Abstract
Periodic environments may either enhance or suppress a population via resonant or attenuant cycles. We derive signature functions for predicting the responses of two competing populations to 2-periodic oscillations in six model parameters. Two of these parameters provide a non-trivial equilibrium and two provide the carrying capacities of each species in the absence of the other, but the remaining two are arbitrary and could be intrinsic growth rates. Each signature function is the sign of a weighted sum of the relative strengths of the oscillations of the perturbed parameters. Periodic environments are favourable for populations when the signature function is positive and are deleterious if the signature function is negative. We compute the signature functions of four classical, discrete-time two-species populations and determine regions in parameter space which are either favourable or detrimental to the populations. The six-parameter models include the Logistic, Ricker, Beverton–Holt, and Hassell models.
AMS Subject Classifications:
1. Introduction
Populations live naturally in changing environments in which variations caused by food availability, migrations, tidal patterns, and mating habits may be assumed to be cyclic or periodic. As a result, fluctuating environments are of particular interest to population biologists Citation2 Citation3 Citation6 Citation10 Citation15 Citation16 Citation22. For example, in the 1980 controlled laboratory experiments of flour beetles (Tribolium), Jillson observed that a periodic food supply generated cyclic oscillations within the beetle population. The alternating habitat in fact supported a larger beetle population, more than twice that of a constant environment even though the average flour volume remained the same Citation17 Citation18. It has since been demonstrated that under certain conditions, periodic environments may be advantageous to a population and yet deleterious under others Citation5 Citation9 Citation11. In other words, the average of the resulting population oscillations in a periodic environment may either be greater or be less than, respectively, the average of the carrying capacities in an otherwise constant environment.
In constant environments, many discrete-time two-species models are extensions of single-species models that contain at least two parameters, usually the carrying capacity and a demographic characteristic Citation3–5. Examples of such single-species models include the Logistic, Ricker, Beverton–Holt, and Hassell models. In recent papers, Franke and Yakubu Citation11 Citation12 have investigated the responses of classical, single-species, discrete-time models to periodic oscillations in two and three parameters. In this paper, we extend their investigations to study the responses of a two-dimensional system to periodic fluctuations in six model parameters and compare them against those elicited by a constant environment. More specifically, we analyse the effects that a 2-periodic forcing of six parameters has on a discretely reproducing two-species population model of a competitive type and investigate a 2-cycle population that bifurcates from the perturbed interior fixed point, which we refer to as the carrying capacity fixed point. We take a special interest in the case where the two species are very similar as might happen when a mutation occurs in a species.
Section 2 introduces a general framework for studying the impact of environmental fluctuations on a discrete-time two-species population model with six oscillating parameters. Here, we provide conditions sufficient for our unforced model to have a stable fixed point. In Sections 3 and 4, we impose 2-periodic forcing on the model and prove that small 2-periodic perturbations support 2-cycle populations. We study the 2-cycle that must, for small forcing, be close to the carrying capacity fixed point defined in Section 2.
In Section 5, we derive a signature function, ℛ d , for determining whether the average total biomass is suppressed via attenuant stable 2-cycles or augmented via resonant stable 2-cycles. As in Citation11 Citation12, ℛ d is the sign of a weighted sum of the relative strengths of the oscillations of the carrying capacities and the remaining parameters. When these oscillations are small and the environment is 2-periodic, we prove that the average total biomass of the two-species system diminishes if ℛ d is negative and is enhanced when ℛ d is positive. Also in Section 5, we derive similar signature functions, 𝒮 d and 𝒯 d , for each individual species and determine the response that species makes to the periodic environment. We then obtain conditions on the relative strengths of the oscillations sufficient for obtaining positive or negative signature functions.
Furthermore, we establish that the relative strengths of the oscillations of this carrying capacity fixed point and the remaining four model parameters are critical factors in predicting how a two-species population responds to a periodic environment. We observe that changes in the relative strengths of the parameter fluctuations are capable of shifting population dynamics from resonant to attenuant 2-cycles or vice versa. It is known that this dramatic shift is not possible in single-species models with a single oscillating parameter Citation11 Citation14 Citation15. We illustrate these concepts in Section 6 by computing the signature functions of four discrete-time, competitive, two-species models and provide parameter regimes for the occurrence of stable attenuant and resonant 2-cycles that bifurcate from the carrying capacity fixed point. Finally, the implications of our results are discussed in Section 7, and the derivations of the signature functions are included in Appendix 1.
