
ABSTRACT
Background
Nepenthes clipeata is a Critically Endangered plant species with the population at its only location in the wild, Gunung (Mount) Kelam in Kalimantan, Indonesia, under threat – its nutrient relationships have not been studied.
Aims
To improve understanding of the autecology of N. clipeata by assessing the current population and providing information on its mineral nutrient relationships.
Methods
A survey was undertaken in 2019 when population numbers and habitat preferences of N. clipeata were recorded. Nutrient concentrations in soil, leaves and pitcher fluid were analysed and compared with those in other Nepenthes species.
Results
Eighteen individuals of N. clipeata were found on Gunung Kelam (only one of which was female). Seven other Nepenthes species were found and hybrids with two of these and N. clipeata were observed. Although the foliar nutrient concentrations (‘ionome’) of N. clipeata appeared distinct from that of other Nepenthes species, with N. clipeata having greater nutrient concentrations, particularly nitrogen, potassium and calcium, nitrogen limitation was still prevalent.
Conclusions
N. clipeata still persists in the wild but with very low numbers, indicating that conservation actions are required. Along with other species of Nepenthes, it is limited by soil nitrogen concentrations although it has a distinct ionome derived from high foliar nutrient concentrations.
Introduction
Nepenthes (Nepenthaceae) is a Palaeotropical genus of dioecious pitcher plants comprising around 180 species distributed from Madagascar to New Caledonia with by far the greatest diversity in the Philippines and Indonesia. The centre of diversity of Nepenthes is Borneo: some species found there, such as N. ampullaria Jack, N. gracilis Korth., N. rafflesiana Jack and N. reinwardtiana Miq. (authorities follow www.ipni.org), have broad distributions, but there are also over 30 endemic species recorded from the island (Clarke Citation1997). Many of these are highly localised and only found on a single mountain or mountain range such as N. faizaliana J.H.Adam & Wilcock (Gunung Mulu), N. murudensis Culham ex Jebb & Cheek (Gunung Murud), N. pilosa Danser (Bukit Batu Lesung), N. rajah Hook.f. (Gunung Mulu and Gunung Tambuyukon) and, the focus of this paper, N. clipeata Danser. Nepenthes are usually found growing on infertile edaphic substrates such as heath forest white sands, ultramafic soils, montane forests and, less frequently, limestone soils. The key morphological adaptation that these species possess is their pitcher, which is a jug-like container developed at the end of the leaves that is modified to hold liquid and has a slippery peristome and/or waxy zone to trap and retain prey (Clarke Citation1997; Mansur Citation2006). However, there are a wide range of pitcher forms that have diverged evolutionarily to target contrasting prey spectra (Lam et al. Citation2018; Thorogood et al. Citation2018). These captured prey provide additional nutrients to the plant and help them to grow in nutrient-poor soils.
Nepenthes clipeata () is an exceptionally rare species found only on Gunung (Mount) Kelam, West Kalimantan, Indonesian Borneo. It was first discovered by Johannes Gottfried Hallier in 1894 and first determined by Danser (Citation1928). However, this point endemic N. clipeata population is now severely depleted due to over-collection by illegal plant harvesting and recent fires related to El Niños (Lee Citation2009). In 1995, Jebb (in Simpson Citation1995) reported 15 individuals remaining, Clarke (in Cheek and Jebb Citation2001) suggested only 2 to 6 plants might remain and Lee (Citation2009) reported just ‘a few’ individuals remaining. Recently, Lovadi (Citation2010) reported 260 individuals from 45 points between 765 and 875 m a.s.l. and Anonymous (Citation2011) found an additional 63 individuals from six points between 535 and 665 m a.s.l. However, the population is still in a precarious situation in the wild with reports of illegal plant collecting as of 2019 by skilled plant ‘poachers’ and, according to the International Union for Conservation of Nature and Natural Resources (IUCN), this species is classified as ‘Critically Endangered’ (Clarke and Lee Citation2014).
