
Abstract
Background: Variation in style length has been reported in Senecio vulgaris and has been associated with outcrossing rate.
Aims: To determine if (i) long styles lack germinated pollen on stigmas left to self-pollinate, (ii) successful self-pollination causes styles to stop elongating and shrink in length and (iii) seed set increases with the amount of pollen deposited on stigmas.
Methods: Determined germinated self-pollen on stigmas of long and short styles after auto-self-pollination; scored style length over 48 h in self-pollinated and non-pollinated florets; recorded seed set after placing different amounts of pollen on stigmas.
Results: Most long-styled florets had zero or low amounts of germinated pollen on stigmas in contrast to most short-styled florets. Styles initially elongated to the same length in self-pollinated and non-pollinated florets, then shrank in length in self-pollinated florets while continuing to elongate in non-pollinated florets. Seed set increased with number of pollen grains deposited on stigmas.
Conclusions: Successful self-pollen deposition and/or germination on stigmas of S. vulgaris are indicated by presence of short styles, whereas the opposite is indicated by presence of long styles in florets left to self-pollinate. Self-pollination causes styles to shrink after initially elongating. Seed set is dependent on the amount of pollen deposited on stigmas.
Introduction
Flowering plants exhibit various floral mechanisms to reduce the likelihood of self-pollination within cosexual flowers. Protandry, where anther dehiscence precedes stigma receptivity within a flower, is a mechanism prevalent in the Asteraceae, which comprises ca. 24,000 species (Funk et al. Citation2009). Many species of Asteraceae produce either radiate or discoid flower heads (capitula) (Gillies et al. Citation2002). Radiate capitula are composed of an outer whorl of ray florets, which are female (pistillate) or sterile, surrounding a central disc of hermaphrodite disc florets. In contrast, discoid (non-radiate) capitula are composed entirely of hermaphrodite disc florets. In both radiate and non-radiate capitula, all hermaphrodite disc florets exhibit protandry. In subfamily Asteroideae, which comprises the majority of species of Asteraceae, pollen is released from anthers positioned near the apex of the corolla tube above the style, and as the style elongates, pollen is pushed out of the floret by sterile hairs at the end of the closed and unreceptive bifid stigma (Leins and Erbar Citation2006). The stigma then opens and is receptive to pollination at which point self-pollen may fall on its receptive surface, be picked up by a pollinator, be blown away or fall from the stigma on to other parts of the capitulum.
In self-incompatible Asteraceae, self-pollen fails to germinate on stigmas of the same plant, but its deposition can cause pollen or stigma clogging (Barrett Citation2002), thus reducing access of cross-pollen to stigma surfaces and the likelihood of a floret to set seed. In contrast, in self-compatible species, self-pollen is expected to germinate rapidly on the stigma in the same floret, or a neighbouring floret, and lead to rapid self-fertilisation and prevention of outcrossing, although as far as we know this has never been investigated in detail in any self-compatible Asteraceae.
Here we focus on the pollination biology of the self-compatible species Senecio vulgaris L. (Asteraceae, subfamily Asteroideae) for which both radiate and discoid variants are recognised. In populations containing both variants, radiate plants always exhibit a higher outcrossing rate (3–35%) relative to non-radiate plants (normally <1%) (Marshall and Abbott Citation1982, Citation1984a). This has been attributed to radiate capitula being more attractive to pollinators (Abbott and Irwin Citation1988) and because their female ray florets outcross at higher rates than hermaphrodite disc florets (Marshall and Abbott Citation1984b). Moreover, recently it was reported (Irwin et al. Citation2016) that radiate capitula also contain a higher frequency of long-styled florets, i.e. with stigmas exserted well above the rim of the corolla tube relative to those of short-styled florets (see figure in Irwin et al. Citation2016), and it was considered that this might be a further factor boosting the outcrossing rate of the radiate variant. Trow (Citation1912) previously noted that plants of S. vulgaris bearing capitula in which all florets have long styles fail to set seed when left to auto-self-pollinate, although set good seed when manually self-pollinated. In their recent study, Irwin et al. (Citation2016) showed that 24 h after self-pollen was applied to stigmas in emasculated florets, styles were much shorter than those in florets that were emasculated but not self-pollinated (i.e. left non-pollinated). Whereas non-pollinated styles lacked germinated pollen on their stigmas, 90% of self-pollinated styles had germinated pollen on their stigmas. Absence of germinated pollen on stigmas of the remaining 10% of self-pollinated styles was likely due to them being washed off accidentally during the analytical procedure. Though the results of Irwin et al. (Citation2016) strongly indicated that style length variation in S. vulgaris is dependent on whether self-pollen deposition and germination had occurred, the development of style length and how it is affected by these processes remained unclear. It was proposed that styles might stop elongating after self-pollen deposition and germination on stigmas, but continue to elongate if either of these processes fails. Alternatively, styles might rapidly shrink in length following self-pollen deposition and germination on stigmas, but continue to elongate or not shrink in length when neither process occurs.