2. Two-species population models of six parameters
Most single-species ecological models contain two or more model parameters and so, by extension, many two-species ecological models contain at least four parameters Citation5 Citation23. To study the combined effects of 2-periodic forcing on models with six parameters, we consider two-species population models of the general form
To simplify the notation, we let for a pre-selected set of positive parameters
. We can then define the function
We frequently refer to the total biomass and the carrying capacity fixed point throughout our discussion. These are defined below.
Definition 2.1
The total biomass of the system defined by Model
Equation(1)
refers to the sum of the x and y populations at the same generation: x(t)+y(t).
The interior fixed point, (L, M), of Model Equation(1) is often globally attracting in our examples Citation23. This fixed point thus captures the idea of carrying capacity and we make the following definition.
Definition 2.2
The carrying capacity fixed point is the fixed point population, (L, M), of Model
Equation(1).
For each set of six positive constants , we define
One way to generate models of the type we are studying is to start with a one-species model:
Table 1. Examples of multiparameter population models.
The following result provides necessary and sufficient stability conditions whereby the carrying capacity fixed point is locally attracting for Model Equation(1).
Theorem 2.3
The carrying capacity fixed point is a hyperbolic attracting fixed point of Model
Equation(1)
if and only if
Proof
The derivative matrix of F(P(x, y)) evaluated at the carrying capacity fixed point, , is given by
If the carrying capacity fixed point is both hyperbolic and locally attracting, then the eigenvalues of EquationEquation (4) lie entirely inside the unit circle, in which case
For the reverse direction, first use to obtain
A direct application of the inequalities in Theorem 2.3 to the four models in yields the necessary and sufficient conditions for (L, M) to be a hyperbolic attracting fixed point presented in .
Table 2. Stability conditions for the six-parameter two-species models given in .
3. 2-Periodic forcing
We introduce 2-periodic forcing into our model by having each parameter oscillate about its original value by fractional amounts. Let a, b, c, d, e, and f be the maximal fractional oscillations of L, M, S, T, B, and C, respectively. Model Equation(1) becomes
To study the changing behaviour of our system as the fractional oscillations of B, C, S, T, L, and M vary from zero to their maximal amounts, we introduce three bifurcation parameters: . We are particularly interested in the case where the two species differ only slightly, as might happen after a mutation has occurred within one species. It is thus reasonable to assume that each term in the pairs (L, M), (S, T), and (B, C) behaves similarly to fluctuations in the environment, and the parameters
, and γ are taken to be the fraction of the maximal oscillations of these pairs, respectively. This extends EquationEquation (9)
to our general, periodically forced model for this paper, Model Equation(10)
,
In Section 6, we study the four 2-periodically forced versions of the models listed in to illustrate specific examples of Model Equation(10).
When
Similar to Franke and Yakubu Citation7 Citation8 Citation11 Citation12, if the carrying capacity fixed point, (L, M), is the source of a 2-cycle, we use the following definition to compare the average of the 2-cycle with the unforced total biomass.
Definition 3.1
A 2-cycle,
of Model
Equation(10)
is attenuant (resonant) if its average total biomass,
A two-species population cycle of Model Equation(10) is thus attenuant (resonant) if its average total biomass decreases (increases) in the advent of periodic forcing. If
is a 2-cycle for Model Equation(10)
, then
and
are each 2-cycles for the individual x-species and y-species. Analogous definitions of attenuance and resonance for a single-species 2-cycle of Model Equation(10)
are provided below.
Definition 3.2
A 2-cycle,
or
of a single species of Model
Equation(10)
is attenuant (resonant) if its average value is less (greater) than L or M, respectively, the corresponding component of the carrying capacity of the system
Citation5
Citation11.
Attenuant and resonant cycles thus refer to a decrease and an increase, respectively, in average total population sizes.