Figure 1. (a) Typical habitat, (b) pitcher and (c) flowering plant of Nepenthes clipeata growing on the granite rock at Gunung Kelam, West Kalimantan, Indonesia. Photographic credit: M. Mansur.

It is considered that Nepenthes are nitrogen (N) limited (Brearley and Mansur Citation2012), but the nature of nutrient limitation may differ depending on the soil type on which plants grow. For example, whilst heath forests are considered N limited (Brearley et al. Citation2011; Sellan et al. Citationin revision), ultramafic soils may be more phosphorus (P) limited (Brearley Citation2005; Porder and Ramachandran Citation2013). One way we can assess nutrient limitation is through an examination of nutrient concentrations and ratios in plant tissue (usually leaves) and, using this approach, Ellison (Citation2006) has suggested that the majority of carnivorous plants were N and P limited but noted a lack of data for Nepenthes. Examination of the suite of mineral nutrients present in a plant’s tissue (‘ionome’) can shed further light on its adaptation to a given environment. For example, calcicoles (plants restricted to limestone substrates) may have a greater physiological requirement for calcium (Ca) than calcifuges that avoid this habitat (White and Broadley Citation2003) and plant nutrient requirements may allow hypotheses related to species edaphic preferences to be developed. The foliar ionome in relation to substrate preference has been poorly explored for the tropical flora and not at all for Nepenthes species.
The aim of this study was to provide an update on the status of N. clipeata at Gunung Kelam and present additional data on its autecology with reference to mineral nutrition by determining the nutrient concentrations of leaves, pitcher fluid and soil around the plant. We anticipate that this new information on its nutritional requirements via examination of its foliar ionome will shed light on potential edaphic habitat preferences of this very rare species.
Materials and methods
Study site
Gunung Kelam is an immense monolithic granite dome – one of the largest in the world besides Uluru in Australia. It is located in Kelam Permai District, Sintang Regency, West Kalimantan, around 20 km from the town of Sintang and is protected as a Taman Wisata Alam (Nature Tourism Park) with an area of 520 hectares (Gustamansyah Citation2019). The granite dome has very steep cliffs with many areas that are inaccessible due to this topography. The climate is type ‘A’ according to Schmidt and Fergusson (Very wet) with a mean (± S.D.) annual precipitation of 3160 ± 661 mm between 2000 and 2017 (Mansur et al. in review).
Population estimation
A preliminary survey of the individuals of N. clipeata found on Gunung Kelam was made in March 2019 (two people surveying for three days), with a more extensive survey made in September 2019 (eight people surveying for three days including the use of ropes to access steep areas) during which we conducted searches for the target species focusing on an area of about 4 ha where it was previously reported. Locations of all individuals was recorded with a Garmin GPSMAP 64s. Additional records of other species of Nepenthes present were made, along with their elevation and habitat type.
Nutrient relationships
Samples of leaves, pitcher fluid and soil were collected for nutrient analyses (N, P, potassium (K), Ca, magnesium (Mg) and sodium (Na)) from three plants growing at 800 to 820 m. The second youngest leaves (estimated as six months old) were collected and dried (50°C for 5 days). Soil and leaf N was analysed with a Yanako JM macro corder 1000 CN with a JMA auto sampler 1000. For other nutrients, plant and soil materials were digested in mixed acid (H2SO4, HClO4 and HNO3) for 24 hours at 170°C or filtered (pitcher fluid) and analysed for K, Ca, Mg and Na using atomic absorption spectrophotometry (Shimadzu AA-6200) and N (pitcher fluid) and P using a Shimadzu UV Mini-1240 spectrophotometer and the Nessler and vanadomolybdate colorimetric methods, respectively. Additionally, the chlorophyll concentration was measured on three leaves per plant using a Minolta SPAD-502 meter.