Figure 1. Stages in style length development during phases of elongation and contraction in Senecio vulgaris. (A) Elongation phase scores: 1, style remains inserted in corolla tube just after floret has opened; 2, closed stigma protrudes from corolla apex; 3, style exserted just above rim of corolla tube with stigmatic lobes not completely unfurled; 4, style exserted from corolla tube with stigmatic lobes completely unfurled just above corolla rim; 5, style fully elongated with stigmatic lobes unfurled above corolla rim. (B) Contraction phase scores: 4, as 4 above; 3, similar to 3 above; 2, as 2 above, but stigmatic lobes remain partially unfurled with tips protruding from corolla tube apex; 1, as 1 above, but stigmatic lobes remain partially unfurled though fully reinserted into corolla tube.

In the study reported here, we examine further the effects of self-pollination and self-pollen germination on style length in S. vulgaris. First, we test whether long styles lack germinated self-pollen on their stigmas, in contrast to short styles, when florets are left to self-pollinate. We show that most long-styled florets have either zero or a low number of germinated pollen grains on their stigmas in contrast to higher amounts present on stigmas of most short-styled florets. Second, we examine the effect that self-pollination has on style length recorded at intervals from the time that florets open to 48 h later and show that shorter style length following self-pollination (Irwin et al. Citation2016) results from shrinkage in style length. Finally, we examine the relationship between seed set and the amount of pollen deposited on stigmas and show that seed set increases as the number of pollen grains deposited on stigmas increases from 1 to a maximum of 10.
Materials and methods
Plants of both the radiate (S. vulgaris var. hibernicus) and non-radiate variants of S. vulgaris were grown from seed in an illuminated greenhouse. Seed came from inbred lines derived (following ca. 10 generations of selfing) from plants that occurred in populations containing both variants at Edinburgh, Scotland, Cardiff, Wales, and New Broughton, North Wales, UK (see Marshall and Abbott Citation1984a, Citation1984b). Cultivation conditions were the same as those described in Irwin et al. (Citation2016).
Amount of self-pollen germinating on stigmas of short and long styles
Three plants of each variant from each of the three populations were treated as follows. Florets in several capitula per plant were individually marked with indelible ink using a fine pen at the time when they had just opened. After 24 h, marked florets were removed from capitula using watchmaker forceps and classified according to floret position, i.e. from the outer or inner whorls of the capitulum, and also for style length, i.e. short or long (after Irwin et al. Citation2016), determined using a binocular microscope. In the case of radiate plants, the outer florets represented ray florets, whereas inner florets represented disc florets. In this way, 207 and 247 florets were examined from the radiate and non-radiate variants, respectively. Following classification, florets were fixed in formaldehyde alcohol acetic acid (10%, 50%, 5% plus 35% double distilled H2O) before removing styles and counting the number of germinated pollen grains on stigmas using decolourised aniline blue staining and viewing with a Leica fluorescence microscope fitted with an ultra-violet light filter (method modified from Martin Citation1958).
Data (Table S1) were analysed in two ways. First, a comparison was made of the frequency of long and short styles having zero or more than zero germinated pollen grains on their stigmas. Second, a comparison was made of the mean number of germinated pollen grains present on stigmas of long and short styles. In the second form of analysis, data were subjected to analysis of variance (ANOVA) using the generalised linear model procedure (Proc GLM) of the SAS statistical software v.93 (SAS Citation2011).
Effect of self-pollination on style length
Another five radiate and five non-radiate plants derived from the Cardiff population were chosen for study. Two or three young capitula on each plant were selected and divided vertically in half by inserting a small square of stiff paper between the unopened florets. Several florets in each half of each capitulum were marked with a fine pen to facilitate subsequent recognition. Only disc florets of radiate capitula and from the inner whorls of non-radiate capitula were chosen for treatment. Treatment involved leaving the florets in one half of a capitulum to self-pollinate, while pollen was removed from all florets in the other half as it was released. Removal of pollen was conducted using a pipette connected to a filter pump before germination on stigmas could take place.
Immediately following treatment (0 h) and then at intervals of 6, 22, 24 and 48 h, style length in each floret was scored according to its stage of development as shown in . Irwin et al. (Citation2016) had previously shown that a difference in style length between self- and non-pollinated florets was apparent 24 h after treatment began. We wished to determine if a difference was also apparent at earlier and later stages following treatment and chose the times of 6, 22 and 48 h for this purpose. Because florets were followed at intervals after treatment, it was possible to score style length during the phases of elongation and subsequent contraction (in cases where this occurred), thus distinguishing between similar scores during these two phases of development ().