4. 2-cycle population oscillations from unforced carrying capacity
Henson demonstrated that small perturbations of the food supply of a single species can generate 2-periodic population cycles of period two Citation14. Later, Franke and Yakubu Citation11 showed that small, 2-periodic perturbations of the carrying capacity and the demographic characteristic of discrete-time, single-species population models produce 2-cycle populations. In this section, we demonstrate that small, 2-periodic fluctuations of the six parameters in Model Equation(1) can produce 2-cycle populations,
, with x
0 and x
1 near L and y
0 and y
1 near M. That is,
is near L+M. In a constant environment, this 2-cycle population reduces to the carrying capacity fixed point.
We simplify the notation again and let
The following theorem provides conditions sufficient for Model Equation(10) to produce a 2-cycle that is perturbed from the carrying capacity fixed point.
Theorem 4.1
Suppose
Proof
Let
Note that . We must establish that when
, the eigenvalues of
are non-zero where I
2 is the 2×2 identity matrix. Since Model Equation(10)
reduces to the unperturbed system of Model Equation(1)
,
If is non-zero, then
Since the carrying capacity fixed point is a hyperbolic fixed point of F(P(x, y)) when the eigenvalues of its derivative matrix lie off the unit circle, the following consequence of Theorem 4.1 is immediate.
Corollary 4.2
If the carrying capacity fixed point of Model
Equation(1)
is hyperbolic, then for all sufficiently small
and
Model
Equation(10)
has a 2-cycle population,
By Corollary 4.2, also describes parameter regions for the occurrence of an asymptotically stable 2-cycle in the four population models given in under small period-two perturbations of the parameters near the carrying capacity fixed point. In Section 5, we derive an equation for the average total biomass of these 2-cycles.
5. Resonance versus attenuance: 2-cycle bifurcation from unforced carrying capacity
In single-species population models, small perturbations of a single parameter usually generate either attenuant or resonant cycles but not both, whereas small perturbations in single-species population models of two parameters can generate both attenuant and resonant 2-cycles that depend on the relative strengths of the fluctuations Citation11
Citation14. We demonstrate that small perturbations of the six parameters in Model Equation(1) generate both attenuant and resonant 2-cycles depending on the relative strengths of the fluctuations. As in the previous sections, we assume that the 2-cycle must, for small forcing, be close to the carrying capacity fixed point and that the model is competitive. For this 2-cycle, we derive a signature function, ℛ
d
, for determining whether the average total biomass is enhanced via resonance or suppressed via attenuance. We also define signature functions, 𝒮
d
and 𝒯
d
, to determine how each individual species responds to the alternating environment.
When the carrying capacity fixed point, (L, M), of EquationEquation (2) is hyperbolic, Corollary 4.2 guarantees that the 2-cycle solution of Model Equation(10)
may be expanded in terms of
, and γ as follows:
The expansion of the second point in the 2-cycle with respect to and γ is similar:
The next two lemmas eliminate several of the coefficients in EquationEquations (11) and Equation(12)
and establish the following expression for the average total biomass of the 2-cycle:
Similarly, the average biomasses of the individual populations of the x-species and y-species, respectively, are given by
Lemma 5.1
In Equations
Equation(11)
and
Equation(12)
,
Proof
When α=0,
However, since α is the fraction of the maximal oscillation of the carrying capacities, the fixed points of and
are both (L, M). Thus, for
we have
, and hence
By this result, the coefficients of the relative strengths of and γ2 in EquationEquations (11)
and Equation(12)
are zero. The next result establishes that the sum of the coefficients of the relative strengths of α in EquationEquations (11)
and Equation(12)
is also zero.
Lemma 5.2
If
, then
Proof
We have
Since , we have
Let
In the following result, we show that ℛ
d
indicates when the 2-cycle is either attenuant or resonant. Analogous results follow for 𝒮
d
and which describe the individual responses of the x-species and y-species, respectively, to a periodic environment.
Theorem 5.3
If the carrying capacity fixed point of Model
Equation(1)
is hyperbolic, then for sufficiently small
and
Model
Equation(10)
has an attenuant (a resonant) 2-cycle if ℛ
d
is negative (positive).
