
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
We investigate the properties of five mathematical models used to represent the growth of a single population. By imposing a common set of (normalizing) initial conditions, we are able to calculate and explicitly compare the time intervals required to reach specific values of population levels. Based on these results, we conclude that one must be careful when applying these models to interpret the dynamics of single-population growth. An additional implication is that they provide evidence that such caution should also be extended to the incorporation of these models into the formulation of interacting, multi-population models, which are used, for example, to study the spread of disease.
1. Introduction
The purpose of this paper is to discuss five models of single-population growth, with a particular emphasis on the length of time each takes to reach specific population levels. A reason for carrying out this study is to see what differences exist with regard to this feature for the models. However, a more important issue is what exactly happens when these different models are, respectively, used in more complex interacting population systems. If the individual single-population models greatly differ in the properties of their solutions, then it might be expected that their inclusion, as sub-systems in interacting systems [Citation1, Citation4, Citation6], might lead to substantial changes in the prediction of the population levels at some time t. This issue shall not be taken up in this paper, but will appear in a future work.
The single-population models, considered here, belong to the class of mathematical representations which depend on two parameters. If we let be the population at time, t, and denote the two parameters as
, then the population dynamics is given by the relation
(1)
(1) where
are positive. We also restrict
such that
(2)
(2) Given a function
, our basic methodology is to first determine the solution
, and then calculate the times it takes to reach specific population levels. The comparisons will be among the times it takes each model to achieve a given population value.
The paper is organized as follows. Section 2 presents the five single-population models and gives references to their origin and/or detailed discussions of their formulations and applications. In Section 3, we provide a set of initial conditions, to be applied to each model differential equation, such that the relevant solutions may be directly compared with each other. This allows the direct expression of the particular parameters, , of a given model to be expressed in terms of the initial conditions. Remarkably, the exact solutions to each of the single-population differential equations can be calculated and these are presented in Section 4. Section 5 gives the results for
, where
is the time taken for the population to reach the value
, that is,
. Finally, we end the paper with a discussion of the essential results, several general comments, and possible extensions of our current work.
2. Five single-population models
We consider five single-population models listed below, along with some brief comments and references to their origins. Several of them are widely used not only as models for single-population growth, but also as representations of sub-populations in systems containing the interactions among multi-populations [Citation1, Citation4, Citation6].
In the work to follow, all parameters are assumed to be positive and independent of time.
Model A: Perhaps the simplest of the models is that given by the expression
(3)
(3) While it is widely used in multi-population interacting systems [Citation1, Citation4, Citation6], it generally has validity only for situations where one has control over the introduction of new members of the population. An example of the ideal case for this type of equation is in the modelling of chemostat-related phenomena [Citation7].
Model B: One of the best-known models is the logistic equation [Citation6, Citation15],
(4)
(4) There are many single-population systems for which its solutions provide an accurate representation of the experimental data [Citation6, Citation15].
Model C: The so-called combustion equation is
(5)
(5) The full details of the interesting mathematical features of its solutions are given in the book by O'Malley [Citation13].
Model D: All of the previous differential equation models have polynomial functions on their right sides. Another possible single-population model which does not have this feature is
(6)
(6) This representation is based on previous work by Mickens [Citation11] related to travelling wave solutions for a nonlinear reaction–diffusion partial differential equation.
Model E: Similarly, a second non-polynomial model is
(7)
(7) This is the Gompertz equation and has been used extensively to represent the growth of tumours [Citation16].
Inspection of Equations (Equation3(3)
(3) )– (Equation7
(7)
(7) ) allows the following conclusions to be reached:
Each of the models depend on just two, a priori, unknown parameters.
Except for Model A, all the related differential equations have two fixed-points or constant solutions, one at
, a second at
In general, we expect the ‘axiom of parenthood’ [Citation9] to hold. Mathematically, this implies that in Equation (Equation1
As will be shown, in the next section, exact solutions may be calculated for all five models [Citation5, Citation12, Citation14].
Using standard results from calculus and differential equations [Citation3, Citation12, Citation14], the solutions to all five models have the following property. If
3. Methodology
To compare the time evolution properties of the five single-population models, an a priori set of mathematical conditions must be selected and all of the models must be required to satisfy these requirements. We call these requirements ‘normalization conditions'.
For our investigations, are taken to be the same for the five models. These three quantities are defined as follows:
(10)
(10)
The basic methodology is to solve each mathematical model, , for its solution,
, and then use the relations
(11)
(11) to express
in terms of
.
