
Abstract
Decline in mean temperature with elevation correlates with a more or less gradual change in forest floristics mediated by minimum temperature tolerances and competition, and in its diversity, well documented on wet equatorial mountains. On the mountains of monsoon tropical Asia, the predominance of a single wet and dry season offers opportunity to relate increasing cloud and rainfall seasonality with a departure in a zonation most clearly recognised on equatorial mountains. Observations indicate that the lowland to lower montane forest transition is gradual and constant in elevation throughout the monsoon tropical Asia region, and contrasts with the variable elevation at which forest structure and physiognomy, alternatively floristics alone, changes between lower and upper equatorial montane forests. Both ecotones are observed to correlate with the elevation and seasonality at which decline in temperature daily results in condensation of water vapour into cloud, resulting in cloud shadow, and fog therefore water supply where cloud penetrates the canopy. Seasonal variation in these climatic interactions influence forest structure and physiognomy and, by mediating the interaction between vegetation and substrate, also leads to zonation in soils and consequently floristics. Historical biogeographical factors also influence zonal floristics.
Introduction
Global variation in the elevational patterns of mountain vegetation is directly, or indirectly, determined by decline in temperature with elevation, climate seasonality, especially cloud and rainfall at regional scale, geology and relief which influence soils at local scale, the pool of available species and biotic interactions (Jenny Citation1941, 1994; Nagy and Grabherr Citation2009; Körner et al. Citation2011). In the humid tropics, forests clothe mountains from lowlands to tree line. Forest stature declines and becomes more uniform with elevation, while tree net primary production declines and shoot extension effectively ceases at a global tree line (Kitayama and Aiba Citation2002; Körner Citation2012), which coincides with a universal growing season mean temperature of ca. 6.5°C. A pattern of elevation zonation into physiognomically, structurally and floristically distinct belts and intervening ecotones has nevertheless been recognised on continuously wet equatorial mountains, based on studies in the Neotropics and Asia (Grubb et al. Citation1963; Whitmore Citation1984; Ashton Citation2003, Citation2014; which is broadly consistent with globally proposed bioclimatic elevation zones (Körner et al. Citation2011); but its applicability to the rainfall-seasonal tropics has yet to be examined. The following provide a review of field observations recorded from one of the three major tropical regions, Asia. It cannot be stressed enough that this review identifies correlations between forest and environmental variation, but the causes of such correlations can only be established by combined field and ex situ experiment (Körner Citation2012), so far still awaited in the Asian tropics.
Table 1. The global characteristics of forest zones by elevation in the equatorial and oceanic tropics, as observed in Asia (after Whitmore Citation1984; Ashton Citation2014).
Temperature alone predictably declines with elevation. Mean monthly variation in temperature varies less than 1°C in equatorial regions, whereas seasonal variation increases with latitude until, at the margin of the tropics, the difference between summer and winter monthly average can reach 20° in dry climates (Walter and Lieth Citation1960). Decline in mean summer, that is rainy season temperature with elevation, is universally ca.0.65°K/100 m ascent.
Precipitation, if seasonal, is concentrated in the tropics when the sun is overhead and temperature highest. This is the time when tropical easterly winds trend towards the climatic equator bringing moisture off the oceans. This Tropical Convergence moves annually, with the sun overhead, north to the Tropic of Cancer, and south to the Tropic of Capricorn, bringing a single rainy summer over most of tropical Asia. Thanks to the influence of low summer pressure over the high Tibetan Plateau, monsoon rain brought by the convergence is drawn to the north-east Himalayan foothills, beyond the geographical tropics at 27°N.
Condensation of water vapour into cloud occurs when saturated air is cooled. This may occur as low as 800 m in the tropics where moist air is daily drawn in over the sea, more generally at 1000–1200 m above closed forest in leaf, and at these same elevations under the convergence as it crosses the equator to higher latitudes in summer. The resulting cloud base and shadow coincides with an ecotone from lowland tropical soils where particulate organic matter is concentrated near the surface, to tropical montane soils in which it is carried down to depth by earthworms and other organisms: an ecological frontier as will be described below.
The forests of tropical Asia have been subject to several detailed accounts, and some to their associated physical environment; while the field accounts of several botanical ascents, from the equator to the tropical margin and from oceanic to continental regions and climates, have been published. The north-south disposition of the several mountain chains of the Asian tropics, each set in different levels of climatic continentality, combine to present an unparalleled opportunity to review the variability of montane forests and its relation with climate and soils throughout one tropical region.
Relatively low temperature seasonality in the tropics correlates with more sharply defined altitudinal zonation of forests than on temperate mountains (Janzen Citation1967). These zones have been defined by forest stature, structure, physiognomy and associated herbaceous life forms, notably epiphytes. There is an underlying continuous turnover of species along the altitudinal gradient, broadly correlated with a lapse rate in temperature between 0.4 and 0.7°C/100 m ascent (Richards Citation1996). Although species distributions transgress the zones thus defined to varying extent, each zone is distinguished by the relative abundance of a suite of species whose stature and physiognomy characterise it (Whitmore Citation1984; Ashton Citation2003). An account of montane forest zonation in one tropical region can provide a test of the extent to which the relation between forest structural, physiognomic and floristic zonation remain constant.
I earlier showed that zonal forest ecotones correlate with specific and differing changes in the physical environment, rather than intervals along a continuous climate gradient, such as proposed by Kira (Citation1977).
Forest zonation in aseasonal wet tropical Asia. In 1925, Charles Lane-Poole, an Australian forester in New Guinea, described the zonation of forests there, from the lowlands to the tree line. His classification was subject to a major revision by Peter Grubb and Timothy Whitmore and minor amendments by others (Grubb et al. Citation1963; Grubb Citation1973, Citation1977; Whitmore Citation1984; Grubb and Stevens Citation1985; Ashton Citation2003, Citation2014); (). This classification has proven robust in summarising the major forest zones on wet mountains in the equatorial tropics, and most specifically in the Neotropics and South-East Asia from where the majority of observations have been published ().
In addition to consistent zonation, two other general patterns have been observed. The effect of mass elevation, first observed in the Alps, whereby the elevation of zonation is particularly elevated on higher and large massifs relative to lower and isolated mountains, has been found to be exaggerated on wet equatorial mountains, and clearly not caused by differences in the temperature lapse rate with elevation (Whitmore Citation1984; Ashton Citation2014). There is also a descent below the average elevation range of temperate herbs, following cold nocturnal valley downdrafts (van Steenis Citation1972a, Citation1972b, Citation1984)
All these studies had been based on observations on single mountains or massifs within a region. In 2003, I published a preliminary review of the extent to which this proven classification of zonation applies in the Eastern Himalaya of Bhutan, at 27°N, on mountains at the northern ecological margin of the Asian tropics, by comparison with that on a near-equatorial high mountain, Kinabalu, 6°03ʹ N. (I define the tropical-warm temperate ecotone as the plant geographic line where exclusively tropical taxa, including the pantropical families Dipterocarpaceae, Myristicaceae and Lecythidaceae, reach their limits and are replaced, over quite a short distance, by exclusively temperate taxa. There is a massive turnover of species in Asia within those many genera which cross the line. Many of the same species continue along each side of this line: from the western end of the wet tropics in eastern Nepal, 88°E, where the line occurs at c.2000 m thanks to shielding from cold winter north-easterlies by the east-west trending Himalaya, to the southernmost lands of Yunnan and Guangxi, 23°N, 108°E, where it lies at <1600 m.). My review was based on personal observation, fortified by details of the elevation range of all recorded tree species, documented for Bhutan by Grierson et al. (Citation1983-2001) and for Kinabalu, by Beaman and others (Citation1998, Beaman et al. Citation2001, Citation2004) and the transects of Kitayama (Citation1992; Aiba and Kitayama (Citation1999). Other mountains were reviewed, and comparisons have been extended (Ashton Citation2003). I later presented evidence, from limited sites, that zonation changes in important respects both with seasonal variation and the declining mean minimum in winter temperature with increasing latitude, and with increasing rainfall seasonality within the continent (Ashton Citation2014).
Here further, albeit more summary accounts of forest zonation on Asia’s tropical mountains are used to present, for one tropical region, the pattern of forest elevation zonation as it varies across the region, to test previous observations, and to further hypothesise what physical environmental or historic factors might be the cause. I am unaware of comparable reviews for other regions. My accounts are based substantially on my own observations, which I lack for Africa and are limited in the Neotropics.
My objective therefore is to present a summary of forest zonation across one of the three main tropical land surfaces that others may determine which patterns are universal, which unique to Asia. The principal questions I address are (1) are the structural and physiognomic criteria for forest elevation zonation, which have held well for wet equatorial mountains, valid for tropical mountains at higher latitudes, and in regions with more seasonal rainfall? (2) To what extent, if at all, are the ecotones between forest zones different beyond the equatorial tropics? (3) To what extent is zonation of climate, soil and forest correlated? And (4) to what extent does zonation of forest structure and physiognomy reflect any zonation in floristic composition?
Singularities of tropical Asia
Tropical Asia differs from other tropical continents in respects that importantly influence the structure, composition and distribution of its montane forests:
Whereas South America and Africa represent unitary continental plates, tropical Asia, although currently overwhelmingly north of the equator, has its origins as a series of separate fragments which have broken off the southern megacontinent, Gondwana, at different times since the Permian epoch. Those several fragments that now comprise the Himalaya and Tibet, and all Indo-Burmese lands south-east to Wallace’s Line, that is to Borneo and Bali, arrived before the origin of flowering plants, leaving no evident biogeographic imprint.