Proof
Lemmas 5.1 and 5.2 establish that the average of the 2-cycle predicted in Corollary 4.2 satisfies
Since
Theorem 5.4
If the carrying capacity fixed point of Model
Equation(1)
is hyperbolic, then for all sufficiently small
and
the x
-species in Model
Equation(10)
has an attenuant (a resonant) 2-cycle if 𝒮
d
is negative (positive).
Theorem 5.5
If the carrying capacity fixed point Model
Equation(1)
is hyperbolic, then for all sufficiently small
and
the y
-species in Model
Equation(10)
has an attenuant (a resonant) 2-cycle if 𝒯
d
is negative (positive).
As demonstrated in Lemma 5.1, when both the individual carrying capacities, S and T, and the two arbitrary model parameters, B and C, are fluctuating but the parameters L and M are constant (i.e. α=0, β≠0, and γ≠0), the 2-cycle degenerates into a fixed point at the carrying capacity fixed point. However, if the carrying capacity fixed point, (L, M), is fluctuating while the remaining parameters are constant (α≠0, β=0, and γ=0), Theorem 5.3 gives the following result.
Corollary 5.6
If the carrying capacity fixed point of Model
Equation(1)
is hyperbolic and the individual carrying capacities, S and T, as well as the two arbitrary model parameters, B and C, are all constant
then for all sufficiently small
(i)
| |||||
(ii)
| |||||
(iii)
| |||||
Population models with six parameters which are 2-periodically forced are capable of experiencing both resonance and attenuance. We formalize this in the following result.
Corollary 5.7
If the carrying capacity fixed point of Model
Equation(1)
is hyperbolic, then for all sufficiently small
and
Model
Equation(10)
has an attenuant 2-cycle and a resonant 2-cycle for different choices of
and γ, provided that either u
2
or u
3
is non-zero.
Proof
If u
2>0, take α>0, and γ=0. Then,
and
is negative. The 2-cycle is therefore attenuant. For a resonant 2-cycle, take
and γ=0. If u
2<0, take
(
), and γ=0 to obtain an attenuant (resonant) 2-cycle. Similar arguments establish the rest of the proof. ▪
Analogous results hold for Model Equation(10) when considering the x-species or y-species separately. In this case, either substitute w
1, w
2, and w
3 or v
1, v
2, and v
3 for u
1, u
2, and u
3, respectively, in Corollary 5.7 to see that if either w
2 or w
3 is non-zero or either v
2 or v
3 is non-zero, then Model Equation(10)
is capable of generating both attenuant and resonant 2-cycles in the x-species or y-species, respectively.
6. Signature functions for classical models
In this section, we use our theorems to study the impact of the combined effects of fluctuations in the carrying capacities and in the arbitrary model parameters on the average total biomass of each population governed by the models given in . Appendix 1 provides more specific details for obtaining general formulas for ℛ d , 𝒮 d , and 𝒯 d in terms of g 1 and g 2 and the partial derivatives of g 1 and g 2, all evaluated at P(L, M). Unfortunately, each of these formulas is much too long to be included, and for these reasons, we instead present some interesting results for the special case where B=C, S=T, and L=M.
Under these conditions, the growth rate functions of EquationEquation (3) reduce to
We observe what effects fluctuations in the parameters have on the total average biomass of the system as well as on each species. The signature functions are computed in order to investigate regions in the parameter space of attenuance and resonance of the 2-cycle which is perturbed from the carrying capacity fixed point.
In each model, the six constants a, b, c, d, e, and f are the maximal fractional oscillations of the six parameters, while the parameters S, T, L, and M are also consistent with the general periodically forced model Equation(10). The arbitrary parameters B and C are regarded in the Logistic, Ricker, and Beverton–Holt (with B, C>1) models as inherent growth rates, but in the Hassell model, they are more complicated and are taken instead to be demographic parameters.