Data (Table S2) in the form of mean scores per capitulum over treatments (self-pollinated vs. non-pollinated), variants, times and plants were subjected to maximum likelihood estimates of parameters in linear mixed-effects models using the lmer function in the lme4 package for R (Bates et al. Citation2015). In these models, variant (V), pollination treatment (PT) and time at which mean style score was recorded (T) were considered as fixed effects while plant was treated as a random effect.
Amount of pollen required for seed set per floret
Five radiate and five non-radiate plants derived from the Cardiff population were chosen for study. Several capitula on each plant were emasculated prior to anthesis, using the method described by Irwin et al. (Citation2016). After emasculation, capitula were enclosed in paper bags made from lens tissue and left for 72 h, at which point styles were fully elongated and stigmas were receptive to pollen. Emasculated capitula were checked under a dissecting microscope (×20 magnification) and those with residual pollen were discarded. From each of the remaining emasculated capitula, several florets were removed to allow easier manipulation of the remaining florets. Mature non-emasculated capitula that were positioned close to emasculated ones on the same plant were also removed to reduce the risk of unintended pollination. Fresh pollen was collected from other non-emasculated capitula on each plant for self-pollination of emasculated florets and also from one donor plant of the other variant for cross-pollination.
Pollen was applied individually to stigmas within emasculated florets (under a × 20 dissecting microscope) using a single eyelash glued to the end of a cocktail stick. It was not possible to control rigorously for the exact site on the stigma where pollen was deposited; however, every attempt was made to place pollen grains on the receptive surface. On each plant, stigmas within three emasculated capitula were pollinated with either one, between 2 and 5, or between 6 and 10 self-pollen grains, respectively. The process was then repeated on another three emasculated capitula on the same plants using cross-pollen from the other variant. In radiate capitula, both ray and disc florets were subjected to the same pollination treatment. Following pollination, capitula were rebagged and left to set seed.
Data (Table S3) in the form of the proportion of pollinated florets per capitulum to set seed with self- or cross-pollen across the three pollination treatments and five plants per variant were subjected to ANOVA using the generalised linear model procedure (Proc GLM) of the SAS statistical software v.93 (SAS Citation2011).
Results
Amount of self-pollen germination on stigmas of short and long styles
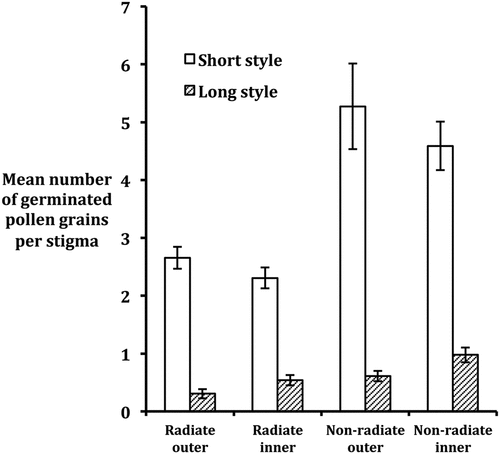
The frequency of styles with zero germinated pollen on their stigmas was much greater for long than short styles (, data pooled over floret positions, plants and populations). This was evident for both radiate and non-radiate variants. In the radiate variant, ca. 77% of long styles compared with only ca. 11% of short styles had zero germinated pollen on their stigmas. Similarly, in the non-radiate variant ca. 71% of long styles compared with only ca. 13% of short styles lacked germinated pollen on their stigmas. Although germinated pollen was present on stigmas of some long styles, very few of such stigmas had more than three germinated pollen grains on them (<5% in both variants). When comparisons were made between outer and inner florets, no significant difference was evident for frequency of styles lacking germinated stigmas on their stigmas for either long styles in the non-radiate variant or short styles in both variants (). However, there was a significant effect of floret position for long styles in the radiate morph with a greater than expected number of long styles lacking germinated pollen present among outer (ray) florets than inner (hermaphrodite) florets.
Table 1. Frequency (%) of long and short styles with 0, 1, 2, 3 or >3 germinated pollen grains on their stigmas in the radiate and non-radiate variants of Senecio vulgaris in Scottish and Welsh populations.
Table 2. Frequency (%) of styles (pooled over populations) with 0, 1, 2, 3 or >3 germinated pollen grains on their stigmas of Senecio vulgaris in Scottish and Welsh populations.
In terms of mean number of germinated pollen grains on stigmas, a much higher mean was evident for short than long styles in both the radiate and non-radiate variants (; F1, 46 = 46.18, P < 0.0001 in ANOVA, Table S4). It was also evident that the mean amount of germinated pollen on stigmas of short styles was significantly greater in the non-radiate than in the radiate variant [F1, 46 = 9.63, P = 0.0033 for difference between variants; F1, 46 = 5.23, P = 0.0268 for variant (V) × style length (SL) interaction, Table S4]. However, floret position (outer vs. inner) had no apparent effect on mean number of germinated pollen on stigmas in either short- or long-styled florets (, Table S4). The latter was a rather surprising finding for the radiate variant, given the higher proportion than expected of long styles lacking germinated pollen on their stigmas in the outer floret fraction (). ANOVA (Table S4) confirmed there was no effect of population on mean number of germinated pollen grains on stigmas, and that none of the interactions between the four main effects investigated, apart from V × SL, was significant.