South Asia. India arrived c.55 Mya, from a warm temperate Gondwana but picking up tropical flora as it slid north-north-east past Madagascar and East Africa. A wet tropical flora then migrated from India eastwards to the western Pacific coast, and south-east through the Sunda lands, only partially halted at Wallace’s Line as it came to dominate the lowland forests east to New Guinea and beyond, also those of the Philippines, an oceanic island arc (Whitmore Citation1988). A first migration in the Eocene and early Oligocene, of perhumid climate elements, had to cross the nascent Bay of Bengal. This was followed by an overland transit at higher latitude and therefore seasonal wet climate, which peaked in the mid-Miocene. So complete has been this lowland migration that the signs of an Indian Gondwanan origin are hardly evident, but for the marooning there of two basal Dipterocarpoid genera (Ashton Citation2014). But there is no montane element of clear South Asian Gondwanan origin, other than montane species of predominantly lowland genera in East Asia, most of which are lower montane. A single indisputable Gondwanan taxon, mainly montane or of whitewater upland river banks, exists in the everwet south-west of Sri Lanka: Hortonia, with two to three species, comprising the ancient subfamily Hortonioideae of the southern family Monimiaceae (Renner et al. Citation2010). (Nageia, Podocarpaceae, with one rare species in the Western Ghats and others in the lowland and hill forests of the Far East, may be another). Instead, in South Asia (which I will use here to abbreviate “peninsular India and Sri Lanka”) there are striking absences. These include the northern family Fagaceae which dominate lower montane forests in seasonal wet tropical East Asia, and remain abundant although not dominant in lower montane forests in the perhumid Far East from Peninsular Malaysia to New Guinea; and associated but seldom dominant East Asian and Sunda lower montane genera of northern affinity including Daphniphyllum (Daphniphyllaceae), Engelhardia (Juglandaceae), Altingia and other Hamamelidaceae, Nyssa (Nyssaceae), Anneslea, Schima (Theaceae), Eriobotrya and Photinia (Rosaceae), Acer (Aceraceae), or Ulmus (Ulmaceae), and Betula, Carpinus and Alnus (Betulaceae)each with one tropical montane species. Symplocos (Symplocaceae), Eurya (Theaceae), Bischofia (Euphorbiaceae) and two Michelia (Magnoliaceae) species are the noteworthy exceptions which have penetrated South Asia. Also absent are Australasian elements which variously migrated west of Wallace’s Line: Podocarpus, Dacrydium and all Podocarpaceae except Nageia. These absences obscure floristic boundaries, between lowland and lower montane and lower and upper montane forests, which may nevertheless still be defined by change in species’ dominance as well as structure and physiognomy. It is noteworthy that all but Betulaceae, Corylopsis (Hamamelidaceae) Photinia and Nyssa occur or are occasionally recorded in East Asian lowland forest, though Altingia and Exbucklandia (Hamamelidaceae) not below 500 m. The Gangetic Plain, successor to the Tethys Sea, appears to have been a lowland trough since its origin, further constraining migration since the end of the Eocene epoch by its rainfall seasonality (Ashton Citation2014).
All other Australasian elements are absent from South Asian mountains too, but for Syzygium (Myrtaceae), ubiquitous in Asian evergreen and semi-evergreen forests. Instead, families of putative Indian Gondwanan origin dominate forest zones along elevation. Most notable is the dominance of Dipterocarpaceae in the lower montane forests of everwet Sri Lanka, where the emergent Shorea gardneri, of the Sri Lankan endemic section Doona, was once monodominant in forests now depleted by tea cultivation, while the endemic Sri Lankan basal Dipterocarpoid Stemonoporus, wingless fruited andTrigona-pollinated, has diversified into a host of narrowly allopatric lower montane subcanopy entities.
The Far East. A distinct feature of evergreen forests in the Far Eastern tropics is the dominance, in the different elevation zones, and humult versus udult soils, of families of different continental origin. It is these families which give the forest elevation zones their floristic distinction.
Taxa of Laurasian origin predominate in lower montane forests, warm temperate but for Bischofia which is exclusively tropical. Fagaceae as a family form a characteristic element in the canopy east to New Guinea the Lesser Sunda Islands excepted; Lithocarpus and Castanopsis (Fagaceae),Engelhardia and perhaps Rhododendron subgenus Schistanthe (“Vireya”) (but see Brown et al. Citation2006), have reached New Guinea, Acer Sulawesi and Timor, Ulmus Sulawesi and the Lesser Sunda Islands, Daphniphyllum Sulawesi, and Sycopsis(Hamamelidaceae) Flores: Wallace’s Line, that is the marine Straits of Makassar, serve as a lesser barrier than the terrestrial Gangetic Plain—unless the Laurasian taxa confined in the east to Sulawesi arrived there across the 35-km wide Bali-Lombok channel where few currently extend.
The Australasian floristic element predominates in upper montane forest, in mixture with upper montane elements derived from the general lowland rain forest flora. This Australasian flora is more closely associated with acid humult soils than with elevation, many descending in heath forest (kerangas) to the lowlands in Borneo. No species are confined to upper montane forest as strictly defined, although the Australasian also Neotropical Weinmannia (Cunoniaceae) occurs only sporadically at lower altitudes in lower montane heath forest, and once in a Borneo peat swamp. Tasmannia piperita (Winteraceae), whose stomatal plugs are regarded as an adaptation to prevent inundation in fog, occurs below upper montane forest elevations in New Guinea, where it has diversified into distinct forms, including some in lower montane habitats which whether foggy is undocumented (Vink Citation1970) . In addition though, a number of species, of various geographical affinity are confined to humult soils over ultramafic substrates on Mt. Kinabalu, particularly at upper montane altitudes but also descending below on them (Aiba and Kitayama Citation1999).
Australasia collided with Laurasia c.16 Mya, long after the tropical montane migration route west to South Asia had been broken by India’s relentless trajectory north-north- eastwards, beyond the wet tropics. Of those taxa that have crossed Wallace’s Line westward into Sunda land (Podocarpaceae, Tasmannia, Ascarina (Chloranthaceae), Weinmannia, Scaevola (Goodeniaceae), Styphelia and Trochocarpa (Epacridceae), Kelleria (Thymeliaceae), Xanthomyrtus, Leptospermum, Seorsia and Syzygium (Myrtaceae)), and possibly Rhododendron section Schistanthe (Brown et al. Citation2006) only Podocarpus, Dacrydium, Dacrycarpus (Podocarpaceae), Agathis (Araucariaceae), Styphelia, Syzygium and Leptospermum occur on mountains west of Borneo. Others are apparently restrained by the more limited occurrence of humult soils elsewhere in Sunda land. East of Wallace’s Line, the remaining tropical upper montane forest element of Australasian affinity, the poorly dispersed Nothofagus, like those paradigmatic Australasian vertebrates the marsupials and Rattite birds, is confined to New Guinea.
Culmsee and Leuschner (Citation2013) have shown for the Malesian equatorial region that, on one hand elevational floristic variation dominates over the regional, Wallace’s Line notwithstanding; and on another that phylogenetic diversity increases with elevation. This latter is the reverse of one African example, and may in substantial part be due to the diversity of montane Gymnosperms on many Malesian mountains, and the presence of basal Angiosperm families such as Winteraceae and Chloranthaceae.
The collision of Gondwanan India and Laurasia and its consequences. The contact zone between Indian Gondwana and Laurasia is thought to have remained at current latitudes, 27–32° N, since their collision (R. Hall, pers. comm., 2014), although this view remains controversial. Overall, this collision of two terrestrial continents has nevertheless led to partial subduction of the Indian plate beneath the Laurasian; but the consequent raising of the Tibetan Plateau which had arisen early, and the Himalaya, to their current great heights consequent to continuing uplift of continental rocks by the collision, seems not to have happened before 25 Mya. It was this that initiated the summer low pressure zone over the Tibetan Plateau which, with the overall northern disposition of the hot summer continental mass, drew the southern trade winds north over the Equator and created the south-west Indian wet summer monsoon. It thereby re-extended seasonally wet tropical rainforests westwards into north-east India. A parallel summer wet monsoon occurs east of the Burma-China frontier, where rain-bearing southerly winds reach the east-west Qin Ling Shan at 34° N, and continue up the lands of the western Pacific coast to central eastern Honshu, Japan and beyond. The consequence, a single northern summer-rain and winter-dry climate regime, prevails over nearly all seasonal continental tropical Asia, including all its mountains, providing a simpler correlation between rainfall seasonality and montane vegetation zonation than on other continents. Further, the dry winter weather of the wetter northernmost tropical Asian mountains and adjacent lowlands, from the north-eastern Himalaya to South China, is ameliorated by nocturnal fog and dew (Ashton Citation2014).
The regional lowland forest formations of tropical Asia correlate with the length of the single dry season, that is the number of contiguous months during which expected evapotranspiration exceeds mean precipitation (generally approximated as 100 mm (e.g. Walter and Lieth Citation1960)) but, importantly, not with mean annual precipitation which can vary between 1800 and 5500 mm through the perhumid tropics, yet 2000–13,000 mm through the seasonal tropics, where there at least two dry months.
The latitudinal gradient in temperature is more complex, particularly in South Asia, as semi-arid climates of the plains separate the wet climates of southern, equatorial regions and those of the northern valleys and east-west trending Himalayan massifs, and this affects the climates of those few mountains of the drier regions. Minimum winter temperatures are lowest, not at the latitudinal tropical margin, but in arid regions; these also experience the highest summer temperatures which in addition are most variable consequent on occasional rain storms. Whereas air frost may not occur below 3000 m on the slopes of equatorial mountains, and 2000 m in the eastern Himalaya, freezing air may descend periodically or shortly seasonally dry valleys, both equatorial and eastern Himalayan, to 1500 m, and to 800 m on mountains experiencing dry winters at intermediate latitudes.
The Himalaya draws tropical ecosystems beyond the geographical tropics. Trending East-West, the Himalayas, as the world’s highest ranges, act as a barrier to cold winter north-easterly winds. Frost-free therefore tropical climates and vegetation thereby reach to 32°N in the north-west Indian wet lowlands, and to 27°N in the wet north-eastern hills where the frost line on slopes descends to no lower than 2000 m (Ashton Citation2014).
The proximity of the Pacific Ocean to the east. The Pacific, as the world’s widest and largest ocean, is the motor for a globally influential climatic phenomenon, the Southern Oscillation (ENSO), the roughly five-year oscillation between droughts in its west and floods in its east (El Niño) and their reverse (La Niña). Most intense in regions adjacent to this ocean, El Niño remains strong enough in South Asia to cause the intense droughts that destroy crops and create famine, and to initiate supra-annual flowering in the lowland forest canopy in perhumid south-western Sri Lanka (Ashton Citation2014). Droughts can be intense in equatorial upper montane and subalpine vegetation as well (Kitayama Citation1996).
The north-east trending western Pacific current hugs the coast north to north-east Honshu, at 35.0° N the same latitude as Cape Hatteras on the west coast of the North Atlantic, before moving offshore. But the land continuity of the mild wet climate it causes is continuous from the equator north, and not broken at that critical point—the tropical margin—as it is in the Americas by an incursion of the Caribbean sea. As the earth has continued to spin in the same direction, the Hadley cells, their associated wind systems and therefore many ocean currents are likely to have persisted in their present position since the origin of flowering plants and beyond. Combined with the wet summer East Asian monsoon, this has supported an evergreen forest continuum from the equator north along coastal regions to southern and Central Japan, and inland regions to an east-west mountain barrier, Qin Ling, at 34°N. It has therefore sustained continued opportunities for adaptation of tropical biota into temperate climates, and vice versa. The tropical-temperate vegetational change is therefore of particular interest in East Asia, at all elevations.