Recall that each model given in is assumed to be competitive at the carrying capacity fixed point. This is realized if and only if and
are both negative when evaluated at P(L, L), the carrying capacity in this special case. Finally, we continue with the notation of Model Equation(10)
by using
6.1. Model I, Logistic
Both the carrying capacities and the inherent growth rates are 2-periodically forced in Model Equation(10). Using the classic Logistic model, we obtain
According to , the carrying capacity fixed point in a constant environment is asymptotically stable as long as
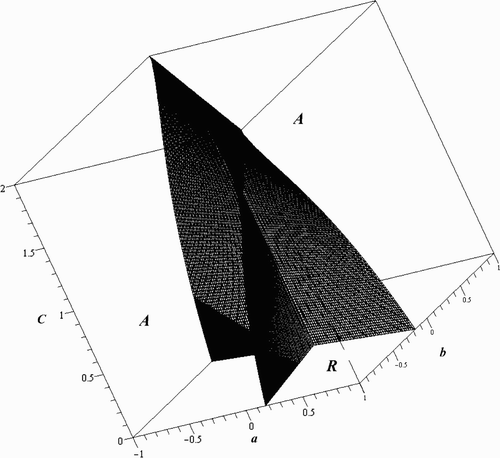
Since we are looking at the scenario where B=C, S=T, and L=M, we begin by considering the case where the maximal oscillations in these pairs of parameters are also equal; that is, when a=b, c=d, and e=f. The calculations discussed in Appendix 1 give
Since , the resonant and attenuant properties of the two individual species are exactly the same. On the stable region, C<2. Thus, if there is no fluctuation in the inherent growth rates (γ=0), then all three of the signature functions are negative on the stable region and we get attenuance of each species as well as of the total biomass. In contrast, we observe a resonant situation in the Logistic model when the fluctuations in the inherent growth rates are strong enough, namely when α>0 and
Fluctuations in the components of the carrying capacity for the system (L, M), which is (L, L) in our special case, are of paramount importance and should be investigated if they are fluctuating differently. To study this case, we let c=d and e=f, but allow a and b to vary. This effectively synchronizes the fluctuations of the intrinsic growth rates and of the individual carrying capacities, but not the fluctuations of the carrying capacities of the system. Here,
When a≠b, the relationship between w
1 and v
1 is more complicated. They become 2-forms in a and b and are equal when , which lies inside the stable region. Their difference,
The quantities w
1 and v
1 are functions of a, b, C, T, and L, but if we restrict to a given hyperplane, L=kT, the surface where is only a function of a, b, and C. Taking γ=0 and L=3T/4, the equation of the surface where u
1=0 is
Figure 1. Regions of attenuance and resonance in the a, b, and C plane for the 2-periodic Logistic model when γ=0 and L=3T/4.
6.2. Model II, Ricker
When both the carrying capacities and the inherent growth rates are 2-periodically forced as in Model Equation(10), the classic Ricker model becomes
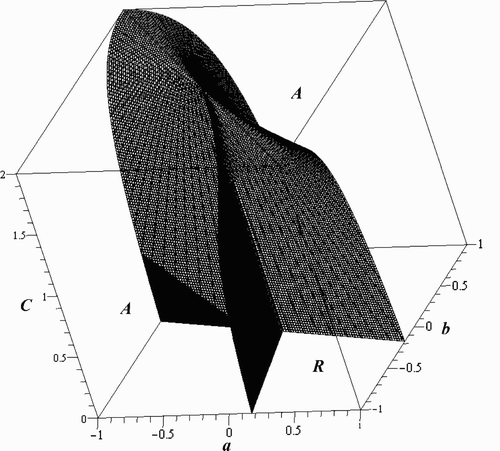
Once more , and the resonant and attenuant dynamics of the two individual species are exactly the same. As with the Logistic model, if there are no oscillations in the inherent growth rates (γ=0), then all three of the signature functions are negative on the stable region (C<2), and we witness attenuance of each species as well as of the total biomass. Resonance is still possible in Model Equation(16)
, provided the oscillations in the inherent growth rates are strong enough. According to Corollary 5.7, this occurs when α>0 and
Oscillations in the components of the carrying capacity for this system, which is (L, L) in this scenario, are of particular importance and, if they are fluctuating differently, should be investigated. In this case, we let c=d and e=f but assume that a and b can be different. The formulas for w 2 and v 2 turn out the same as those in the Logistic model, where
The relationship between w 1 and v 1 is no longer straightforward if a≠b. They are 2-forms in a and b and are equal when C=T/L, which belongs to the stable region. Their difference,
As in the Logistic model, w 1 and v 1 are the functions of a, b, C, T, and L. Restricting to a given hyperplane, L=kT, again reduces the surface where u 1=0 to a more manageable function of three parameters, a, b, and C. To stay in the stable region, we must restrict 1/2<k<1, but the surface retains its same basic shape for this range of k. Taking γ=0, there are four distinct regions, two giving attenuance and two giving resonance. Taking γ=0 and L=3T/4, the equation of the surface where u 1=0 is
Figure 2. Regions of attenuance and resonance in the a, b, and C plane for the 2-periodic Ricker model for γ=0 and L=3T/4.
