminated, because it is unfit to the environment. The outcome of the competition between the strains is decided by the invasion numbers. Example 4 illustrates Case 4 in .
For this example, we chose the transmission and recovery rate of each strain as follows:
With these choices of recovery and transmission rates, we estimate that ,
,
and
. As stated in Case 4, I
2 persists and
. We observe this result in .
Figure 4. Total densities of susceptible individuals, infected individuals with strain one and two at time t.
(d) Example 5. Example 5 illustrates Case 5 in , and it is the symmetrical case to Case 4. If ,
,
, and
, then I
1 persists and
as shown in .
Figure 5. Total densities of susceptible individuals, infected individuals with strain one and two at time t.
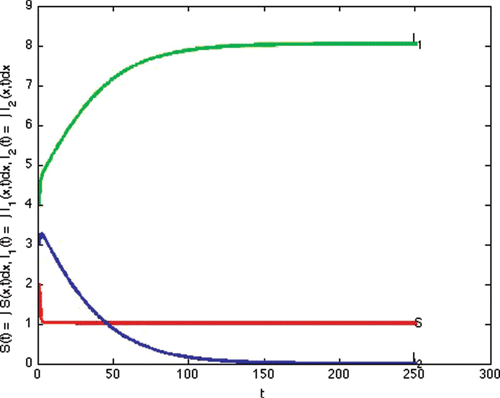
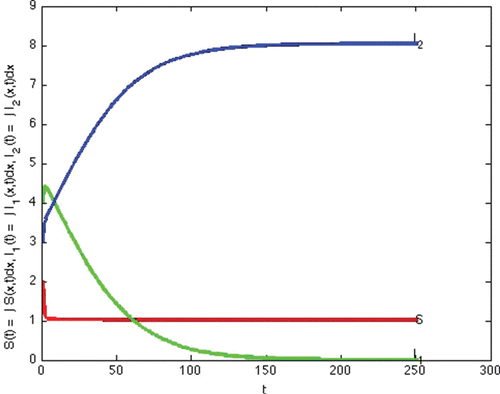
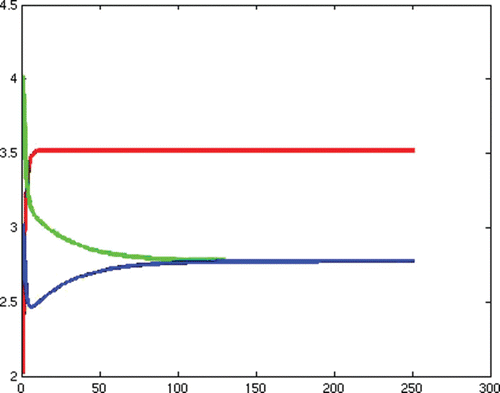
(e) Example 6. Example 6 illustrates Case 7 in . This is the case that both reproduction numbers are above one, and both invasion numbers are above one. The two strains coexist. We show theoretically that coexistence occurs in Lemma 4.3. In this section we demonstrate that the theoretical possibility of coexistence, actually occurs for some specific values of the parameters. We note that the transmission rate of strain one is quite different from the transmission rate of strain two, giving nontrivial different reproduction numbers.
In this simulation, with the following choices of recovery and transmission rates, we estimate that ,
,
, and
, and we observe that coexistence occurs (see )
Figure 6. Total densities of susceptible individuals, infected individuals with strain one and two at time t.
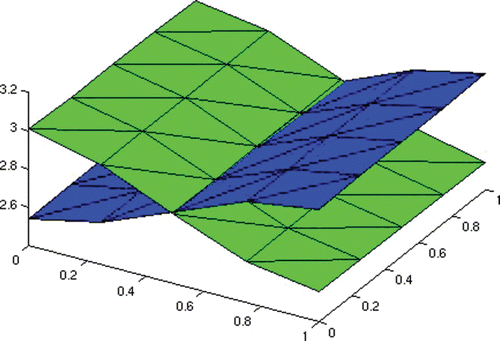
In , we plot the infected individuals with strain one and two at the final time of interest.
Figure 7. Coexistence equilibrium.
8. Discussion
In this article we introduce a spatially heterogeneous deterministic epidemic model of the SIS type where the pathogen is represented by two strains. We allow for spatially heterogeneous transmission and recovery rates, in contrast with most spatial epidemic models where rates are assumed constant. The model is built on the single-strain model considered in Citation2.