The Malesian Archipelago, from Peninsular Malaysia to New Guinea and the Louisiade Archipelago adjacent to the east, is the most extensive region of land masses under an oceanic perhumid (everwet) climate. Mountains in those regions closest to both the ocean and the climatic equator still experience periodic if not continuous orographic cloud and rain during unpredictable lowland droughts.
Departures from the equatorial tropical forest zonation model
Rainfall seasonality and geography. Whereas perhumid climates are dispersed east-west along the equatorial belt, from Sri Lanka to Sunda Land, and on from Sulawesi to New Guinea, the Asian seasonal tropics form a continuum which has the informative presence of two differing south-north trending mountain blocks: the Western and Eastern Ghats of South Asia, and the Far Eastern mountain chains from the Equator north into the temperate Himalaya and South Chinese massifs. Thanks in particular to Stainton’s (Citation1972) extensive description’s of Nepal’s montane forests, ecological studies by Davidar (Davidar et al. Citation2005, Citation2007) in the Western Ghats, and Kanzaki (Kanzaki et al. Citation1999, Citation2004) in northern Thailand, the zonation of forests on those several other mountains in both South and East Asia that have been explored and their floristics described by collectors, can be broadly interpreted, and their leaf size characteristics qualitatively assessed by herbarium examination.
Each of these blocks differs in prevalent rainfall seasonality. Everwet climate is restricted in South Asia to the western faces of the central south-western Lankan massif, which is anyway at ca.7.0° N, that is one degree north of Mt. Kinabalu and the high New Guinea mountains. Mountains with at least seven wet months are restricted to the Western Ghats south of 15°N. The Western Ghats trend further north from there to c.20° N, remaining in the tropics, where they break until the nearest, temperate, north-western Himalaya, at ca.30° N but for the semiarid Aravalli Range which hardly exceed 1000m, and adjacent Mt Abu, at 1722 m. The Western Ghats experience more rainfall-seasonal climates on their eastern than western slopes, enabling close comparison of the influence of seasonality on vegetation. The Eastern Ghats, at c.11-20°30ʹN with the Similipal Range, Odisha, 21° 80ʹ N, isolated to their north, are similarly separated by ca.600 km from the Himalaya, here still wet and tropical, at 26° 50ʹ N, and by a similar distance from the Meghalaya mountains (Khasi Hills) east of the Ganges basin to their north-east. More strongly seasonal overall than the Western Ghats, the Eastern Ghats are wetter on their eastern face, receiving more rain from the weaker, north-east, monsoon over the Bay of Bengal than from the south-westerly.
The mountains of the Far Eastern block under seasonal climate comprise the eastern Himalaya and a series of three north-south trending chains extending southwards from the southern China massif: The westernmost follows close to the India-Myanmar frontier as turbiditic rocks, descending offshore west of the Irrawaddy delta to ascend again first as the Andaman Islands, and then as the Sumatra to Sunda island volcanic “ring of fire”. The second, mainly of granitic rocks, follows the Myanmar-Thailand frontier into the Malaysian Peninsula, terminating in west and central Borneo. The third, as the Annamite Range, granitic with basic volcanic intrusion, runs down the Laos-Vietnam frontier. Between the latter two, low sandstone mountains run south-eastwards from south-east Thailand into coastal Cambodia. The Philippine mountains, mostly influenced by basic volcanic ash and ophiolitic extrusions supporting ultramafic soils, experience increasing rainfall seasonality towards their west. Mountains throughout the Far East receive at least seven months in which average rainfall exceeds evapotranspiration. Most receive this from the summer monsoons, but the Annamite and Philippine mountains receive most from their north-east, winter monsoon. Highest mean annual rainfall throughout tropical Asia is received by mountains fronting the south-west monsoon off the Bay of Bengal, in Meghalaya (Cherrapunji, 1318 m a.s.l., receives 11,000 mm) and along the north-western Myanmar coast (Sandoway, 11 m a.s.l., with 6000 mm), but they still experience up to sevendry months.
Forest zonation on wet equatorial mountains in Asia. Variation in the elevation ranges of forest zones on equatorial mountains has long been documented, but rigorous examination of its cause not fully resolved owing to the continuing reliance on casual field observations in the absence of any elevation transect of meteorological stations. Only recently has it been realised, thanks substantially to the work of Ohsawa (Citation1993, Citation1995)) in the eastern Himalaya, that it is solely the upper zonation of lower montane forest that is affected. The ecotone between lowland evergreen and lower montane forest, albeit gradual, is constantly between ca.750–1300 m throughout both the equatorial zone and higher latitudes, wherever evergreen forests persist across it. It correlates with the cloud base, but in the absence of regular cloud penetration of the forest canopy as fog. It most particularly correlates with an ecotone on clay-rich substrates from tropical lowland yellow-red udult soils with humic discoloration confined to surface horizons, to temperate humic loams in which humic discoloration penetrates to depth and earthworms, manifested as surface casts, become frequent. Importantly, this ecotone has been found to correlate, on these udult loam soils, with the elevation at which rates of litter fall come into balance with rates of decomposition (Dames Citation1955), which itself may be mediated by conditions created by cloud shadow. The ecotone marks the lower limit of temperate year-round vegetable cultivation at low latitudes.
Fog does not penetrate the canopy during the day until higher elevations because the forest canopy retains sufficient latent heat to vaporise fog until air, quickening and cooling as it ascends, in turn cools the canopy to the same temperature as adjacent cloud (Bruijnzeel and Veneklaas Citation1998; Ashton Citation2014). Leaves are wettened by convectional movement of the fog through the canopy; and gas exchange ceases until fog and cloud clears following rain, and intermittently also at subalpine and higher elevations as the cloud breaks up into cumulo-nimbus thunder towers. In the windless climate of the intertropical convergence zone, morning clouds first form orographically, directly above the peaks and following the ridges wherever these exceed ca. 800 m. These generally do not start to accumulate until mid-morning, so that two to three hours of clear sky, and opportunity for free gas exchange from dry canopy leaves and maximum rates of photosynthesis, occur regularly. The clouds thicken, then descend downslope to ca. 850–1300 m, below which the warm lowland air fails to condense until, following afternoon rain, evening fog settles in the cooled valleys.
Cooling and consequent fog penetration of vegetation occurs more quickly on short slopes and on isolated mountains, where the slope updrafts mix earlier with the cool upper atmosphere; and in the cooler saturated sea breezes ascending coastal mountains. The elevation of upper montane forest is lowered thereby. The upper montane forest which adorns the peak of Mt. Santubong, on a coastal peninsula in Sarawak at 1°50ʹ N (location in , is atypical in the thin mossiness of its branches, and the presence of the terrestrial mosses Leucobryum, and Sphagnum bordering a dewpond. These are consistent with observation that diurnal aseasonal summit fog is often driven inland by the onshore breeze. The dense, pervasive bryophyte incrustations characteristic of paradigmatic upper montane “cloud” forest is likewise less evident on the drier, particularly southern slopes of Mt. Kinabalu, 4095 m, at 6° N. Here, the lower montane-upper montane ecotone variously occurs at ca. 1800–2200 m, usually over a short distance. Subalpine elements begin to appear at ca.2900 m, well above, and their increase is gradual. Grubb and Stevens (Citation1985) recorded 40% of woody species to occur in both upper montane forest and subalpine thicket plots at Kerigomna, Papua New Guinea. Occasional killing frost appears to be associated with their presence (Ashton Citation2014). Mossy upper montane forest replaces lower montane forests in everwet Sunda climates on the lower inland Sunda mountains variously at 1500 m (Bukit Retak, Brunei, inland mountains of Sarawak) to 1600–1700 m on Mulu, Sarawak, 1500–1700 m on the Main Peninsular Malaysian range, and 1800–1900 m on the tallest volcanoes of Sumatra and West Java (van Steenis Citation1972a, Citation1972b; Yamada Citation1976). Only Mt. Kerinci, 3805 m, Sumatra is said to experience summit frost away from valleys, but this remains undocumented.
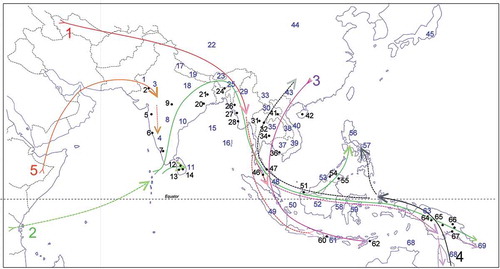
Figure 1. The five immigration tracks of the Tropical Asian flowering plant flora, numbered in large font size: 1, tropical, from the late Cretaceous/Paleocene from North America and Europe; 2, tropical, from the late Cretaceous into Indian Gondwana, and from thence to the Far East from the late Eocene; 3, warm temperate, at first from North America via Europe, but mostly probably from the lower Miocene and East Asia; 4, initially warm temperate, from the lower Miocene, Australasia; 5, tropical arid, from the mid-Miocene, Africa through the middle East. Tracks 2, 3 and 4 make the major contributions to the mountain flora. Place names that are referred to in the text are numbered by in small font size individual localities with black numbering and indicated with a dot, wider areas with blue numbering:
1, Thar Desert; 2, Mt. Abu; 3, Arravalli Range; 4, Western Ghats; 5, Mahableshwar Plateau; 6, Goa; 7, Mt. Nilgiries; 8, Deccan Plateau; 9, Mt. Pachmarhi; 10, Eastern Ghats; 11, Sri Lanka; 12, Mt. Knuckles; 13, Adam’s Peak; 14, Horton Plains; 15, Bengal Bay; 16, Andaman Islands; 17, Himalayas; 18, Ganges Valley; 19, Nepal; 20, Mt. Similipal; 21, Mt. Parasnath; 22, Tibet; 23, Bhutan; 24, Khasi Hills; 25, Meghalaya; 26, Mt. Victoria; 27, Chin Hills; 28, Sandoway; 29, Burma (Myanmar); 30, Indo-Burma; 31, Mt. Doi Chang Dao; 32, Mt. Doi Inthanon; 33, Yunnan; 34, Mt. Kao Yai; 35, Thailand; 36, Mt. Kamchay; 37, Cambodia; 38, Laos; 39, Annamite Range; 40, Vietnam; 41, Mt. Phu Bea; 41, Western Ghats; 42, Hainan; 43, Guangxi; 44, Qin Ling Shan; 45, Honshu; 46, Kangar; 47, Pattani; 48, Peninsular Malaysia; 49, Sumatra; 50, Sundalands; 51, Mt. Santubong; 52, Borneo; 53, Brunei; 54, Mt. Kinabalu; 55, Mt. Silam; 56, Luzon; 57, Philippines; 58, Makassar Strait; 59, Sulawesi; 60, Bali; 61, Lesser Sunda Islands; 62, Timor; 63, New Guinea; 64, Mt. Trikora; 65, Mt. Hagen; 66, Mt. Wilhelm; 67, Mt. Kerigomna; 68, Australasia; 69, Louisiades Archipelago.