6.3. Model III, Beverton–Holt
If both the carrying capacities and the inherent growth rates are 2-periodically forced, as in Model Equation(10), the classic Beverton–Holt model becomes
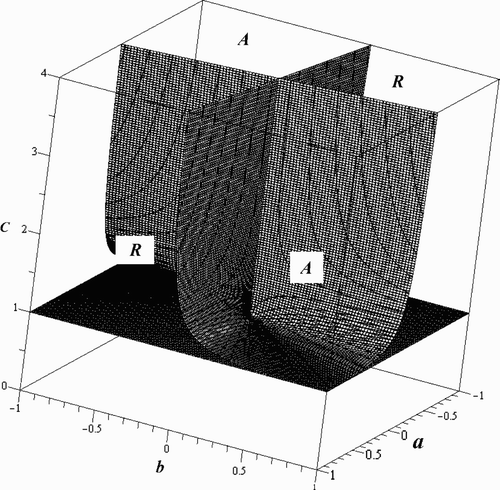
We are investigating the case where B=C, S=T, and L=M, and as before, we start by looking at the situation when the maximal oscillations in these pairs of parameters are also equal (when a=b, c=d, and e=f). The calculations in Appendix 1 give
Again, , and the resonant and attenuant properties of the two individual species are exactly the same. If there are no fluctuations in the inherent growth rates (γ=0), then all three of the signature functions are negative within the stable region (C>1), and we thus observe attenuance in each species as well as in the total biomass. Attenuance is also possible when γ is non-zero, as long as α and γ have the same sign. However, to observe a resonant response, α and γ must have opposite signs with
Next, we consider the situation where the components of the carrying capacity for the system, (L, L) in this special case, might be oscillating differently, but the fluctuations of the remaining parameters are synchronized. We fix c=d and e=f, but allow a and b to differ. The simplest result in this case is
As with Models Equation(15) and Equation(16)
, the relationship in the Beverton–Holt model between w
1 and v
1 is more complicated when a≠b as they are again 2-forms in both a and b. They are equal when
, but, unlike the previous two models, this intersection does not lie inside the stable region. Their difference,
The quantities w 1 and v 1 are still functions of a, b, C, T, and L. By restricting to a given hyperplane, L=kT, the surface where u 1=0 reduces to one of three variables, a, b, and C. We must restrict 1/2<k<1 in order to remain inside the stable region, but we find that the surface maintains a similar shape for these values of k. If γ=0, there are four distinct regions, two giving attenuance and two giving resonance. Further taking γ=0 and L=3T/4, the equation of the surface where u 1=0 is
Figure 3. Regions of attenuance and resonance in the a, b, and C plane for the 2-periodic Beverton–Holt model with γ=0 and L=3T/4.
6.4. Model IV, Hassell
In the Hassell model, the B and C parameters are exponents. As a consequence, their roles are more complicated than in the previous three models but can be thought of as demographic parameters. When the carrying capacities and the demographic parameters are 2-periodically forced as in Model Equation(10), the classic Hassell model becomes
must have the same sign, but are zero when C=(2T+2)/T and Since
, we have
The signs of w
1, v
1, and u
1 change from negative to positive as C goes from below to above . Since
, this sign change occurs inside the stable region. With β=0 and γ=0, we get attenuance in the stable region when C<(T+2)/T and resonance when C>(T+2)/T. This is in marked contrast to what we have seen in the first three models where only attenuance is observed. Since all of the remaining w
2, v
2, u
2, w
3, v
3, and u
3 are positive everywhere on the stable region, we get resonance in the stable region provided that
, and γ are all positive and C>(T+2)/T.