Statistical analyses
The nutrient composition among leaves, pitcher fluid and soil was compared using a one-way ANOVA and Tukey’s test and visualised (N, P and K only) on a ternary diagram. To compare the foliar nutrient composition between N. clipeata and other Nepenthes species, another ternary diagram was created and a principal component analysis (using foliar N, P, K Ca and Mg concentrations) on scaled and centred (z-scores) values was conducted. Stoichiometric distinctiveness was calculated as in Trethowan et al. (Citation2021) and represents the average difference between a species and all others in multivariate space, the axes of which are the concentrations of each leaf nutrient. To determine the spatial distribution of the species, Morisita’s (Citation1959) index of dispersion was calculated using a quadrat size of 50 m x 50 m with locations derived from the GPS points determined as noted above.
Results
Habitat
Nepenthes clipeata grows in open places directly exposed to sunlight on rock cliffs ()) or in places with running water where the growing media is moss (Sphagnum cuspidatum Ehrh. ex Hoffm.; Sphagnaceae) and humus associated with rumput lambang (Panicum sarmentosum Roxb.; Poaceae). Some N. clipeata individuals were also found growing under the shade of pohon embun (Seorsus aequatorius Rye & Trudgen; Myrtaceae) on rock slopes associated with grasses and sedges (Panicum sarmentosum and Scleria oblata S.T.Blake ex J.Kern; Cyperaceae) and terrestrial orchids (Arundina graminifolia (D.Don) Hochr., Spathoglottis gracilis Rolfe ex Hook.f., Paphiopedilum bullenianum Pfitzer and Phaius sp., all Orchidaceae) (Mansur et al. in review). Although N. clipeata was found growing under the shade of trees, it was exclusively prostrate on the surface of the ground and not growing epiphytically.
Population estimate of N. clipeata and the presence of other Nepenthes species
We found 18 plants of N. clipeata between 765 m and 835 m a.s.l. in an area of 3.9 ha. As the majority of plants were in flower, we determined the sex of 14 of these and all but one were male. Morisita’s index of dispersion was 7.71, indicating a clumped distribution. An additional seven species of Nepenthes were found (). Nepenthes gracilis, N. mirabilis (Lour.) Druce and N. reinwardtiana were generally found growing together in open, shrubby thickets of Gleichenia sp. (Gleicheniaceae) and Pteridium aquilinum (L.) Kuhn (Dennstaedtiaceae) between 300 and 800 m a.s.l., whereas N. albomarginata T.Lobb ex Lindl., N. ampullaria and N. rafflesiana were found together in more shaded areas between 700 and 900 m a.s.l. We did not enumerate the individuals of these species; we noted three individuals of N. bicalcarata Hook.f. at an elevation of 540 m a.s.l. in a somewhat closed area as also reported by Hallier (Citation1895). One of each of the hybrids between N. clipeata and N. reinwardtiana and N. albomarginata were found where they grew sympatrically ().
Table 1. Nepenthes species found on Gunung Kelam, West Kalimantan, Indonesia, along with their habitat and IUCN conservation status
Figure 2. (a) Nepenthes clipeata x albomarginata and (b) Nepenthes clipeata x reinwardtiana growing on Gunung Kelam, West Kalimantan, Indonesia. Photographic credit: M. Mansur.

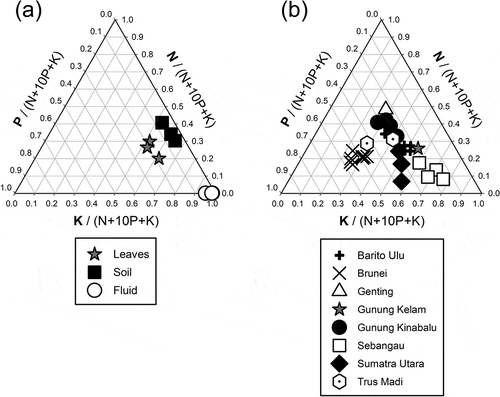
Figure 3. Ternary diagrams of (a) ratios of nitrogen, phosphorus and potassium in soil, leaves and pitcher fluid of Nepenthes clipeata on Gunung Kelam, West Kalimantan, Indonesia, and (b) a comparison of leaves between eight sites in Sundaland. Data are sourced from Brearley (unpubl. data) for Barito Ulu, Osunkoya et al. (Citation2007, Citation2008)) for Brunei, Brearley (Citation2021) for the Genting Highlands, van der Ent et al. (Citation2015, pers. comm.) for Gunung Kinabalu and Trus Madi, Brearley and Mansur (Citation2012) for Sebangau and Mansur (unpubl. data) for Sumatra Utara.