On South Asian and Sunda mountains of perhumid climates, fogginess and mossiness leads to increased water supply through physical capture of water vapour, which is associated with soil surface peat (raw humus) accumulation, beneath a blanket of cryptogams or on moss-free animal paths between tussocks (Whitmore, Burnham Citation1984). This may be wet, over a mire-like organic soil, or more often an incipient podsol occasionally developed sufficiently to have a ferric oxide B horizon. Peaty soils are documented in upper montane forest in New Guinea by Mangen (Citation1993), and at upper montane elevations as well as humic clay loams there, by Haantjens et al. (Citation1967) but are not recorded by Grubb and Stevens (Citation1985) or Wade and McVean (Citation1969) from the eastern mountains Wilhelm and Kerigomna (see also Burnham Citation1984). This is discussed in the next section.
The Far East especially Borneo is well endowed with freely draining siliceous substrates, sandstone and granite, supporting strongly leached humult soils, and podsols. They prevail on ridges and spurs of these mountains. The podsols support a lower montane forest similar in structure and physiognomy to lowland heath forest (kerangas), and short and gnarled on skeletal soils in this resembling upper montane forest. I once even observed, in central Sarawak, Borneo, forest with heath forest floristics, on a peaty gley overlaying a basalt plateau, at 1100 m. Further, upper montane forest in sheltered coves, on Mt. Kinabalu above Paka Cave for example, and widely on the great New Guinea massifs, reach the stature of lowland heath forest, though boles remain more crooked. The lower montane heath forest flora is predominantly that in the lowlands, with a lesser component unique to lower montane elevation, and an increasing representation of species concentrated in upper montane forest at higher elevations as intermittent penetration of fog increases (Ashton Citation2014). Forest with the structure and composition of upper montane forest, but whether as mossy has not been documented, often including local endemics, descends to exceptionally lower elevations on the humult soils over ultramafic substrates (Proctor et al. Citation1988, Citation1989; Aiba and Kitayama Citation1999; Aiba et al. Citation2005, Citation2006).
This floristic correlation with raw humus-bearing soils results in the extension of the upper montane tree flora downslope into lower montane forest on low-nutrient freely draining substrates, especially sandstone but also granite, where the structural and physiognomic characteristics of upper montane forest are reduced or lost. Such soils appear not to occur on wet South Asian mountains on slopes where penetration of fog is limited to a short season. But the Indo-Burmese sandstone mountains Khao Yai, Thailand and Kamchay, coastal Cambodia, with summits barely exceeding 1000 m and a short, two month, dry season, are clothed with such lower montane heath forest over humult soils, in which a canopy of the dominant podocarps Dacrydium elatum and Dacrycarpus imbricatus shelters lower montane elements (Smitinand Citation1968). Kerr reported vegetation resembling heath forest, bearing an abundant small deciduous tree resembling Vaccinium overlying a sandstone exposure, at lower montane altitudes on Phou Bea, in the northern Annamite Range at 19°25ʹ N.
I therefore still subscribe to the existing definition for tropical upper montane forest of Whitmore (Citation1984) and Bruijnzeel and Veneklaas (Citation1998). It differs from lower montane heath forest, and structurally and physiognomically similar forest on ultramafic substrates at lower altitudes, in its continuous cover of moss and other cryptogams, on soil as well as branches where it forms thick sleeves resembling plumbing insulation cladding. Although generally short-trunked and more gnarled than heath forest, it can frequently reach 40 m, and reach 20 m if irregular trunked, over extensive slopes on the taller mountains of New Guinea. The woody species concentrated in upper montane forest bear leaves predominantly smaller than the notophylls dominant in lowland and lower montane heath forest, though smaller size classes also appear to increase with altitude in lower montane forests. Rather than those very few tree species usually confined to upper montane forest, it is defined by its distinct cryptogam, and a distinct vascular epiphytic flora albeit less rich than the lower montane. Its distribution is strictly confined to regions with most continuous diurnal fog penetration. With this definition, tropical upper montane forests are confined to equatorial perhumid climates, but also extend to the margin of the tropics on everwet oceanic islands, as on Kauai, Hawaii (Ashton Citation2014). Here, and on coastal mountains such as Santubong, Sarawak, and Silam, in Sabah, it is the descent of diurnal fog penetration which correlates with the descent of upper montane forest. This does not lead to a contraction of forest zonation overall, but to truncation of the upper range of lower montane forest which, as on Santubong, may be entirely missing.
I define subalpine forest or thicket, and shrublands more by predominance of woody plant leaf sizes smaller than notophyll, and shiny leaves many of which are concave – spoon-shaped with concave surface uppermost (Mangen Citation1993; Ashton Citation2014; . Dense twigginess appears to be more apparent at its elevation and as leaf sizes and shapes change in both South Asia and the Sunda mountains, than in most upper montane forest. Mangen (Citation1993); records these leaf and crown characteristics from Mount Trikora, western New Guinea, though neither Wade and McVean (Citation1969); nor Grubb and Stevens (Citation1985); note it in the eastern New Guinea mountains. Cryptogams and especially vascular epiphytes are much reduced, vines absent. The zone is associated with skeletal soils with patchy moss cover, and mires. It experiences less consistent fog than upper montane forest, and periodic intense drought (Kitayama Citation1996), although Mangen observed subalpine thickets on Trikora to be as densely tussock-mossy as upper montane forest. Although observations are as yet sporadic, subalpine thickets appear to correlate with the lower limit of ground frost on slopes and ridges. The ecotone between upper montane forest and subalpine thicket, structurally defined, is often narrow, but the floristic transition is gradual (Beaman and Beaman Citation1998; Beaman et al. Citation2001, Citation2004).
The tree line in the equatorial tropics. The tree line in the equatorial tropics, defined as the elevation limit of the tree life form either as stands or individuals, has been observed in Asia at 3650–3900 (4200) m in New Guinea, 3400 m on Mt. Kinabalu (Körner Citation2012; Hope Citation1976, Citation1980; Mangen Citation1993; & pers. obs.). This is below the frost line, coincides with the limits of cultivation in New Guinea (Nagy and Grabherr Citation2009), and appears to approximate with a global growing season temperature of ca.6°C (Kudo Citation2001; Körner Citation2012). On wet continental mountains of monsoon Asia there is an ecotone, generally over a narrow elevation, from floristically characteristic tropical lower montane to warm temperate forests, from Mt. Victoria, Burma, 21°12ʹN (Kingdon Ward Citation1958) north to the eastern Himalaya and northern Burma at ca.27°N (Ashton Citation2014 and below). This ecotone descends to the lowlands with little floristic change in southernmost China, at ca.23°N. Whereas there is some familial and generic but predominantly species turnover at this northern tropical margin, species that persist as trees on equatorial mountains, such as Leptospermum recurvum, Schima brevifolia, and Tasmannia piperita on Mt. Kinabalu, all of which occur there on exposed outcrops in upper montane forest as low as 1800 m, extend into alpine elevations as low shrubs in granite fissures to the summit plateau, at 4000 m (Beaman et al. Citation2001, Citation2004). Subalpine tree species likewise extend in New Guinea (Mangen Citation1993).
But whereas soil becomes confined to hollows and fissures in the steep granite slopes of alpine Mt. Kinabalu, and woody vegetation becomes scattered and increasingly sparse, soils of varying depth and bogginess continue to the highest elevations in New Guinea. There, subalpine vegetation below 3750 m forms a mosaic of tree thickets, tall and dwarf shrublands, and grasslands some of which anthropocentric owing to periodic fire and many as Cyathea tree fern savannas; but grasslands extend well into alpine elevations there. A characteristic alpine woody flora, absent on Mt. Kinabalu, also appears, at first mixed with subalpine elements. Nanophyll heathlands, −1 m tall, dense or imbedded in grassland, have been described from Mt. Albert Edward at 3450–2750 m, and more clearly at sabalpine elevations, 4230–4400 m, on recent moraine on Mt. Wilhelm (Hope Citation1980).
Tree mortality at subalpine elevations. Tall individuals of L. recurvum have suffered periodic scattered mortality on Mt. Kinabalu, apparently due to occasional periods of exceptional drought, and perhaps accompanying air frost (Aiba and Kitayama, Citation2002; Ashton Citation2014). Potentially explanatory observations are informative on the Horton Plains, Sri Lanka, 1900–2100 m, and the Western Ghats summit ridges, where frosts frequently occur during winter droughts. The presence of strips across the north-east facing scarp edge of the Plains, of complete forest mortality may be attributable to drought or frost brought by periodic droughts during the winter north-east monsoon. Anthropogenic fire-induced fires result in widespread grasslands in swales and on slopes to the summits. Although woody temperate species occur (Ashton Citation2014, see also below), upper montane woody elements are absent but regeneration has been observed in dense, Sinoarundinaria bamboo and Hedyotis (Rubiaceae) thickets adjacent to forest margins, recalling similar observations at high altitudes in New Guinea (Gillison Citation1970). Further north, in the Nilgiri mountains of the Western Ghats, higher grassland slopes became invaded and covered by the Australian open-crowned and feathery-foliaged tea estate shade tree Acacia mearnsii. This species suffered total mortality In the cold and frosty 2007–8 winter, whereas adjacent upper montane forest, with dense canopy foliage, was unaffected, even in valley bottoms (Ashton Citation2014). The forest canopy appeared to resist frosting on account of the latent heat conserved within, thereby allowing survival of regeneration.