Fluctuations in the components of the carrying capacity (L, M), which is (L, L) in our special case, are of particular importance and should be investigated if they are oscillating differently. We thus fix c=d and e=f, but assume that a and b can vary. This Hassell model gives results that are quite different from those obtained for our first three models. In particular, u 2≠0 since w 2≠−v 2. Instead,
The relationship between w 1 and v 1 is also more complicated than those of the other three models. This time both are different 2-forms in a and b and are equal on the surface:
Where u 1 changes sign is a function of a, b, C, T, and L. Unfortunately, when we fix a hyperplane, L=kT, the surface where u 1=0 is now a function of a, b, C, and T. This would require a four-dimensional graph to illustrate and so we cannot provide one. The equation for the surface is a 2-form in a and b, but cubic in both C and T. One can analyse u 1 to observe parameter regimes in its stable region inducing both attenuance and resonance.
7. Conclusions
Many experimental and theoretical studies predict that single-species populations may either be enhanced or suppressed by periodic environments Citation5 Citation9 Citation11 Citation16 Citation18. We derived a discrete, two-species, competitive, six-parameter model to investigate the effects of 2-periodic forcing of the parameters. Our investigation supports these predictions by establishing parameter regions where the total population is enhanced by the periodic forcing and other regions where it is suppressed.
Two of the parameters in the model give an interior fixed point which is globally attracting in many example models and acts as the system carrying capacity. The next two parameters are the carrying capacity of the individual species in the absence of the other. The remaining two parameters can be quite general, but in our specific examples, they are taken either to be intrinsic growth rates or to be more general demographic parameters.
The 2-periodic forcing is first realized by assuming that each of the six parameters has a maximal fractional oscillation. Next, we introduce three bifurcation parameters, in order to investigate the situation where the oscillations in the six model parameters range from zero to plus or minus the maximal oscillation. These three relative strengths of the oscillation parameters organize the six model parameters into three groups: the system carrying capacity, the two individual carrying capacities, and the two arbitrary parameters.
We proved that small 2-periodic fluctuations are capable of supporting 2-cyclic oscillations of the total biomass of the system and also of each species. We derived signature functions, ℛ
d
, and
for predicting the responses of the total biomass and of each of the two species to 2-periodic fluctuations in the six parameters. Each signature function is the sign of a weighted sum of the three relative strengths of the bifurcation parameters of the oscillations. Periodic environments are deleterious for the total biomass and for each species when the corresponding signature function is negative, but are favourable if the corresponding signature function is positive.
We applied these ideas to four models derived from the classic Logistic, Ricker, Beverton–Holt, and Hassell models and observed both attenuance and resonance in each model. We also observed that changes in the relative strengths of these fluctuations can shift a system from attenuance to resonance or vice versa. The resonance regions in the Logistic and Ricker models are quite similar and considerably smaller than their attenuance regions, as opposed to the Beverton–Holt model where the relative sizes of the regions are interchanged. The resonance and attenuance regions for the Hassell model are more complicated to picture, but in some sense, resonance is a more likely outcome than in the other three models. Special attention is paid to the situation where the terms in each pair of the three groups of model parameters are equal and have the same maximal oscillation. In this case, the two competing species share many common features as could result from one species undergoing a mutation to produce a second competitive species with very similar characteristics.
Our results indicate that the response of a population to a periodic environment is a complex function of the period of the environment, the carrying capacities of the two species, the demographic characteristic, and the type and nature of the fluctuations. These relationships, which affect the dynamics of populations living in periodic environments, are implicitly represented in the signature functions. Further investigations on these relationships and their biological implications are welcome.
Acknowledgements
We are grateful to the referees for the careful reading of this manuscript.
References
- Beverton , R. and Holt , S. 1957 . On the dynamics of exploited fish populations . Fishery Investig. , 19 : 533
- Costantino , R. , Cushing , J. , Dennis , B. , Desharnais , R. and Henson , S. 1998 . Resonant population cycles in temporally fluctuating habitats . Bull. Math. Biol. , 60 : 247 – 273 .