Figure 2. Mean number of germinated pollen grains present on stigmas of styles in long- and short-styled florets of Senecio vulgaris in Scottish and Welsh populations. Standard errors are indicated as bars.
Effect of self-pollination on style length
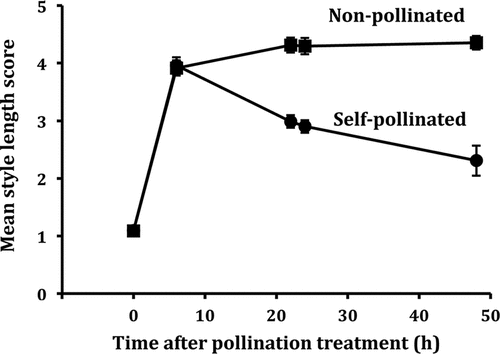
At the time when pollen was either removed from florets or florets were left to self-pollinate (0 h), styles remained inserted in florets that had just opened (mean style score equalled ca. 1 across both variants, and ). After 6 h, mean style length (across both variants) had increased to 3.91 and 3.96 in non-pollinated and self-pollinated florets, respectively (). Thus, at this stage styles in both non-pollinated and self-pollinated florets were exserted from the corolla tube with stigmatic lobes completely unfurled just above the corolla rim (). Over the next 14–16 h, most styles continued to elongate in non-pollinated florets with a mean style length score of ca. 4.3 recorded after 24 h. Thus, by this time, styles in many non-pollinated florets were fully elongated above the corolla rim (stage 5, ), while others remained at earlier stages of elongation (stages 3 and 4). In contrast, in florets left to self-pollinate styles shrank in length between 6 h and 22–24 h with mean style length score reduced to 2.9 after 24 h. Over the next 24 h, style length remained unchanged in non-pollinated florets (mean style length score equalling 4.35 after 48 h), while styles in self-pollinated florets continued to shrink in length (mean style length score = 2.31, with only stigma tips protruding from the corolla apex). The same effect of self-pollination on style length development was exhibited in both variants (Figure S1). Linear mixed-effect model analysis using lmer in the lme4 package for R showed no significant effect for variant, but highly significant effects for pollination treatment and time (χ2(1) = 24.68, P < 0.0001; χ2(4) = 97.29, P < 0.0001, respectively) and the interaction between pollination treatment and time (χ2(4) = 30.96, P < 0.0001).
Figure 3. Mean style length scores of non-pollinated and self-pollinated florets of Senecio vulgaris at time of treatment (0 h) and at intervals of 6, 22, 24 and 48 h afterwards. Standard errors of means are indicated by bars.
Amount of pollen required for seed set per floret
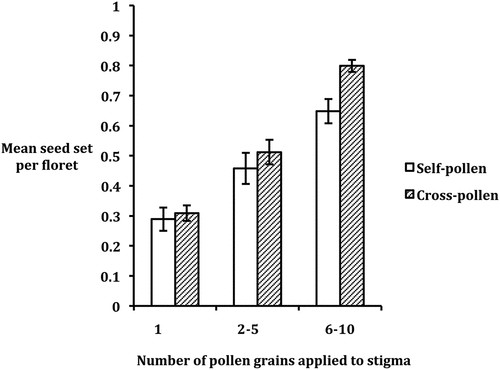
The probability of a floret producing a seed increased with the number of self- or cross-pollen grains deposited on its stigma (). Mean seed set per floret (maximum of 1 as there is one ovule per floret in Asteraceae) across both variants and pollen types was 0.299 when only one pollen grain was placed on a stigma, rose to 0.485 with 2–5 pollen grains on the stigma and further increased to 0.723 with 6–10 pollen grains per stigma. ANOVA of the total data set (Table S5) showed that the effect of pollen number on seed set per floret was highly significant (F2, 40 = 16.65, P < 0.0001), but there was no significant effect of variant (radiate vs. non-radiate) or pollen type (self- vs. cross-pollen). With self-pollen the increase in seed set with pollen number was not as marked in the radiate variant as in the non-radiate variant, with no increase recorded in the radiate variant when pollen number was raised from 1 to 2–5 pollen grains per stigma (). There was also an indication that cross-pollination resulted in greater seed set than self-pollination (across both variants) when 6–10 pollen grains were applied to stigmas (). However, neither of these effects was significant according to the ANOVA, which showed that all interactions between variant, pollen type and pollen number were non-significant (Table S5).
Table 3. Mean seed set per floret of the radiate and non-radiate variants of Senecio vulgaris following placement of 1, 2–5 or 6–10 grains of self- or cross-pollen per stigma.