Nutrient relationships
Nutrients were at greater concentrations in the leaves than the soil with the exception of N and Na (). The leaves were low in essential nutrients with a mean N:P ratio of 13.6. The pitcher fluid was particularly low in all nutrients with the exceptions of K and Na (). The positions of all three on a ternary diagram () were suggestive of N limitation for all of them, following the criteria of Olde Venterink et al. (Citation2003), i.e. N:P < 14.5 and N:K < 2.1. The ternary diagram and principal component analysis examining the foliar nutrient composition of N. clipeata in relation to other Nepenthes species showed greater nutrient concentrations in N. clipeata than other species for many of the nutrients ( and ), with, notably, N, K and Ca leading to N. clipeata having a high stoichiometric distinctiveness () (the majority of more distinct species were from peatland with high foliar K concentrations). The mean (± S.D.) chlorophyll concentration was 44.6 ± 6.6 SPAD meter units.
Table 2. Nutrient concentrations of surrounding soil, pitcher fluid and leaves of Nepenthes clipeata on Gunung Kelam, Kalimantan, Indonesia (n = 3 for each). All values are mean ± standard error; letters indicate significant differences for each nutrient according to Tukey’s tests
Figure 4. (a) Principal component analysis and (b) stoichiometric distinctiveness of the foliar ionome (N, P, K, Ca and Mg) of Nepenthes clipeata on Gunung Kelam, West Kalimantan, Indonesia, and other Nepenthes species from six locations in Sundaland. For (a), the two bar graphs in the bottom right of the panel represent the loadings on the two principal component axes; for (b), greater values on the y-axis indicate a more distinct ionome, i.e. the average difference between a species and all others in a multivariate space, the axes of which are the leaf nutrient concentrations. Where a habitat is noted after a species name, this indicates that this species’ ionome has been determined in two contrasting habitats by different studies. See Figure 3 legend for references to data sources.
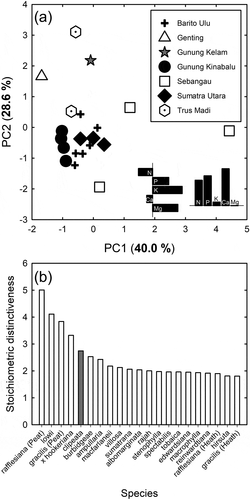
Figure 5. Dead specimens of Nepenthes clipeata on Gunung Kelam, West Kalimantan, Indonesia, that are the remains of plants left by poachers. Photographic credit: M. Mansur.

Discussion
We show here that the Critically Endangered pitcher plant N. clipeata still persists at its only known locality: Gunung Kelam in western Indonesian Borneo. However, the number of plants remains critically low and poaching of these plants still appears to be occurring (M.M. personal observation; ) despite the remaining plants being found in areas of the mountain that are difficult to access. Whilst dioecious plants are often male-biased in their populations, the significant imbalance between male and female N. clipeata plants at Gunung Kelam could be due to increasing levels of environmental stress known to further bias populations towards males (Munné-Bosch Citation2015; Hultine et al. Citation2016). This skewed sex ratio reduces the effective population size and has the potential to be highly limiting for the viability of the remaining population and difficult to rectify. The hybridisation seen is also suggestive of habitat disturbance following fires (see Heon and Clarke Citation2015). The IUCN Red List classification of ‘Critically Endangered A2a,d’ (i.e. direct observation of a reduction in population size ≥ 80% over the last 10 years due to ongoing exploitation) still appears appropriate, although it could also be considered ‘Critically Endangered B2a,b(v)’ (i.e. found at a single location with an area of occupancy of less than 10 km2 showing a decline in the number of individuals). Although the population decline appears to have slowed recently, effective and repeatable measures of the population size are needed within the area of occupancy of less than 10 km2 to assess recent and future changes.