4. Exact solutions
All of the five single-population differential equations can be solved exactly, although, for one case, the solution is implicit. In the calculations to follow, we only provide a brief outline of the procedure for determining the various solutions and give references to where the full details may be found. In most cases, only an elementary knowledge of calculus and differential equations are needed to derive the required solutions.
Model A: . The differential equation and general solution are given, respectively, by the expressions [Citation5, Citation12, Citation14]
(12a)
(12a)
(12b)
(12b) Imposing the conditions in Equation (Equation11
(11)
(11) ) gives
(13)
(13) and solving for
produces the result
(14)
(14) Therefore, the model differential equation and solution are given by the following
(15a)
(15a)
(15b)
(15b)
Using a similar set of procedures, the calculations for the other single-population models may be completed. Only the final results will be given for the remaining models.
Model B: . For this case, we have
(16)
(16) and
(17a)
(17a)
(17b)
(17b)
Model C: . This ordinary differential equation has an exact solution expressible in terms of the Lambert-W function [Citation3]. However, a direct calculation, using the method of partial fractions [Citation12, Citation14], gives the implicit solution
(18)
(18)
Model D: . This differential equation can be transformed into a linear equation by means of the dependent variable change,
. Solving the resulting differential equation, transforming back to x, and expressing
in terms of
gives the following results:
(19a)
(19a)
(19b)
(19b)
Model E: . This equation is the Gompertz model for single-population growth [Citation16]. It and its exact solution may also be expressed in terms of
; they are given by the forms
(20a)
(20a)
(20b)
(20b) where
is
(21)
(21)
5. values
Close examination of the solutions to the five models shows that they all can be rewritten as follows:
(22)
(22)
that is, t is a given function, H, of the indicated variables.
Our comparison criterion consists of selecting a particular set of (reasonable?) numerical values for the parameters , and then determine the time for each population model to achieve this value. Thus, if the value N is chosen for x, the corresponding,
, is
(23)
(23) For our analysis, we selected the following values,
(24)
(24) that is the initial, starting population is composed of
individuals, the limiting or maximum population is
individuals, and the initial (
) birth rate is 10/year. From this choice, we see that the time unit is the year, and the initial population is 1% of the maximum population.
The calculation of the , the time in years to achieve a population of N individuals is straightforward to determine and are listed in Table . For N, we selected the values,
; 1000;2500.
Table 1. Times to achieve population N for each model.
Inspection of Table allows the following conclusions to be reached:
The time for the population to double from its initial value of 100 is essentially the same for all the models. They range from a low of about 5 years to a high of 10 years.
Model C has the most rapid increase of any of the models, achieving a population level of approximately 50 times its initial value in 10 years. This is a general property of combustion-type models [Citation13].
The slowest model to reach a given level of population size is Model A.
The logistic equation is the second fastest model, although it takes approximately four times as much in comparison with the combustion equation, Model C.
If we consider just the three standard models, for single-population growth (Models A, B, and E), then expressed in terms of the time to reach a particular level of population, they can be ranked as follows:
In summary, we may conclude that the five single-population models considered in this study have a wide range of dynamic times associated with the evolution of their respective populations.
Another, somewhat related issue is the intrinsic time scales associated with the mathematical modelling of a system by ordinary differential equations. These intrinsic time scales are calculated by combining powers of the system parameters and the initial conditions in ways such that the new quantities have the physical unit of time [Citation8, Citation10]. In general, it is expected that these scales will provide useful information and insights into the duration of important dynamic features of the system. However, for ‘complex’ systems or systems having many intrinsic time scales, it can be a daunting task to interpret the actual significance of the various time scales.
We now calculate the intrinsic time scales for each of the five single-population models. Their numerical values are evaluated using the parameters listed in Equation (Equation24(24)
(24) ). However, the explicit determination of the time scales will only be illustrated for Model A. Figure presents the numerical values of these scales for all models.
Figure 1. Comparison of intrinsic time scales for the five models. The horizontal scale is time (years).