Figure 4. Mean seed set per floret (across both radiate and non-radiate variants of Senecio vulgaris) following placement of 1, 2–5 or 6–10 grains of self- or cross-pollen per stigma. Standard errors are indicated as bars.
Discussion
The results reported here advance our understanding of the effect of self-pollination on style length development and also seed set in self-compatible species of Asteraceae. They show that long styles tend to develop and persist in S. vulgaris when pollen fails to germinate on stigmas in florets left to self-pollinate. In contrast, short styles tend to develop after pollen successfully germinates on such stigmas. Furthermore, the results make clear that short styles develop due to style shrinkage occurring between 6 and 48 h after self-pollination, whereas long styles develop due to continued elongation up to 22 h if self-pollination is prevented and maintain their length up to at least 48 h after the start of this treatment. Interestingly, styles of both self-pollinated and non-pollinated florets elongated to a similar length up to 6 h after treatments began, at which time their stigmas were completely unfurled and positioned just above the rim of the corolla tube. Finally, our study of the effect of pollination on seed set showed that seed set increases as the number of pollen grains deposited on stigmas is raised from 1 to a maximum of 10.
Amount of self-pollen germinating on stigmas of short and long styles
Examination of the amount of self-pollen germination on stigmas of short and long styles in S. vulgaris revealed a clear, though not complete, relationship between style length and presence of germinated pollen on stigmas. Thus, most long styles tended to have no germinated pollen on their stigmas, whereas most short styles had stigmas with normally one or more germinated pollen grains on them. This was apparent in both the radiate and non-radiate variants of S. vulgaris (). The fact that the relationship was not complete, i.e. between approximately 22% and 29% of long styles had some pollen germinating on their stigmas (dependent on variant), while between approximately 11% and 13% of short styles had pollen-free stigmas, could be explained in different ways. For example, it is feasible that self-pollen was late to germinate on stigmas of those long styles with germinated pollen on their stigmas, in which case these styles may have elongated and been sampled before self-pollen germination had an effect on style length. For short styles lacking germinated pollen on their stigmas, it is possible that pollen was lost from these stigmas during the analytical process. Whatever the actual causes of the incomplete relationship, it is clear from our results that long styles generally do not have germinated pollen on their stigmas whereas short styles do.
Our results also showed that in radiate, but not non-radiate capitula, a higher proportion than expected of long styles lacking germinated pollen on their stigmas occurred in the outer floret fraction than in the inner floret fraction (). This finding was not unexpected, given that ray florets lacking anthers comprise the outer floret fraction of radiate capitula and become self-pollinated only when pollen is transferred to their stigmas from an inner (hermaphrodite) floret within the same or a neighbouring capitulum. Thus, the probability of self-pollination of styles in ray florets is likely to be lower than in hermaphrodite (inner) disc florets, and, consequently, ray florets will be expected to contain a greater proportion of long styles lacking germinated pollen on their stigmas.
In addition to determining the frequencies of long- and short-styled florets either lacking or possessing germinated pollen on their stigmas, we estimated the average amount of germinated self-pollen present on stigmas of the two style types. It was evident that the mean number of germinated pollen grains present on stigmas of short styles was considerably higher than on those of long styles (). Furthermore, the mean number present on stigmas of short styles in the non-radiate variant was more than double that recorded in the radiate variant, irrespective of floret position. This latter finding is of interest as it indicates that the probability of self-pollen deposition and germination on stigmas of hermaphrodite florets is lower in the radiate than non-radiate variant for reasons currently unknown. Taken overall, a surprising feature to emerge from the results is that very little self-pollen produced by a hermaphrodite floret of either variant successfully germinates on the stigmas of florets. Ross and Abbott (Citation1987) estimated that on average the number of pollen grains produced by a hermaphrodite inner floret of the radiate or non-radiate variant exceeds 240. However, according to our results only 1.5 (SE = 0.40) and 3.08 (SE = 0.34) pollen grains germinated on average on the stigmas of styles in radiate and non-radiate hermaphrodite florets, respectively, and that these values fell to 1.09 (SE = 0.00) and 2.19 (SE = 0.24), respectively, on the stigmas of outer florets. Thus, only a small proportion of the pollen produced by a hermaphrodite floret germinates on the stigmas of a floret in the same capitulum and this proportion is lower in the radiate than the non-radiate variant. The greatest number of germinated pollen recorded on a particular stigma in the present study was 20. If due to physical constraints this value represents a ‘ceiling value’ for number of germinated pollen on a stigma, it remains the case that the actual number of self-pollen grains which germinate on a stigma is normally far lower than could potentially germinate.