The soil nutrient concentrations at the study site were fairly high as the soil was organic and mostly derived from decaying plant material. Leaf nutrient concentrations were generally greater than those in the soil as leaves are a place for storing nutrients that are absorbed from the soil. The foliar N:P ratio of c. 14 is suggestive of primary N limitation (i.e. N:P < 14.5 and N:K < 2.1 as noted above), confirming earlier results on Nepenthes (Brearley and Mansur Citation2012; Brearley Citation2021), and differs somewhat from other carnivorous plants that appear co-limited by N and P (Ellison Citation2006). When our data are compared with that of other researchers from a range of sites across Sundaland (Osunkoya et al. Citation2007, Citation2008; Brearley and Mansur Citation2012; van der Ent et al. Citation2015; Brearley Citation2021; Mansur, unpubl. data), it can be seen that the vast majority of Nepenthes species are N limited according to the criteria proposed by Ellison (Citation2006) building on studies of Olde Venterinck et al. (Citation2003). When compared with other sites, and similar to Sebangau peat swamp forest (Brearley and Mansur Citation2012), there was no indication of K deficiency as K is fairly high in granitic rocks (Manning Citation2010). An assessment of habitat preferences allows consideration of the breadth of habitats a plant can colonise, such as ultramafic substrates, and we show here that N. clipeata appears to have an ionomic composition distinct from other Nepenthes species, which is due to its greater nutrient concentrations not only N and K but also P and Ca, perhaps limiting it to more nutrient-rich substrates than localities at which other Nepenthes are found.
Interpreting N:P ratios in the pitcher fluid in terms of nutrient limitation is more challenging, but there was less N than P in the fluid indicating that it was likely to be taken up in preference and/or more rapidly. We present here one of the first chemical analyses of Nepenthes pitcher fluid under natural conditions, although the low N and P in the pitcher fluid were also seen by Buch et al. (Citation2013) in young greenhouse-grown plants who also found that K and Na were high. The pitcher liquid contains a complex microbial community that is influenced by the liquid chemistry (Gilbert et al. Citation2020) so understanding pitcher fluid composition allows better prediction of the infaunal and microbial community.
As the plant is a creeping lithophyte, it only appears to produce one kind of pitcher, i.e. upper pitchers (Cheek and Jebb Citation2001). The chlorophyll concentrations were at the lower half of the range reported by Mansur (Citation2017) for fifteen Nepenthes taxa and, given that chlorophyll concentrations correlate with stomatal conductance (Mansur Citation2017), indicate that N. clipeata is more water-use efficient than other Nepenthes species. The open environment on Gunung Kelam, where the plants grow lithophytically, will have low humidity and the advantages of a slippery, wettable peristome are reduced if the peristome dries out rapidly after rain. The peristome of N. clipeata is very small and is of the ‘dry type’ (Bauer et al. Citation2012; Moran et al. Citation2013). Knowing this, a better understanding of its prey spectra would be informative. It would also be useful to determine if there are any unusual or species-specific infauna (e.g. Rembold et al. Citation2012; Karl and Bauer Citation2020) that are obligate mutualists of N. clipeata and therefore also of conservation concern.