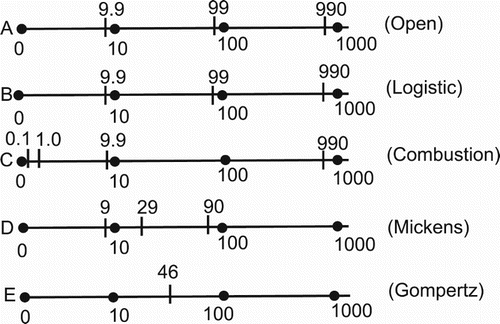
For Model A, there are five parameters to consider: Π, μ, ,
, and
. Denoting the physical dimensions of a quantity Q by
, we have
(25)
(25) where
and T have the units of population number and time. Therefore, it follows that
(26)
(26) From these results, the four possible intrinsic time scales are
c
(27a)
(27a)
(27b)
(27b)
(27c)
(27c)
(27d)
(27d) Inspection of these relations shows that the following inequalities hold
(28)
(28) under the condition
.
One interesting fact should be indicated. Of all five models, only the Gompertz equation has a single, fundamental intrinsic time scale and it is given by
(29)
(29)
A detailed examination of Figure allows the following conclusions to be reached.
All five models exhibit short-time dynamics characterized by time scales in the range of approximately 10–50 years. For Models A, B, C, and D, this value is essentially 10 years.
Three of the Models (A, B, D) have an intermediate intrinsic time scale in the interval 90–100 years.
Only Models (A, B, C) have long-time dynamics characterized by intrinsic time scales of 990 years.
As noted previously, the combustion model has fast dynamics, that is, it achieves a population value near its maximum value quickly, in the order of several 10s of years.
Inspection and comparison of the results given in both Table , and Figure does not allow any easy conclusions to be drawn as to possible relations or connection between the
6. Conclusions
The results of this paper show that for the particular five single-population models we have studied, while there are certain overall regularities, in general, any two given models may have major differences in the numerical values for the times to achieve a given level of the population; see Table . Consequently, we should be very cautious in our use of these models when they are incorporated within the (mathematical) structure of complex interacting population models. It should not be assumed that for a given sub-population each of the single-population models given here are interchangeable. In other words, the populations growth dynamics ‘ greatly affects model outcomes, and alternative model structures can result in very different predicted effects
’ [Citation2].
In a future paper, we will present results related to the issues of the current paper, but investigated within the context of three parameter, single-population models. Two explicit examples are
(30)
(30) and
(31)
(31) where all the indicated parameters are positive.
Disclosure statement
No potential conflict of interest was reported by the author.
Additional information
Funding
References
- H.T. Banks and C. Castillo-Chávez (eds), Bioterrorism: Mathematical Modeling Applications in Homeland Security, SIAM, Philadelphia, 2003.
- S.R. Beissinger and M.I. Westphal, On the use of demographic models of population viability in endangered species management, J. Wildlife Manag. 62 (1998), pp. 821–841.
- J.M. Borwein and R.M. Carless, Emerging tools for experimental mathematics, Amer. Math. Monthly 106 (1999), pp. 889–909.
- F. Brauer and C. Castillo-Chávez, Mathematical Models in Population Biology and Epidemiology, Springer, New York, 2001.
- I.S. Gradshteyn and I.M. Ryzhik, Table of Integrals, Series, and Products, Academic Press, New York, 1965.
- F.C. Hoppensteadt and C.S. Peskin, Modeling and Simulation in Medicine and the Life Sciences, 2nd ed., Springer, New York, 2002.
- S.S. Hsu, S. Hubbell, and P. Waltman, A mathematical theory for single-nutrient competition in continuous cultures of micro-organisms, SIAM J. Appl. Math. 32 (1977), pp. 366–383.
- H.E. Huntley, Dimensional Analysis, Dover, New York, 1967.
- G.E. Hutchinson, Introduction to Population Ecology, Yale University Press, New Haven, 1978, Chapter 1.
- R.E. Mickens, Mathematical Methods for the Natural and Engineering Sciences, World Scientific, London, 2004, Sections 1.3 and 1.4.
- R.E. Mickens, Wave front behavior of traveling wave solutions for a PDE having square-root dynamics, Math. Compt. Simul. 82 (2012), pp. 1271–1277.
- R.K. Nagle, E.B. Saff, and A.D. Snider, Fundamentals of Differential Equations, 7th ed., Pearson, Boston, 2008.
- R.E. O'Malley Jr., Thinking About Ordinary Differential Equations, Cambridge University Press, New York, 1997, Section 6.5.
- S.L. Ross, Differential Equations, Blaisdell, Waltham, MA, 1964.
- H.R. Thieme, Mathematics in Population Biology, Princeton University Press, Princeton, 2003, Chapters 5 and 6.
- C.P. Winsor, The Gompertz curve as a growth curve, Proc. Natl. Acad. Sci. 18 (1932), pp. 1–8.