Wind. Important, but unsystematically documented, is the influence of wind on montane forest canopy structure and physiognomy. Periodic strong winds have been experienced at upper montane and submontane elevations on Mt. Kinabalu, and occur annually with the wet summer monsoon in the seasonal tropics (Ashton Citation2014). Probably more universally important is the observed increase in updraft wind speeds with elevation. It is generally observed that lower montane forests decrease in stature, and their canopy becomes more densely even with elevation, often manifesting crown shyness (Ashton Citation2014). Upper montane forests along exposed and higher ridges are exposed to the full force of prevailing winds. Their stature may also be attributed to shallow soils and greater proneness to water stress, but variation in soils depth along ridges would be expected to be manifested in greater variation in canopy stature along short distances than is observed, as is conspicuously seen in heath forest at lower elevations. Twig expansion is being completed as the wet summer monsoon winds arrive. A conspicuous exception to customary structural change with altitude occurs on the south-west slopes of Adam’sPeak, 2243 m, 6° 49ʹN, and adjacent mountains at the western face of the Peak Sanctuary, south-west Sri Lanka, where the forest receives the full force of the southwesterly monsoon. Forest with the stature and structure of upper montane forest here descends to c.1300 m. The southwest face of the Knuckles Range to its north, and elsewhere may have mainifested similar forest, but are mostly too disturbed now to be certain (Greller and Balasubramanyam Citation1988; Greller et al. Citation1987). The tree flora, though, is lower montane, as are predominant leaf sizes. These provide further evidence that wind, as much as fog or alternating fog and sun, is a major influence on upper montane structure, but not tree species composition or leaf sizes although some lower montane canopy species, notably Shorea gardneri, are absent.
In summary, the elevation of forest zones on equatorial and wet coastal tropical mountains in Asia, and elsewhere where documented, conform in their elevation with the global vegetational and ecological belts presented by Körner et al. (Citation2011) (though differing in terminology) except in the case of upper montane forests, whose elevation and very presence is determined by the elevation at which cloud fog penetrates the canopy, and its seasonal consistency.
Tropical montane forest change with increasing seasonality
Lower montane forests and their lowland ecotone. The forests of rainfall-seasonal tropical continental Asia have been described and classified by Champion (Citation1936), and reviewed by Ashton (Citation2014). Evergreen lower montane forests, termed hill evergreen forests by Champion and subsequent foresters, replace lowland forests which differ there from lower montane forests in their higher proportion of deciduous species, which varies in accordance with the number of dry months and their proneness to fire.
Eastern continental Asia. The Kangar-Pattani plant geographic line on the Malaysian Thai frontier marks the floristic and phenological change from lowland mixed dipterocarp forest to southern seasonal evergreen dipterocarp forest, and approximately correlates with the northern boundary of climates with less than two dry months (Whitmore Citation1984; Ashton Citation2014; . Lower montane forests remain distinguished by a characteristic flora at genus and higher level, in which the canopy is dominated by Fagaceae, north to the tropical margin where they reach their limits. There, the temperate evergreen forests that are above them in the eastern Himalaya, and which are similar in genera but different in species, descend to the lowlands in warmest temperate China (Citation2003). However, there is a substantial change at the Kangar-Pattani Line at species level, notably among canopy Fagaceae whose canopy dominance increases to its north. The ecotone between lowland mixed dipterocarp forest of everwet climates, both southern and northern lowland seasonal evergreen forests, and lower montane forests remains at 750–1300 m throughout the Far East (Ashton Citation2003). In the north, especially where cool season nocturnal fog descends slopes to low altitudes, several lower montane species of humic soils including Schima wallichii, Bischofia javanica, Engelhardia roxburghii and Castanopsis indica descend to the foothills; but the overall composition of these foothill forests remains that of the lowland flora.
In the most seasonal climates abutting Far Eastern high mountains at middle latitudes, experiencing five to seven dry months and sheltered from wet winds off the oceans, periodic fire in the lowlands, mostly anthropogenic, leads to predominance of moist deciduous forests (Ashton Citation2014). This is associated with a relatively narrow ecotone from lowland to lower montane forest as high as 1300 m, but more often at c.1000 m.
The driest slopes, from Meghalaya to the drier mountains of Luzon, Philippines, especially where sheltered from the monsoon, support a savanna woodland with an ecotone at c. 1000 m, where the lowland Pinus merkusii of edaphically dry deciduous dipterocarp forest gives way to a savanna dominated by Pinus kesiya. On the continent, P. kesiya is associated with a distinct albeit impoverished woody flora, partially shared with deciduous dipterocarp forest. This includes Ternstroemia gymnanthera (which extends to warm temperate Japan), semi-evergreen Castanopsis argyrophylla, deciduous Quercus kerrii, and evergreen Craibiodendron stellatum and Vaccinium sprengelii in the understory.
South Asia. Sri Lanka’s seasonal mountain slopes once again differ from those of the Indian peninsula. Lowland eastern and northern Sri Lanka, which experiences both south-westerly and north-easterly wet monsoons and two dry seasons (merging in the north-east) in which monthly rainfall is less than 100 mm (Walter et al. Citation1975), lack deciduous forest or much of its woody flora, while a semi-evergreen forest, occupying a narrow band between mixed dipterocarp forests and semi-evergreen notophyll woodlands, is floristically impoverished. What little remains of forest at lower montane elevations on these dry slopes is short, much branched, and with its own distinct flora.
The Western Ghats provide the best understanding of the fate of the lowland to lower montane forest ecotone as rainfall seasonality increases, through their increase in rainfall seasonality both from south to north, and from western to eastern slopes. Their montane forests have been subject to a monograph, based on extensive field observation, by Blasco (Citation1971). In the absence of the Fagaceae dominant in Far Eastern lower montane forests, and the dominant Shorea gardneri and rich woody flora confined to lower montane forest in everwet south-west Sri Lanka, the lowland seasonal evergreen forest-lower montane forest ecotone on the western slopes of the Western Ghats is more subtly distinguished. The dipterocarps die out by 1300 m (Vateria indica; Dipterocarpus indicus and Hopea parviflora to 1200 m).Towards the south, in a climate with up to four dry months, the ecotone is marked by a zone in which lower montane specialists Palaquium ellipticum, Cullenia exarillata and Mesua ferrea, together or singly, become subdominant. Towards the somewhat more seasonal north, and on basaltic trap-derived red-brown soils, Poiciloneuron indicum is locally dominant. Above each are mixed evergreen lower montane forest communities which change in composition with elevation. Mostly comprised of lowland genera, with both lowland and lower montane specialists, a few tree taxa are confined to these forests, including the widespread principally lower montane forest Bischofia javanica, Meliosma species (Sabiaceae), Aglaia roxburghiana and Garcinia echinocarpa. Lauraceae and Clusiaceae are more strongly represented here than in either lowland or upper montane forests.
Peaks hardly exceed 1000 m north of the Nilgiries. North of ca.14 N, along the western face of the Western Ghats where southern lowland seasonal evergreen forests give way to semi-evergreen forests, the ecotone to lower montane forest becomes obscure. On the drier eastern slopes, semi-evergreen forests hug the foothills, here descending to the Deccan Plateau at >600 m, where some still survive. Here, and north of Goa on the western slopes with more than seven dry months, the influence of fire becomes increasingly pervasive, and lowland evergreen is replaced by deciduous forests. Again there is no longer a clear lower montane ecotone, at least partially owing to centuries of human disturbance. Blasco implies that, although teak (Tectona grandis) does not occur above 1100 m, and the short deciduous forest bamboo Dendrocalamus strictus above1500 m, many deciduous forest taxa including Anogeissus latifolia, Pterocarpus marsupium, Terminalia tomentosa, T. chebula, Acacia spp., Buchanania lanzan, and Bridelia retusa extend higher. The higher Western Ghats end at the Mahabeleshwar Plateau, c.17° 60ʹ N, 1000–1470 m, with 6330 mm mean annual rainfall concentrated over four wet months. There are still lowland semi-evergreen remnants to 1050 m, and evergreen lower montane forests on western slopes which include Artocarpus heterophyllus, Actinodaphne angustifolia, Cinnamomum macrocarpus, C. zeylanica, Nothopodytes nimmoniana, Litsea stocksii, L. wightiana, Beilschmiedia dalzelii, Myristica dactyloides, Olea dioica, Aglaia lawii, Symplocos beddomei and the vines Clematis wightiana and Jasminum malabaricum; but tall deciduous forest elements, including Emblica officinalis, Canthium dicoccum and Terminalia chebula extend here into lower montane elevations (Deshpande et al. Citation1993). Soils are described as “red”. The summit plateau supports an unique, thinly mossy forest with 5–12 m tall upper montane structure, but notophyll-microphyll woody flora including Memecylon umbellatum and Syzygium species.
North again and at the southern end of the Aravalli Range, Mt. Abu, 24° 41ʹ N, 1722 m, at the edge of the semi-arid zone with thorn woodland in the lowlands, lacks lower montane evergreen forest (Ahluwalia Citation1965; Jain Citation1968). However, its 'upper valleys' sustain the evergreen trees Aglaia odoratissima, Syzygium cumini, and Mallotus philippensis, and many deciduous forest species including Anogeissus latifolia, Crateva nurvala, Bauhinia purpurea, Lannea coromandelica, Adhatoda beddomei and Sterculia urens. Mt. Abu is north of the Tropic of Cancer. Although its lower slopes bear short tropical deciduous forests, its upper elevations support 83 angiosperms regarded as temperate, with ranges of 19 extending to the Himalaya, 38 on the Western Ghat summits, and 26 more widespread (Jain Citation1968). Neither the upper limits of tropical species, nor the lowest elevation of frost, seem to have been recorded, but from personal observation on the highest Eastern Ghats appears to be near 1000 m. To its north is the Desert of Thar, with annual frost into the lowlands north of the Aravalli range which hardly exceed 1000 m. North again and west to 76° N in the Himalayan foothills, tropical deciduous S. robusta forest reaches 32° N. Once opened by felling, the Shorea juveniles are subject to killing by frost. Tropical mountains with upper montane elevations, that is above 1800 m, north of 12°N are therefore confined in Asia to the Far East.
The Eastern Ghats are bordered by short deciduous forests to their west, and semi-evergreen and tall deciduous forests to their east. Few hills exceed 1000 m. Their lower montane forest component is reduced to a few elements. At Similipahar, Odisha (21°38ʹN, 86°26ʹE), they include Meliosma pinnata and some Lauraceae – Actinodaphne angustifolia, Neocinnamomum caudatum, Phoebe lanceolata, mixed with lowland evergreen elements: Alstonia scholaris, Mangifera indica(Saxena and Brahmam Citation1989). A short-statured upper hill form of tall deciduous forest prevails to the Meghasani summit, 1166 m, with Shorea robusta dominant except on the summit rock, and in the frost prone valley bottoms where an extraordinary low thicket of shoots, arising from persisting S. robusta lignotubers, set in a sward of festucoid grasses with temperate herbs, Corydalis, Hypericum, Viola, are periodically cut to the ground by frost, (plate 4.15g, Ashton Citation2014).