Whilst we found 18 individuals of N. clipeata in total, the comparison of our population survey data with those of earlier studies is difficult as it is not known if individual plants or stems were enumerated by others. To the best of our knowledge, our study location overlaps with that of earlier researchers (Lovadi Citation2010; Anonymous Citation2011) who reported a greater number of plants than earlier reported censuses (Jebb in Simpson 1995; Clarke in Cheek and Jebb Citation2001; Lee Citation2009) as they counted each stem as an individual plant. In this study, we counted each individual stem that emerged from the ground, although it is possible that they were connected under the soil surface. In our study, N. clipeata was found at elevations between 765 and 835 m in open or slightly shaded areas. However, earlier reports that N. clipeata was found down to an elevation of 575 m suggest that it may still be possible to find individuals in other locations that are difficult to access (although the N. clipeata individual at an altitude of 500 m a.s.l. found by Lovadi (Citation2010) appears to be no longer there). The population was highly aggregated, meaning that it may be at greater risk from collectors who are likely to encounter additional plants once an initial one is found. It is encouraging that local authorities are implementing efforts to prevent poaching of N. clipeata by blocking access to its habitat for climbers, implementing tighter security at the Park entrance (M.M. pers. obs.) and bringing legal sanctions against poachers (Pontianak Tribune Citation2020), but, with such a small population, the continued existence of N. clipeata in the wild remains tenuous.
Further conservation efforts for N. clipeata have been proposed for many years including propagation techniques to bring plants into cultivation, flooding the market with cultivated plants and thus relieving collecting pressure on the remaining natural population (Lee Citation2009). Nepenthes clipeata already exists in cultivation with many plants produced by micro-propagation and, although the number of clones in cultivation is limited (Cantley et al. Citation2005), it represents an additional genetic source to help with ex situ conservation. Indeed, ex situ conservation through plant propagation and then re-introducing plants to their natural habitat with strict monitoring appear to be the most appropriate conservation approach in this instance. Research should also focus on understanding the environmental preferences of the species (as done partly in this paper) as well as its reproductive and regeneration dynamics and how these might differ between sexes. Further work should also examine the genetic diversity and viability of the Gunung Kelam populations and consider these in the light of the possibility for re-introduction in the future.
Conclusions
Nepenthes clipeata still persists in the wild at its only known location but with very low numbers and an imbalanced sex ratio. It appears that it has an ionomic composition distinct from many other species of Nepenthes derived from high foliar nutrient concentrations. Illegal plant poaching still occurs, indicating that conservation actions are urgently required for the persistence of this species in its natural habitat.
Supplemental Figure
Download JPEG Image (860.1 KB){kind=link}
Acknowledgements
We thank Simon Dowell, Scott Wilson, Mike Jordan, Hanna Khwaja and Stuart Young, staff of Chester Zoo who supported this research. We also thank the Head of Research Center for Biology-LIPI and the Head of Botany Division, the Natural Resources Conservation Centre, Pontianak and Sintang who granted the research permits (SI.61/BKSDA.KALBAR/KK/9/2019), Liam Trethowan for helping with analyses and the expedition team to Gunung Kelam: Fauzi Rachmat, Supardi Jakalalana (RCB-LIPI), Gustamansyah (PEH BKSDA-Sintang), Donni Mahyudi Erpah (FPTI-Sintang), Edi Setyawan, Arnol Raimon Yudianto, Dedimus Fokas (Kapuas University) and our field guide Yusran Efendi Ritonga.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
Notes on contributors
Muhammad Mansur
Muhammad Mansur is a plant ecologist with an interest in the composition, population structure and ecophysiology of Indonesian tropical forest plants and trees. He has a special fascination for Nepenthes pitcher plants.
Francis Q. Brearley
Francis Q. Brearley is a tropical ecologist with an interest in the functional importance of plant-soil interactions for ecosystem processes. He has a particular focus on the forests and ecosystems of South-east Asia.
Philip J. Esseen
Philip J. Esseen curates the UK National Collection of Nepenthes at Chester Zoo and is interested in actively promoting the in situ and ex situ conservation of this family and other threatened plants.
E. Johanna Rode-Margono
E. Johanna Rode-Margono specialises in the ecology and conservation of South-east Asian species. She has a special interest in taxonomic groups that have been overlooked by previous conservation or applied research efforts.
M. Rafi’i Ma’arif Tarigan
M. Rafi’i Ma’arif Tarigan Tarigan is a lecturer in education and science. He focuses on Nepenthes taxonomy and ecology and has conducted research on Nepenthes pitcher plants in the forests of the islands of Java, Kalimantan, Sulawesi and Sumatra in Indonesia.