Fundamentally, our model is very simple – its spatially homogeneous ODE version has dynamics that leads to competitive exclusion. In other words, if any of the reproduction numbers is above one, the strain with the larger reproduction number persists, while the other one dies out (see Appendix). The introduction of space leads to larger variety of outcomes. Intuitively, it seems clear, that when space is taken into account, multiple pathogens could simultaneously persist by partitioning the region into subregions and each occupying separate subdomains. This scenario is known in ecology as ‘niche partitioning’. Here, we hypothesize that space alone may lead to coexistence of the pathogens, and we test this hypothesis both analytically and numerically. In particular, we define spatial reproduction numbers and invasion numbers for both pathogens. We find three types of equilibria: DFE, two dominance equilibria, one corresponding to strain one and the other one corresponding to strain two, and coexistence equilibria. We establish that if the reproduction numbers of both strains are below one, the DFE is globally asymptotically stable. Using the monotonicity of the time-independent problem, we show analytically, that if both invasion numbers are above one, coexistence equilibrium exists ().
To check the persistence of both strains in this case, we develop a fully discrete finite element method to simulate the system of partial differential equations in time. The spatial domain is divided into triangles with acute angles. We approximate the integrals with the vertex quadrature rule. Time is discretized with the backward Euler method. Although most finite element methods may not preserve the positivity of the solution, our discretization allows us to show rigorously that the discrete solution is nonnegative, just as the continuous solution it approximates. We also establish both analytically and numerically that the discrete solution converges to the exact solution with an error of order two in space and order one in time.
Finally, we simulate a number of dynamical outcomes of the competition of the strains controlled by different values of the reproduction numbers and invasion numbers. In particular, numerically we show that if one of the reproduction numbers is above one, and the other is below one, then the strain with the larger than one reproduction number persists, while the strain with the smaller than one reproduction number dies out. If both strains have reproduction number above one, then the outcome of the competition depends on the invasion numbers. If exactly one strain has an invasion number above one, that strain persist while the other is eliminated. If both strains have invasion numbers above one, then they both persist.
Acknowledgements
The authors thank two knowledgeable referees and the extremely helpful comments of the handling editor that helped improve the paper. Patrick De Leenheer contributed a great deal to the proof of the coexistence equilibrium. We also benefited from our discussions with Yuan Lou and Georg Hetzer. Maia Martcheva acknowledges NSF support from grant DMS-0817789.
Additional information
Notes on contributors
Necibe Tuncer
Current affiliation: Department of Mathematical and Computer Sciences, University of Tulsa, 800 South Tucker Drive, Tulsa, OK, 74104, USA.Related Research Data
References
- Adams , R. 1975 . Sobolev Spaces , New York : Academic Press .
- Allen , L. J.S. , Bolker , B. , Lou , Y. and Nevai , A. 2008 . Asymptotic profiles of the steady states for an SIS epidemic reaction–diffusion model . Discrete Contin. Dyn. Syst. , 21 : 1 – 20 .
- Babuska , I. and Osborn , J. E. 1987 . Estimates for the errors in eigenvalue and eigenvector approximation by Galerkin methods, with particular attention to the case of multiple eigenvalues . SIAM J. Numer. Anal. , 24 : 1249 – 1276 .
- Babuska , I. and Osborn , J. E. 1989 . Finite element Galerkin approximation of the eigenvalues and eigenvectors of selfadjoint problems . Math. Comput. , 52 : 275 – 297 .
- Berman , A. and Plemmons , R. J. 1979 . “ Nonnegative Matrices in the Mathematical Sciences ” . In Computer Science and Applied Mathematics , New York : Academic Press [Harcourt Brace Jovanovich Publishers] .
- Brenner , S. and Scott , L. R. 2007 . The Mathematical Theory of Finite Element Methods , New York : Springer-Verlag .
- Cantrell , R. and Cosner , C. 2003 . Spatial Ecology via Reaction–Diffusion Equations , Chichester : Wiley .
- Cantrell , R. S. and Cosner , C. 1998 . On the effects of spatial heterogeneity on the persistence of interacting species . J. Math. Biol. , 37 ( 2 ) : 103 – 145 .
- Ciarlet , P. 2002 . The Finite Element Method for Elliptic Problems , SIAM, Philadelphia .
- Ciarlet , P. G. and Raviart , P.-A. 1973 . Maximum principle and uniform convergence for the finite element method . Comput. Methods Appl. Mech. Eng. , 2 : 17 – 31 .