- Cushing , J. 1980 . Two species competition in a periodic environment . J. Math. Biol. , 10 : 385 – 400 .
- Cushing , J. 1984 . Periodic two-predator, one-prey interactions and the time sharing of a resource niche . SIAM J. Appl. Math. , 44 : 392 – 410 .
- Cushing , J. and Henson , S. 2001 . Global dynamics of some periodically forced, monotone difference equations . J. Difference Equ. Appl. , 7 : 859 – 872 .
- Cushing , J. , Levarge , S. , Chitnis , N. and Henson , S. 2004 . Some discrete competition models and the competitive exclusion principle . J. Difference Equ. Appl. , 10 : 1139 – 1151 .
- Franke , J. and Yakubu , A. 2005 . Periodic dynamical systems in unidirectional metapopulation models . J. Difference Equ. Appl. , 11 : 687 – 700 .
- Franke , J. and Yakubu , A. 2005 . Population models with periodic recruitment functions and survival rates . J. Difference Equ. Appl. , 11 : 1169 – 1184 .
- Franke , J. and Yakubu , A. 2006 . Signature function for predicting resonant and attenuant population 2-cycles . Bull. Math. Biol. , 68 : 2069 – 2104 .
- Franke , J. and Yakubu , A. 2006 . Globally attracting attenuant versus resonant cycles in periodic compensatory Leslie models . Math. Biosci. , 204 : 1 – 20 .
- Franke , J. and Yakubu , A. 2006 . Attenuant cycles in periodically forced discrete-time age-structured population models . J. Math. Anal. Appl. , 316 : 69 – 86 .
- Franke , J. and Yakubu , A. 2007 . Using a signature function to determine resonant and attenuant 2-cycles in the Smith-Slatkin population model . J. Difference Equ. Appl. , 13 : 289 – 308 .
- Hassell , M. 1975 . Density-dependence in single-species populations . J. Anim. Ecol. , 44 : 283 – 295 .
- Henson , S. 1999 . The effect of periodicity in maps . J. Difference Equ. Appl. , 5 : 31 – 56 .
- Henson , S. 2000 . Multiple attractors and resonance in periodically forced population models . Phys. D , 140 : 33 – 49 .
- Henson , S. and Cushing , J. 1997 . The effect of periodic habitat fluctuations on a nonlinear insect population model . J. Math. Biol. , 36 : 201 – 226 .
- Henson , S. , Costantino , R. , Cushing , J. , Dennis , B. and Desharnais , R. 1999 . Multiple attractors, saddles, and population dynamics in periodic habitats . Bull. Math. Biol. , 61 : 1121 – 1149 .
- Jillson , D. 1980 . Insect populations respond to fluctuating environments . Nature , 288 : 699 – 700 .
- Marsden , J. and Hoffman , M. 1993 . Elementary Classical Analysis , 2 Edited by: Freeman , W. H. New York
- May , R. 1976 . Simple mathematical models with very complicated dynamics . Nature , 261 : 459 – 467 .
- Ricker , W. 1954 . Stock and recruitment . J. Fisheries Board Canada , 11 : 559 – 623 .
- Rosenblat , S. 1980 . Population models in a periodically fluctuating environment . J. Math. Biol. , 9 : 23 – 36 .
- Wendi , W. and Zhengyi , L. 1999 . Global stability of discrete models of Lotka–Volterra type . Nonlinear Anal. , 35 : 1019 – 1030 .
Appendix 1
Here, we outline the derivation of the formulas for w
1, w
2, w
3, v
1, v
2, and v
3 which determine u
1, u
2, and u
3. From these coefficients, one may construct the signature functions derived in Section 5 in terms of ,
, g
1, g
2, and the partial derivatives of g
1 and g
2. This appendix ends with proofs for Theorems 5.4 and 5.5.
We apply the implicit function theorem to
Since
Proof
[Proof of Theorems 5.4 and 5.5] Lemmas 5.1 and 5.2 establish that the average of the 2-cycle predicted in Corollary 4.2 satisfies the equation
Since