In summary, I hypothesise, from the evidence overall, that the motor to the gradual lowland-lower montane forest ecotone is the cloud base which, by bringing diurnal shadow, accelerates reduction of diurnal temperatures and increases humidity, especially beneath the canopy. But the direct cause appears to be a soil ecotone on loams, from lowland yellow-red soils in which organic matter discoloration is restricted to surface horizons, to montane soils similar to lowland temperate mulls in which humus discolors the profile to depth. This is the elevation at which rates of litterfall in evergreen forests is said to come into balance on tropical loams (Dames Citation1955), and at which dominance of termites as fine litter decomposers is replaced by that of earthworms (Collins Citation1980; T. Abe, pers. comm,.). In regions where the rain season and cloud shadow is relatively brief, forests become susceptible to fire to higher altitude, even to the frost line, thereby eliminating lower montane forest except in moist shady valleys. Soils remain of the lowland type. I infer that the deciduous forests at lower montane elevations in South Asia have failed to evolve a distinct woody flora, perhaps because they are too young, alternatively because the prevailing lowland deciduous flora remains competitive to much higher altitudes in the absence of diurnal cloud and, specifically, a major change in soils.
Upper montane and subalpine forests. On the undulating Horton Plains, 1900–2400 m, Sri Lanka, at 8.0° N, frost is of frequent occurrence in the swales and apparently more occasionally elsewhere, particularly on crests facing the drier eastern scarp, a cliff, described above. Here, low stature forest with the overall structure of upper montane forest is ubiquitous. Festoons of the lichen Usnea imply periodic drought (e.g. Smith Citation1995). Canopy crowns are markedly twiggy, but mossiness more patchy than on the Sunda mountains. Some canopy tree species, including Syzygium revolutum, S. orbiculatum (Myrtaceae), Alseodaphne speciosa, A. ambigua, and Litsea ovalifolia(Lauraceae), do bear obtuse or subacuminate cupped or revolute leaves with glossy upper surfaces (), but they are notophyll or microphyll. Indeed, few species with leaf sizes smaller than nanophyll as would be expected in upper montane or subalpine vegetation are represented. Further, Lauraceae are represented by a rich and more abundant flora than in Sunda upper montane forests.
The Western Ghat higher crests above c.1900 m are well endowed with a distinct tree flora with upper montane leaf sizes, and the lower montane flora is reduced to a few persisting species. Mossiness is relatively sparse here and Usnea is again a frequent presence. The lower to upper montane forest ecotone is less clear than on the Horton Plains. On the northernmost mountains of upper montane elevation, the Nilgiries, c. 11° 30ʹ N, forests with the stature and leaf physiognomy of the higher occurrences of lower montane forests in the Far East occur to at least 2200 m on east-facing slopes, while adjacent west facing slopes bear forest with the structure and physiognomy of upper montane forest (Ashton Citation2014 p.282, table 4.6b). Some of the field layers of these lower montane upper woodlands are rich in Zingiberaceae. Whereas the western face is bathed in cloud from the south-west monsoon and orographic upwelling for at least nine months, the eastern is sheltered from the monsoon and lacks moist orographic upwelling, instead receiving intermittent cloud during the five-month north-east monsoon. Lower montane humic mull soils here similarly extend to summit ridges on east facing slopes, while poorly developed leached humult podsolic soils cover the upper west facing slopes.
The Far East. Upper montane forests and subalpine thickets are confined to equatorial mountains, where maximum and minimum monthly temperatures differ little and frosts are occasional, aseasonal and supra-annual. But wet Far Eastern mountains north of c.20°N latitude experience increasing differences in seasonal mean monthly temperature, and the occurrence of increasingly regular winter, that is dry monsoon, frosts above 2000 m. Doi Inthanon, 18° 60ʹ N, 2320 m, the highest mountain in Thailand, is clothed with lower montane forest dominated by Fagaceae to its summit, where it bears a dense understory of Acanthaceae and Zingiberaceae. Weakly frost-hardy Rhododendron arboreum ssp. delaveyi occurs on open ground, while an adjacent high valley bottom bears a grassland fringed with the warm temperate Lyonia ovalifolia. This summit vegetation confirms the trend observed in the Nilgiries, that lower montane forest extends to higher elevations once fog declines to a certain seasonal frequency: persistent diurnal fog is the factor which limits the upper elevational range of lower montane forest, not temperature, nor competition, nor perhaps soil.
The mountain backbone of New Guinea is by far the largest mountain system in equatorial Asia. Its latitudinal spread contrasts with the north-south trend of both the Neotropical cordillera and African high tropical mountains. Whereas the description of soils and forest zonation on the upper slopes of Mt. Trikora by Mangen (!993), western New Guinea, parallel those on Mt. Kinabalu, Grubb and Stevens (Citation1985), on Mt. Kerigomna, eastern New Guinea, documented humic mull soils in upper montane forest, and at upper montane forest altitudes, more resembling those of Doi Inthanon or east facing Nilgiri slopes, and Wade and McVean (Citation1969) appear to concur for the eastern Mt. Wilhelm. The thorough soil surveys of Papua (eastern) New Guinea (Haantjens et al. Citation1967) have indicated, though, that “peat” soils are indeed widespread between 2000–4000 m on Mt. Hagen and elsewhere in the east, but are also often replaced by lower montane humic brown clay loams, consistent with Grubb and Stevens (Citation1985). Haantjens’ group variously attribute these to basic volcanic substrates or ash, or to steep topography. Whitmore and Burnhm (Citation1969) and Burnham (Citation1984) described the presence of humic red-brown mull soils at upper montane sites in the seasonal wet tropics of the Far East, but implies that peat bearing soils are pervasive in everwet regions. Grubb and Stevens (Citation1985), instead, noted that the diurnal cloud base in the central New Guinea massif ascends to 2800 m, and forest with lower montane characteristics extend to 2850 m, albeit with increasing upper montane woody elements, but also observed only thin cover by moss and other cryptogams, either on ground or bark. They failed to record characteristic floristic elements of upper montane podsolic soils, notably Syzygium and Nothofagus. I am sceptical that paradigmatic thickly mossy equatorial upper montane forest, dripping diurnally with fog-captured precipitation, can long avoid accumulation of surface peat. The meteorological data of Grubb and Stevens imply that it is also possible that the vast massif of eastern New Guinea possesses rain shadow slopes and valleys, where short annual dry seasons occur comparable to the upper seasonally wet eastern slopes of the Ghats. At higher New Guinea elevations, where structure and physiognomy, and increasingly floristics, gradually come to conform with that of upper rather than lower montane forests and eventually subalpine thicket, stand structure may be primarily influenced, and become limited, by temperatures as mean rates of respiration approach mean rates of photosynthesis (Körner Citation1998), even in the absence of persistent aseasonal diurnal fog penetration. A similar primary influence of temperature may be observed as the tree limits are approached, and in other high tropical massifs notably the Andes.
At 18°58ʹ N, Phou Bea, 2819 m, highest mountain in the Annamite chain, is surrounded by northern seasonal evergreen forest but for moist deciduous and deciduous dipterocarp forest to its south-west. Its shale summit, unique among mountains at this latitude, bears a stunted thickly mossy woodland with the physiognomy of upper montane forest (Kerr Citation1933). Snow has been observed on the summit, and I conclude that its vegetation represents a warm temperate community. At 3063 m, Mt. Victoria, 21° 12ʹ, at the northern end of the Arakan Yoma where they meet the Chin Hills in western Myanmar, is by far the tallest peak in the seasonal Far Eastern tropics south of the Himalaya. Kingdon Ward (Citation1958) observed lowland semi-evergreen forest to 1000 m, and from there a lower montane forest association to 2000 m, which also marked the limit of shifting cultivation. The frost-hardy Rhododendron arboreum was noted at this elevation. Ward described the forest above that elevation as temperate semi-evergreen rain forest, observing Carpinus, Betula, Alnus and Acer campbellii here for the first time. He noted Symplocos, Pyrus, Michelia doltsopa, Eriobotrya, Cornus capitata, Prunus cerasoides, Ilex, then Taxus (the only temperate conifer, but extending to seasonally fog-free lower montane forest at c.2000 m in Sumatra (Ohsawa et al. Citation1985; Spjut Citation2007) and Luzon (pers. obs. 1975), Symplocos, Quercus xylocarpa and other Fagaceae, Malus, Photinia, and Viburnum further up, and continuing presence of Lauraceae; but no Hamamelidaceae, or other exclusively temperate East Asian tree families. At highest elevations, Ward described the stunted forest as dominated by Quercus semicarpifolia which, if correct, would indicate seasonally dry warm temperate forest. Surprisingly, he does not document his principal interest: a rich temperate herbaceous flora.
Open ground approaching the summits of many higher mountains of seasonal Asia support temperate shrubs, grass genera, and herbs. The repertoire is limited on Horton Plains at 8° N, and along the Western Ghats where there occurs a Gaultheria, a few Vaccinium, Berberis asiatica, and local subspecies of Rhododendron arboreum. Parasnath, 23° 57ʹN, in central India south of the Ganges, 1350 m, supports a rich temperate herbaceous flora, while B. asiatica occurs on Pachmarhi, 1350 m, 22° 29ʹ N. In the Far East, this temperate element does not extend south of 16° N. High limestone karst is particularly rich in temperate climate herbs, documented for Doi Chiangdao, Thailand, 2175 m, 19° 22ʹ N (Smitinand Citation1966). The presence of these herbs is indicative of ground frost on open ground, but closed canopy forest is clearly immune to frosting to higher elevation. In the eastern Himalaya, valley frost descends, as in New Guinea, to 1500 m, but temperate climate tree species are recorded no lower than 1800 m. Few tropical tree species exceed 2400m in Bhutan, the majority not exceeding 2100 m. In conclusion, by 18° N lower montane forest is again becoming truncated, this time by warm temperate forest.