Effect of self-pollination on style length
A clear result to emerge from our study of the effect of self-pollination on style length was that styles in both self-pollinated and non-pollinated florets elongated to the same length 6 h after treatment began (). At this stage, styles had developed to the point where they were exserted from the corolla tube with stigmatic lobes completely unfurled just above the corolla rim. Thereafter, however, self-pollinated styles shrank in length, while non-pollinated styles continued to elongate until 22 h and then retained their longer length up to 48 h when the experiment ended. This pattern of response was evident in both radiate and non-radiate variants of S. vulgaris (Figure S1).
A previous study by Irwin et al. (Citation2016) showed that self-pollination of emasculated florets resulted in production of short styles, whereas non-pollination caused long styles to be produced after 24 h. However, their study did not determine whether short styles were produced due to failure to elongate following self-pollination and/or because they shrank in length after such treatment. Our results make clear that short styles develop because they stop elongating between 6 and 22 h after self-pollination and instead shrink in length during this period. It was notable that up to 6 h after self-pollination self-pollinated styles elongated to the same length as non-pollinated styles and only after 6 h did they stop elongating and begin to shrink in length.
Amount of pollen required for seed set per floret
In view of our finding that only a small proportion of pollen produced by a hermaphrodite floret germinates on the stigmas of such florets when plants are left to auto-self-pollinate, it was of interest to establish how much pollen needs to be deposited on a stigma to insure seed set in these single-seeded florets and whether the probability of seed set increases with the amount of pollen deposited. Our study showed that average seed set was only 30% when one pollen grain was deposited manually on a stigma, increased to approximately 50% when 2–5 pollen grains were deposited and further rose to approximately 72% with 6–10 deposited. This pattern of response was not significantly affected by either variant (radiate vs. non-radiate) or pollen type used (self- vs. cross-pollen) (Table S5). Clearly, the probability of seed set increases with the amount of pollen deposited on a stigma under the conditions employed in our experiment, and it is possible that further increases in seed set would arise if more pollen were deposited on stigmas.
In our study of the amount of pollen occurring on stigmas of long and short styles 24 h after a floret opened, we recorded only germinated pollen on stigmas because the analytical procedure used to detect pollen was very likely to wash off much of the pollen that failed to germinate. Furthermore, in our study of the amount of pollen required for seed set, we did not determine what proportion of pollen grains that were manually deposited on a stigma either germinated or failed to germinate. Consequently, we are not able to say how much self-pollen is normally deposited naturally on to the receptive surface of the stigma in a hermaphrodite floret at the time when the stigma opens or what proportion of pollen deposited actually germinates. It is possible that a considerable amount of self-pollen is deposited during auto-self-pollination, of which only a small proportion germinates. If this is the case, the presence of pollen that fails to germinate could reduce access of cross-pollen to the stigma and therefore the potential to outcross. Alternatively, only a small amount of pollen released from anthers might be deposited on a stigma of which a reasonable proportion germinates. If this is the case, opportunities for cross-pollination and outcrossing might be increased.
Further considerations and future directions
More than a century ago, Trow (Citation1912) reported that some plants of S. vulgaris produce capitula containing long-styled florets in contrast to other plants having capitula composed of short-styled florets. Trow noted that long-styled florets failed to set seed unless pollinated manually or by a pollinator, suggesting that auto-self-pollination failed in these florets. Recently, Irwin et al. (Citation2016) reported that the frequency of long-styled florets in capitula was positively associated with outcrossing rate in S. vulgaris. Radiate plants, which exhibit greater outcrossing than non-radiate plants, were shown to produce capitula containing a higher frequency of long-styled florets. Furthermore, Irwin et al. (Citation2016) showed that in emasculated florets manual self-pollination causes short styles to develop after 24 h, whereas non-pollination results in development of long styles. Our studies have developed the work of Trow (Citation1912) and Irwin et al. (Citation2016) further and showed that the majority of long styles in florets left to auto-self-pollinate have no germinated pollen on their stigmas, while short styles normally have one or more germinated pollen grains on their stigmas. Our results further showed that short styles result from cessation of style elongation and shrinkage in style length between 6 and 48 h after self-pollination. In contrast, long styles develop when stigmas are not pollinated due to continued elongation between 6 and 22 h followed by retention of a longer length up to at least 48 h.
The fact that the radiate variant produces capitula with a higher frequency of long-styled florets than the non-radiate variant suggests that auto-self-pollen deposition and/or self-pollen germination are less successful in the radiate variant. This is to be expected to some degree as the radiate variant produces capitula that contain an outer whorl of ray florets that are male sterile (lack anthers). However, we found that the radiate variant also shows a higher frequency of long-styled florets than the non-radiate variant when comparisons are made only between the inner hermaphrodite disc florets of each variant. Currently, we are not able to explain the cause of this difference. Both variants produce, on average, similar amounts of pollen per inner hermaphrodite floret (Ross and Abbott Citation1987) and so we can rule out the possible effect that a difference in amount of pollen available for self-pollination might have on frequency of long-styled florets. It would seem that within the radiate variant self-pollen is deposited less efficiently on the stigma of a hermaphrodite floret or germinates less readily than in the non-radiate variant. We previously ruled out the possibility that the radiate variant might exhibit a weak form of self-incompatibility inherited from self-incompatible Senecio squalidus during its introgressive origin (Kim et al. Citation2008), which would prevent or delay self-pollen germination (Irwin et al. Citation2016). This was because 90% of manually self-pollinated styles were found by Irwin et al. (Citation2016) to have germinated pollen on their stigmas after 24 h and, further, because it is known that the radiate variant ultimately exhibits high seed set when left to self (Abbott et al. Citation1990). Future studies should aim to determine which causes are responsible for the difference in frequency of long-styled florets recorded between the radiate and non-radiate variants.