- Cosner , C. , Beier , J. , Cantrell , R. , Impoinvil , D. , Kapitanski , L. , Potts , M. , Troyo , A. and Ruan , S. 2009 . The effects of human movement on the persistence of vector-borne diseases . J. Theor. Biol. , 258 : 550 – 560 .
- Diekmann , O. and Heesterbeek , J. A.P. 2000 . Mathematical Epidemiology of Infectious Diseases , Chichester : Wiley .
- Evans , L. C. 1998 . “ Partial Differential Equations ” . In Graduate Studies in Mathematics , Vol. 19 , Providence , RI : American Mathematical Society .
- Fiedler , M. 1986 . Special Matrices and Their Applications in Numerical Mathematics , Dordrecht : Martinus Nijhoff Publishers . Translated from the Czech by Petr Přikryl and Karel Segeth
- Fitzgibbon , W. and Langlias , M. “ Simple models for the transmission of microparasites between host populations living on noncoincident spatial domains in Structured Population Models in Biology and Epidemiology ” . In Lecture Notes in Mathematics , Edited by: Magal , P. and Ruan , S. 115 – 164 . Berlin : Springer .
- Fitzgibbon , W. , Langlais , M. and Morgan , J. 2001 . A mathematical model of the spread of feline leukemia virus (FELV) through a highly heterogeneous spatial domain . SIAM J. Math. Anal. , 33 : 570 – 588 .
- Fitzgibbon , W. , Langlais , M. and Morgan , J. 2004 . A reaction--diffusion system modeling direct and indirect transmission of diseases . Discrete Contin. Dyn. Syst. B , 4 : 893 – 910 .
- Henry , D. 1981 . Geometric Theory of Semilinear Parabolic Equations , New York : Springer-Verlag .
- Hsieh , Y.-H. , van den Driessche , P. and Wang , L. 2007 . Impact of travel between patches for spatial spread of disease . Bull. Math. Biol. , 69 : 1355 – 1375 .
- Johnson , C. 2009 . Numerical Solution of Partial Differential Equations by the Finite Element Method , Mineola , NY : Dover Publications .
- Kim , M. 2006 . Global dynamics of approximate solutions to an age-structured epidemic model with diffusion . Adv. Comput. Math. , 25 : 451 – 474 .
- Kim , K. , Lin , Z. and Zhang , L. 2010 . Avian--human influenza epidemic model with diffusion . Nonlinear Anal.: Real World Appl. , 11 : 313 – 322 .
- Martcheva , M. , Bolker , B. and Holt , R. 2008 . Vaccine-induced pathogen strain replacement: What are the mechanisms? . J. Roy. Soc. Interface , 5 : 3 – 13 .
- Mora , X. 1983 . Semilinear parabolic problems define semiflows on C k spaces . Trans. Amer. Math. Soc. , 278 ( 1 ) : 21 – 55 .
- Pacala , S. W. and Roughgarden , J. 1982 . Spatial heterogeneity and interspecific competition . Theor. Popul. Biol. , 21 ( 1 ) : 92 – 113 .
- Pao , C. V. 1992 . Nonlinear Parabolic and Elliptic Equations , New York : Plenum Press .
- Smoller , J. 1983 . Shock Waves and Reaction--Diffusion Equations , New York : Springer-Verlag .
- Strang , G. and Fix , G. 1973 . An Analysis of the Finite Elment Method , Englewood Cliffs , NJ : Prentice-Hall .
- Strauss , W. A. 1992 . An introduction in Partial Differential Equations , New York : John Wiley & Sons .
- Thomèe , V. 1997 . Galerkin Finite Element Method for Parabolic Problems , Berlin : Springer .
- Varga , R. S. Matrix Iterative Analysis . expanded ed., Springer Series in Computational Mathematics Vol. 27, Springer-Verlag, Berlin, 2000.
- Vejchodskyacute; , T. 2004 . “ On the nonnegativity conservation in semidiscrete parabolic problems in Conjugate Gradient Algorithms and Finite Element Methods ” . In Scientific Computing , 197 – 210 . Berlin : Springer .
Appendix 1. The ODE model
The ODE version of the model (Equation1) is
Let β1−γ1>0 and β2−γ2>0, so that -equilibrium and
-equilibrium exist, then the stability matrices for these equilibria are
To study the global stability of the coexistence equilibrium, set , then