The summit vegetation of Phou Bia, and the extension of tropical montane genera notably Ardisia, Symplocos, Meliosma, and of course Ilex, with profuse if thin mossiness into Himalayan warm temperate forest, might imply continuity of tropical upper montane forest into the wet warm temperate zone. Many canopy species here also bear cupped leaves, glossy above. But these temperate forests are tall, straight-boled, and predominantly notophyll, in this more resembling tropical lower montane forest. Their species, with few exceptions, are also different, while their cupped glossy leaves are more a characteristic of subalpine than upper montane tropical communities.
Species of exceptional stature. Some species in the Asian tropics albeit restricted by history to certain regions, provide local challenges to montane forest definitions, in all cases in the form of species which exceed usual stature for their zone. Several monopodial gymnosperms (Podocarpaceae, Agathis) become emergent in both lower and upper montane forest, and are abundant, though generally scattered on Mt. Kinabalu and in New Guinea (Grubb and Stevens Citation1985; Beaman and Beaman Citation1998). In Sri Lanka it is a short but distinctly emergent and microphyll monodominant dipterocarp, Shorea gardneri, once universal throughout the lower montane forests except in areas subject to the full force of monsoon winds. On Doi Inthanon, northern Thailand, Podocarpus neriifolius and Mastixia euonymoides(Cornaceae) reach 40 m in sheltered lower montane coves, their crowns still trending towards the north-east to the lee of the south-west monsoon. In New Guinea it is the distinct belt of dominant or co-dominant Nothofagus species, which range between the elevations elsewhere described for upper montane forest. Predominantly microphyll or nanophyll, and densely mossy, they are associated with species typical of upper montane forest, although some more typical of lower montane forest are present at their lower ranges (Hynes Citation1974; Soepadmo Citation1976). I interpret them as an upper montane forest in which an Australasian canopy genus of warm temperate climate origin has invaded. More observations are needed to test their correlation with soil, which I observed to be humult. In upper montane and more particularly subalpine forest zones, there are species which exceptionally emerge above an otherwise dense and uniform canopy: Leptospermum recurvum on Mt. Kinabalu, Schefflera altigena and Saurauia alpicola in New Guinea, all of which can reach 8 m, and Calophyllum walkeri on Sri Lanka’s Horton Plains. These species can suffer periodic mass mortality, the cause of which remains conjectural (Ashton Citation2014, and above).
Do structural-physiognomic and floristic ecotones coincide? This question can only be rigorously tested by census of continuous elevational transects; these await implementation. The evidence rests therefore on individual small plots, notably those documented by Aiba and Kitayama (Citation1999, Citation2005, Citation2006) and by Kitayama (Citation1992) on Mt. Kinabalu; and histograms of the numbers of tree species present along the elevation gradient, and their lowest and highest records, such as those illustrated by Ashton (Citation2014) in 300 m sectors for Kinabalu and the Bhutan Himalaya, derived respectively from the floras of Beaman and Beaman (Citation1998, Citation2001, Citation2004), and Grierson et al. (Citation1983–2001). Whereas the plots are set in forests defined by structure and physiognomy, they ignore floristic ecotones. Peaks in species presences which would serve as evidence of ecotones are muffled: at the lowland to lower montane ecotone by its elevation spread over 400 m, and at the lower to upper montane ecotone by its variability in elevation, even between peripheral and central peaks and ridges compared with slopes on a single massif. The evidence from records of individual species’ ranges, and personal experience indicates that, although few species are strictly confined to individual forest structurally defined elevation forest zones, every zone is nevertheless distinguished by an array of species and genera which are consistently more abundant within it.
Conclusion
ereas a gradient of continuous floristic change with elevation is described on all mountains visited whether in perhumid or seasonally dry, a consistent sequence of zones, with floristically, structurally, physiognomically, and/or phenologically distinct forest types, separated by distinct ecotones of varying breadth, is also manifest, and is broadly consistent with global bioclimatic belts along elevation (Körner et al. Citation2011), albeit differing in terminology. The influence of rainfall seasonality and decline in minimum winter temperatures on forest characteristics and zonation has only recently been documented (Ashton Citation2014). Whereas rainfall seasonality is mediated by continentality, minimum temperature is primarily correlated with latitude although the elevation of frost inception is also depressed in areas with dry winters. The lowland – lower montane forest ecotone, albeit gradual, remains at the same elevation throughout the Asian tropics. Even where both lowland and lower montane elevations are clothed with deciduous forest some floristic change, either of the disappearance of lowland species or the arrival of species confined to lower montane elevations, implies that a change in physical environment continues to occur in climates where diurnal cloud is strongly seasonal (). What that factor is remains unclear, although it may be edaphic. Persistent diurnal fog determines the very presence of tropical upper montane forest; lack of fog leads to the persistence of lower montane forest to higher elevations, wherever an annual season without fog occurs. Frost at higher latitudes limits the upper limit of all tropical forest types.
Table 2. Summary of elevational zonal changes in climate, soils and forests in the humid tropics. The motor to habitat change with elevation is temperature decline, ca. 6.5 K km-1 ascent on tropical mountains, and its interaction with atmospheric humidity to produce cloud (fog) and consequent precipitation. These factors of climate, with substrate, determine soil formation in interaction with forest cover and litter production. Absence or presence of cloud/fog seasonality results in differing soil and forest zonation, indicated in the paired columns. Temperatures are mean monthly values.
Some (Kira Citation1977; Ohsawa Citation1993; Citation2006) have presented indices, derived from combinations of climatic measures, which identify intervals along whose elevation range, it is claimed, ecotones consistently occur. I have questioned whether that is so (Ashton Citation2014, p. 267), as no mechanistic explanations for such correlations been presented. Rather, it is the floristic gradient along elevation that would appear to be caused by continuously changing climatic factors, notably temperature, acting on the competitive interrelationships between species populations as they each differentially change in their performance in response to change in ambient climate. However the zonal ecotones along this gradient are seen to correlate with change in limiting factors, climatic such as diurnal fog or nocturnal frost, or edaphic which may in part be initiated by a climatic factor such as diurnal cloud shadow, and which are the consequence of interaction between such factors and the tree flora present. These ecotones promote floristic and structural-physiognomic change in the whole forest community; and those changes persist at increasing elevations until another ecotone is reached. They appear to be mediated by differing species-specific responses to differing limiting physical factors, and at different stages of their life history, as can most clearly be seen as frost becomes more frequent: These are topics still awaiting rigorous examination and experimental testing. As an overall result, floristic zonation, albeit correlated with the structural-physiognomic, is defined by changes in quantitative abundances rather than qualitative floristic differences.
In answer to my opening questions, I conclude, 1) that the forest elevational zones now well accepted on wet tropical mountains do indeed remain valid under conditions of increasing diurnal temperature variability and decreasing minimum temperature, and with increasing rainfall seasonality; 2) that although the lower ecotone of equatorial upper montane forest does vary in altitude, those of lower montane forest and of subalpine thicket hardly do so; but upper montane forest is replaced by lower montane forest in climates with a short cloudy season, which in turn becomes increasingly truncated by winter frost in drier climates and at increasing latitudes; 3) floristic change with elevation correlates quantitatively with zones defined by structure and physiognomy, but few tree species are sufficiently confined to zones to be diagnostic.
Acknowledgements
This review has been much strengthened by the comments and suggestions of two reviewers, anonymous but clearly leading authorities in the wonderful field of mountain ecology; and by the patience and invaluable suggestions of our skilful editor.
Disclosure statement
No potential conflict of interest was reported by the author.
Additional information
Notes on contributors
Peter S. Ashton
This is the sixtieth year of Peter Ashton’s field studies in the floristic and dynamic ecology of tropical Asian forests, and the systematics of their dominant canopy tree family, Dipterocarpaceae; starting with a decade as a forest service botanist, and still continuing after retirement as Bullard Professor of Forestry at Harvard.
References
- Ahluwalia KS. 1965. Contributions to the flora of Mt. Abu. The Indian Forester. 91:824–828.
- Aiba S-I, Kitayama K. 1999. Structure, composition and species diversity in an altitude-substrate matrix of rainforest tree communities on Mount Kinabalu, Borneo. Plant Ecology. 140:139–157.
- Aiba S-I, Kitayama K. 2002. Effects of the 1997-98 El Niño drought on rain forests of Mt. Kinabalu, Borneo. Journal of Tropical Ecology. 18:215–230.
- Aiba S-I, Suzuki E, Kitayama K. 2006. Structural and floristic variation among small replicate plots of a tropical montane forest on Mount Kinabalu, Sabah, Malaysia. Tropics. 15:219–236.
- Aiba S-I, Takyu M, Kitayama K. 2005. Dynamics, productivity, and species richness of tropical rainforests along elevational and edaphic gradients on Mount Kinabalu, Borneo. Ecological Research. 20:279–286.
- Ashton PS. 2003. Floristic zonation on wet tropical mountains revisited. Perspectives in Plant Ecology. 6:87–104.
- Ashton PS. 2014. On the Forests of tropical Asia, lest the memory fade. Royal Botanic Gardens, Kew and Arnold Arboretum, Harvard University.
- Beaman JH, Anderson C, Beaman RS. 2001. The plants of Mount Kinabalu. 4. dicotyledon families acanthaceae to lythraceae. Kota Kinabalu: Natural History Publications (Borneo) Sendirian Berhad.
- Beaman JH, Anderson C, Beaman RS. 2004. The plants of Mount Kinabalu.5. Magnoliaceae to Winteraceae. Kota Kinabalu: Natural History Publications (Borneo) Sendirian Berhad.
- Beaman JH, Beaman RS. 1998. The plants of Mount Kinabalu. 3. Gymnosperms and non-orchid monocotyledons. Kota Kinabalu: Natural History Publications (Borneo) Sendirian Berhad.
- Blasco F. 1971. Montagnes du sud de l’Inde. Pondicherry: French Institute.
- Brown GK, Nelson G, Et Ladiges PY. 2006. Historical biogeography of Rhododendron section Vireya and the Malesian Archipelago. Journal of Biogeography. 33:1929–1944.
- Bruijnzeel IA, Veneklaas EJ. 1998. Climatic conditions and tropical montane forest productivity: the fog has not lifted yet. Ecology. 79:3–9.
- Burnham CP. 1984. Soils. Chapter 11. In: Whitmore. ibid.; p. 137–154.
- Champion HG. 1936. A preliminary survey of the forest types of India. Indian Forest Records, New Series. Silviculture. 1.
- Collins NM. 1980. The distribution of soil macrofauna on the west ridge of Gunung (Mount) Mulu, Sarawak. Oecologia. 44:263–275.