Although we cannot explain why auto-self-pollination within hermaphrodite florets is less successful in the radiate relative to the non-radiate variant, our results do explain why radiate capitula contain higher frequencies of long-styled florets 24 h after stigmas first become receptive (Irwin et al. Citation2016). Because self-pollen deposition and/or germination must be delayed, on average, for longer in radiate capitula, long-styled hermaphrodite florets will persist for longer in such capitula relative to non-radiate capitula. Consequently, 24 h after stigmas first become receptive to pollen a higher frequency of long-styled florets is expected to be present in radiate relative to non-radiate capitula.
Mazer et al. (Citation2016) recently highlighted the ubiquity of style length variation within and between plant taxa, emphasising its functional significance. In the case of S. vulgaris, our results suggest that style length variation in the species is a side effect of whether self-pollen deposition and germination has occurred or not. However, the development and maintenance of long styles when self-pollen deposition and/or germination have failed may increase the access of stigmas to pollinators, which in turn should increase the probability of a floret becoming pollinated and indeed cross-pollinated. Thus, style length variation in S. vulgaris, though a side effect of the success or failure of self-pollen deposition and germination, is nonetheless likely to be of functional significance and affects the probability of cross-pollination and outcrossing of a plant.
Future studies should determine whether what we have found in S. vulgaris occurs in other self-compatible Asteraceae. We consider that shrinkage in style length soon after self-pollination, but continued elongation in the absence of pollination, will likely be a feature of many self-compatible Asteraceae. Wagenius (Citation2004) has shown that in self-incompatible Echinacea angustifolia (Asteraceae) receptive styles are exserted from florets prior to pollination and remain exserted for up to 10 days if self-pollinated, but shrink in length within 24 h when cross-pollinated. The effect of self-pollination in this species can be considered equivalent to non-pollination in self-compatible S. vulgaris in that it results in failure of pollen germination on stigmas. Given the similar stylar response to failure of pollen germination on stigmas in these two species, we consider that the effect may be common in many other Asteraceae, at least in the large subfamily Asteroideae, which includes Senecio and Echinacea. In such self-compatible Asteraceae, the development and persistence of long-styled florets could be a common signal of failed self-pollen deposition and/or self-pollen germination on stigmas and serve to promote cross-pollination and outcrossing.
Although there are no previous investigations of the molecular mechanisms and cellular changes involved in style length development in Senecio, a study by Koning (Citation1983) on Gaillardia grandiflora (Asteraceae, subfamily Asteroideae) showed that epidermal cell elongation accompanies style elongation and is promoted by auxin but inhibited by gibberelins and ethylene. In flowers of Nicotiana tabacum, ethylene production increases rapidly following pollination (De Martinis et al. Citation2002) and plays an important role in triggering the degeneration of cells comprising the transmission tract of the style, thus aiding the passage of pollen tubes through this tissue (Wang et al. Citation1996). Ferradás et al. (Citation2014) have further shown that cellular changes indicative of programmed cell death are accelerated by pollination in stigmas and styles of kiwi fruit (Actinidia chinensis var. deliciosa). Although some molecular genetic mechanisms involved in style elongation (in Arabidopsis thaliana, Fuentes et al. Citation2012) and degeneration following pollination (in N. tabacum, Wang et al. Citation1996) are now known, they remain to be investigated in Senecio.
Table S5
Download MS Word (47.4 KB)Table S4
Download MS Word (57.6 KB)Table S3
Download MS Excel (39 KB)Table S2
Download MS Excel (38.9 KB)Table S1
Download MS Excel (37.4 KB)Figure S1
Download MS Word (65.6 KB)Acknowledgements
The authors thank David Forbes for laboratory assistance. They are also grateful to three anonymous reviewers for their constructive comments on earlier versions of the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplemental material
The supplemental data for this article can be accessed here
Additional information
Funding
Notes on contributors
John Love
John Love is an associate professor who conducts research into plant signalling and bioenergy.
Sean W. Graham
Sean Graham is a a professor who conducts research into plant systematics and evolution.
Judith A. Irwin
Judith Irwin is a senior scientist who conducts research on the genetics of vernalisation response in Brassica.
Paul A. Ashton
Paul Ashton is a professor of botany who conducts research on the ecological genetics of limes and sedges.
Francois Bretagnolle
Francois Bretagnolle is a professor who conducts research into diverse aspects of ecology and evolution.
Richard J. Abbott
Richard Abbott is emeritus professor of biology who conducts research on diverse aspects of plant evolution, especially plant speciation and the evolutionary consequences of hybridisation.
Related Research Data
References
- Abbott RJ, Irwin JA. 1988. Pollinator movements and the polymorphism for outcrossing rate at the ray floret locus in groundsel, Senecio vulgaris L. Heredity 60:295–298.
- Abbott RJ, Irwin JA, Forbes DF. 1990. Absence of a radiate morph bearing self-incompatible ray florets from Edinburgh populations of Senecio vulgaris L. Heredity 64:391–393.
- Barrett SCH. 2002. Sexual interference of the floral kind. Heredity 88:154–159.
- Bates D, Maechler M, Bolker B, Walker S. 2015. Fitting linear mixed-effect models using lme14. Journal of Statistical Software 67:1–48.
- De Martinis D, Cotti G, te Lintel Hekker S, Harren FJM, Mariani C. 2002. Ethylene response to pollen tube growth in Nicotiana tabacum. Planta 214:806–812.
- Ferradás Y, López M, Rey M, González MV. 2014. Programmed cell death in kiwifruit stigmatic arms and its relationship to the effective pollination period and the progamic phase. Annals of Botany 114:35–45.
- Fuentes S, Ljung K, Sorefan K, Alvey E, Harberd N, Østergaard L. 2012. Fruit growth in Arabidopsis occurs via DELLA-dependent and DELLA-independent gibberellin responses. The Plant Cell 24:3982–3996.
- Funk VA, Susanna A, Stuessy TF, Robinson H. 2009. Classification of compositae. In: Funk VA, Susanna A, Stuessy TF, Bayer R, editors. Systematics, evolution and biogeography of the compositae. Vienna: IAPT. p. 171–189.
- Gillies ACM, Cubas P, Coen ES, Abbott RJ. 2002. Making rays in the Asteraceae: genetics and evolution of radiate and discoid flower heads. In: Cronk QCB, Bateman RM, Hawkins JA, editors. Developmental genetics and plant evolution. London: Taylor & Francis. p. 233–246.
- Irwin JA, Ashton PA, Bretagnolle F, Abbott RJ. 2016. The long and the short of it: long-styled florets are associated with higher outcrossing rate in Senecio vulgaris and result from delayed self-pollen germination. Plant Ecology & Diversity 9:159–165.
- Kim M, Cui M-L, Cubas P, Gillies A, Lee K, Chapman MA, Abbott RJ, Coen E. 2008 . Regulatory genes control a key morphological and ecological trait transferred between species. Science 322:1116–1119.
- Koning RE. 1983. The roles of plant hormones in style and stigma growth in Gaillardia grandiflora (Asteraceae). American Journal of Botany 70:978–986.
- Leins P, Erbar C. 2006. Secondary pollen presentation syndromes of the Asterales – a phylogenetic perspective. Botanische Jahrbücher fur Systematik, Pflanzengeschichte und Pflanzengeographie 127:83–103.
- Marshall DF, Abbott RJ. 1982. Polymorphism for outcrossing frequency at the ray floret locus in Senecio vulgaris L. I. Evidence. Heredity 48:227–235.
- Marshall DF, Abbott RJ. 1984a. Polymorphism for outcrossing frequency at the ray floret locus in Senecio vulgaris L. II. Confirmation. Heredity 52:331–336.
- Marshall DF, Abbott RJ. 1984b. Polymorphism for outcrossing frequency at the ray floret locus in Senecio vulgaris L. III. Causes. Heredity 53:145–149.
- Martin FW. 1958. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technology 34:125–128.
- Mazer SJ, Moghaddasi A, Bello AK, Hove AA. 2016. Winning in style: longer styles receive more pollen, but style length does not affect pollen attrition in wild Clarkia populations. American Journal of Botany 103:408–422.
- Ross MD, Abbott RJ. 1987. Fitness, sexual asymmetry, functional sex and selfing in Senecio vulgaris L. Evolutionary Trends in Plants 1:21–28.
- SAS. 2011. Statistical analysis system for microsoft windows: v.9.3. Cary (NC): SAS.
- Trow AH. 1912. On the inheritance of certain characters in the common groundsel—Senecio vulgaris, Linn.—and its segregates. Journal of Genetics 2:239–276.
- Wagenius S. 2004. Style persistence, pollen limitation, and seed set in the common prairie plant Echinacea angustifolia (Asteraceae). International Journal of Plant Sciences 165:595–603.
- Wang H, Wu H-M, Cheung AY. 1996. Pollination induces mRNA poly(A) tail-shortening and cell deterioration in flower transmitting tissue. The Plant Journal 9:715–727.