- Culmsee H, Leuschner C. 2013. Consistent patterns of elevational change in tree taxonomic and phylogenetic diversity across Malesian mountain forests. Journal of Biogeography. 40:1997–2010.
- Dames TWG. 1955. The soils of East-Central Java. In: Contributions of the general agricultural stations, Bogor. p. 141.
- Davidar P, Mohandas D, Vijayan SL. 2007. Tropical Ecology. Vol. 48, p. 15–25.
- Davidar P, Puyravaud J-P, Leogh EG. 2005. Changes in rainforest tree diversity, dominance and rarity across a seasonality gradient in the Western Ghats, India. Journal of Biogeography. 12:493–501.
- Deshpande S, Sharma BD, Nayar MP. 1993. Flora of Mahabeleshwar and adjoinings, Maharashtra. Calcutta: Botanical Survey of India.
- Gillison AN. 1970. Structure and floristics of a montane grassland/forest transition, Doma Peaks region, Papua. Blumea. 18:71–86.
- Greller AM,Balasubramanyam S. 1988. Vefetational composition, leaf size, and climatic warmth in an altitudinal sequence of evergreen forests in Sri Lanka (Ceylon). Tropical Ecology. 29:121–145.
- Greller AM, Gunatilleke IAUN, Jayasuriya AHM, Gunatilleke CVS, Balasubramanyam S,Dassanayake MD 1987. Stemonoporus (Dipterocarpaceae)-dominated montane forests in the Adam’s peak Wilderness, Sri Lanka. Journal of Tropical Ecology. 3:243–253.
- Grierson AJC, Long DG, Noltie HJ. 1983-2001. Flora of Bhutan. Vol. 3. Edinburgh: Royal Botanic Garden.
- Grubb PJ. 1977. Control of forest growth and distribution on wet tropical mountains with special reference to mineral nutrition. Annual Review of Ecology and Systematics. 8:83–107.
- Grubb PJ. 1973. Factors controlling the distribution of forest types on tropical mountains: new facts and a new perspective. In: Hull Geog. Dept. Misc. Ser. 13 Flenley JR, ed. Transactions of the Third Aberdeen-hull symposium on Malesian ecology. 13–46.
- Grubb PJ, Lloyd JR, Pennington TD, Whitmore TC. 1963. A comparison of montane and lowland forest in Ecuador.I. The forest structure, physiognomy and floristics. Journal of Ecology. 31:567–601.
- Grubb PJ, Stevens PF 1985. The forests of the Fatima Basin and Mount Kerigomna and a review of montane and subalpine forests elsewhere in New Guinea. Australian National University BG 8.
- Haantjens HA, Rajenders JJ, Mouthaan WNLPJ, Van Baren FA. 1967. Major soil groups of New Guinea and their distribution. Communication 55. Amsterdam: Departtment of Agricultural Research, Royal Tropical Institute.
- Hope GS. 1976. The vegetation of Mt. Jaya. In: Hope GS, al. eds. The equatorial glaciers of New Guinea. Rotterdam: Balkema; p. 113–172.
- Hope GS. 1980. New Guinea mountain vegetation communities. In: Van Royen P, Cramer J, eds. Liechtenstein, 153–223
- Hynes RA 1974. Nothofagus Forest in New Guinea. An Ecological Introduction. Unpubl. Ms.
- Jain SK. 1968. Phytogeographic considerations of the flora of Mt. Abu. Bulletin, Botanical Survey of India. 9:68–78.
- Janzen DH. 1967. Why mountain passes are higher in the tropics. American Naturalist. 101:233–249.
- Jenny H. 1941. Factors of Soil Formation: A system of quantitative pedology. Reprint 1994 Dover (New York): McGraw Hill, New York.
- Johns RJ, Stevens PF. 1972. Mount Wilhelm flora. A Checklist of the Species. In: Botany Bulletin 6. New Guinea: Div. Bot. Dept. For. Lae.
- Kanaki M, Hara M, Yamakura T, Ohkubo T, Tamura MN, Sri-Ngernyuang K, Sahunalu P, Teejuntul S, Bunyajejchewin S. 2004. Doi inthanon forest dynamic plot, Thailand. In Losos EC, Leigh Jr EG, editors. Tropical forest diversity and dynamism: findings from a large-scale plot network. University of Chicago Press; p. 474–481.
- Kanzaki M, Sri-Ngernyuang K, Fuji K, Mizuno T, Noguchi H, Yamakura T, Hara M, Ohkubo T, Sahunalu P, Dhanmonand P, et al. 1999. Comparative community ecology of tropical montane and temperate lucidophyll forests. Comparison of niche differentiation pattern. Tropics. 9:211–228.
- Kerr AFG. 1933. A trip to Phu Bia in French Laos. Jourhal of the Siam Society. 9:193–223.
- Kingdon Ward F. 1958. A sketch of the flora and vegetation of Mt. Victoria in Burma. Acta Horti Gotoburgensis. 22:53–74.
- Kira T. 1977. A climatological interpretation of Japanese vegetation zones. In: Mayawaki Et A, Tüxen R, eds. Vegetation Science and Environmental Protection. Tokyo: Maruzen. p. 21–30.
- Kitayama K. 1992. An altitudinal transect study of the vegetation of Mt. Kinabalu, Borneo. Vegetatio. 102:149–171.
- Kitayama K. 1996. Climate of the summit region of Kinabalu (Borneo) in 1992, an El Niño year. Mountain Research and Development. 16:65–75.
- Kitayama K, Aiba S-I. 2002. Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. Journal of Ecology. 90:37–51.
- Körner C. 1998. A reassessment of high altitude treeline positions and their explanation. Oecologia. 115:445–459.
- Körner C. 2012. Alpine tree lines. Functional ecology of the global high elevation tree limits. Basel: Springer.
- Körner C, Paulsen J, Spehn E. 2011. A definition of mountains and their bioclimatic belts for global comparisons of biodiversity data. Alpine Botany. 121:73–78.
- Kudo G 2001. Unpubl. Data.
- Lane-Poole CE. 1925. The Forests of Papua and New Guinea. Emp Forest Journal. 4:206–234.
- Mangen J-M. 1993. Ecology and Vegetation of Mount Trikora, New Guinea (Irian Jaya, Indonesia). Travaux Scientifiques du Musée d’Histoire Naturelle de Luxembourg.
- Nagy L, Grabherr G. 2009. The biology of alpine habitats. Oxford University Press.
- Ohsawa M. 1993. Latitudinal patterns of mountain vegetation zonation in southern and eastern Asia. Journal of Vegetation Science. 4:13–18.
- Ohsawa M. 1995. Latitudinal comparison of altitudinal change in forest structure, leaf type, and species richness in humid monsoon Asia. Vegetatio. 121:3–10.
- Ohsawa M. 2006. Climate change impacts on vegetation in humid Asian mountains. Global Environment Research. 10:13–20.
- Ohsawa M, Nainggolan PH, Anwar C. 1985. Altitudinal zonation of forest vegetation on Mount Korinci, Sumatra: with comparisons of zonation in temperate regions of East Asia. Journal Of Tropical Ecology. 1:293–316.
- Proctor J, Lee YF, Langley AM, Munro WRC, Nelson T. 1988. Ecological studies on Gunung Silam, a small ultrabasic mountain in Sabah, Malaysia. I. Environment, forest structure and floristics. Journal of Ecology. 76:320–340.
- Proctor J, Phillips C, Duff G, Heaney A, Robertson FM. 1989. Ibid.: II. Some forest processes. Journal of Ecology. 77:317–331.
- Renner SS, Strijk JS, Strasberg D, Thébaud C. 2010. Biogeography of the Monimiaceae (Laurales): a role for East Gondwana and long-distance dispersal, bu not West Gondwana. Journal of Biogeography. 37:1227–1238.
- Richards PW. 1996. The tropical rain forest. An ecological study. 2nd ed. Cambridge University Press.
- Saxena HO, Brahmam M. 1989. The flora of similipahar (Similipal), Orissa, with particular reference to the potential economic plants. Bhubaneswar: Regional Research Laboratory.
- Smith CW. 1995. Lichens as indicators of cloud forest in Hawai’i. In: Hamilton LS, Juvik JO, Scatena FN, eds. Tropical Montane cloud forests. Proceedings of an international symposium. Ecological studies. Vol. 110. 309–314.
- Smitinand T. 1966. The vegetation of Doi Chiengdao, a limestone massif in Chiengmai, North Thailand. Natural History Bulletin of the Siam Society. 21:93–128.
- Smitinand T 1968. The vegetation of Khao Yai National Park. 22: 289–305.
- Soepadmo E. 1976. Fagaceae. In: Van Steenis CGGJ, ed. Flora Malesiana. Vols. 1, 7. 265–403.
- Spjut RW. 2007. Journal of the Botanical Research Institute of Texas 1:291–332.
- Stainton JDA. 1972. The Forests of Nepal. London: John Murray.
- van Steenis CGGJ. 1972a. The mountain flora of Java. Brill (Leiden).
- van Steenis CGGJ. 1972b. The mountain flora of the Malaysian tropics. Endeavour. 21:183–193.
- van Steenis CGGJ. 1984. Floristic altitudinal zones in Malesia. Botanical Journal of the Linnean Society Soc. 89:289–292.
- Vink W. 1970. The Winteraceae of the Old World. I. Pseudowintera and Drimys. Blumea. 18:225–354.
- Wade LK, McVean DN. 1969. Mount Wilhelm Studies 1. The Alpine and Subalpine Vegetation. Canberra: Australian National University Press.
- Walter H, Harnickell E, Mueller-Dombois D. 1975. Climate-diagram maps of the ecological regions of the earth. Berlin: Springer.
- Walter H, Lieth H. 1960. Klimadiagramm Weltatlas. Jena: G. Fischer.
- Whitmore TC. 1984. Tropical rain forests of the far east. 2nd ed. Oxford: Clarendon Press.
- Whitmore TC. 1988. Phytogeography of the eastern end of Tethys. In: Audley-Charles MG, Hallam A, eds. Gondwana and Tethys. Oxford University Press. p. 307–311.
- Whitmore TC, Burnham DFRP. 1969. The altitudinal sequence of forests and soils on granite near Kuala Lumpur. Malayan Nature Journal. 22:99–118.
- Yamada I. 1976. Forest ecological studies of the montnae forest of Mt. Pangrango, West Java. II. The stratification and floristic composition of the forest vegetation of the higher part of Mt. Pangrango. Tonan Ajia Kenku (Southeast Asian Studies). 13:4.