Related Research Data
References
- Anonymous. 2011. Inventarisasi Nepenthes clipeata, Taman Wisata Alam Bukit Kelam, Kabupaten Sintang, Kalimantan Barat, Indonesia: Balai Konservasi Sumber Daya Alam Pontianak, Kalimantan Barat.
- Bauer U, Clemente CJ, Renner T, Federle W. 2012. Form follows function: morphological diversification and alternative trapping strategies in carnivorous Nepenthes pitcher plants. J Evol Biol. 25(1):90–102. doi:https://doi.org/10.1111/j.1420-9101.2011.02406.x.
- Brearley FQ. 2005. Nutrient limitation in a Malaysian ultramafic soil. J Trop For Sci. 17:596–609.
- Brearley FQ. 2021. Nutrient and metal concentrations in Nepenthes macfarlaneii Hemsl. (Nepenthaceae) from a Malaysian montane forest. Notulae Sci Biol. 13(2):10976. doi:https://doi.org/10.15835/nsb13210976.
- Brearley FQ, Fine PVA, Perreijn K. 2011. Does nitrogen availability have greater control over the formation of tropical heath forests than water stress? A hypothesis based on nitrogen isotope ratios. Acta Amazon. 41(4):589–592. doi:https://doi.org/10.1590/S0044-59672011000400017.
- Brearley FQ, Mansur M. 2012. Nutrient stoichiometry of Nepenthes species from a Bornean peat swamp forest. Carniv Plant Newsl. 41:105–108.
- Buch F, Rott M, Rottloff S, Paetz C, Hillke I, Raessler M, Mithöfer A. 2013. Secreted pitfall-trap fluid of carnivorous Nepenthes plants is unsuitable for microbial growth. Ann Bot. 111(3):375–383. doi:https://doi.org/10.1093/aob/mcs287.
- Cantley R, Clarke CM, Cokendolpher J, Rice B, Wistuba A. 2005. Nepenthes clipeata survival project. Carniv Plant Newsl. 34:116–120.
- Cheek M, Jebb M. 2001. Nepenthaceae. Flora Malesiana Ser I - Seed Plants. 15:1–157.
- Clarke CM. 1997. Nepenthes of Borneo., Kota Kinabalu, Sabah, Malaysia: Natural History Publications.
- Clarke CM, Lee C 2014. Nepenthes clipeata. The IUCN Red List of Threatened Species 2014: e.T39652A19631488; [accessed 2020 May 18]. doi:https://doi.org/10.2305/IUCN.UK.2014-1.RLTS.T39652A19631488.en
- Danser BH. 1928. The Nepenthes of the Netherlands Indies. Bulletin du Jardin Botanique de Buitenzorg Série III. 9:249–438.
- Ellison AM. 2006. Nutrient limitation and stoichiometry of carnivorous plants. Plant Biol. 8(6):740–747. doi:https://doi.org/10.1055/s-2006-923956.
- Gilbert KJ, Bittleston LS, Tong W, Pierce NE. 2020. Tropical pitcher plants (Nepenthes) act as ecological filters by altering properties of their fluid microenvironments. Sci Rep. 10(1):4431. doi:https://doi.org/10.1038/s41598-020-61193-x.
- Gustamansyah. 2019. Laporan Pelaksanaan Kegiatan Monitoring Kantong Semar Kelam (Nepenthes clipeata Danser) di TWA Gunung Kelam. Indonesia: Balai Konservasi Sumber Daya Alam, Pontianak, Kalimantan Barat.
- Hallier H. 1895. Ein neues Cypripedium aus Borneo. Natuurkd Tijdschr Voor Ned Indië. 54:450–452.
- Heon SP, Clarke CM. 2015. Prey capture patterns in Nepenthes species and natural hybrids — are the pitchers of hybrids as effective at trapping prey as those of their parents?. Carniv Plant Newsl. 44:62–79.
- Hultine KR, Grady KC, Wood TE, Shuster SM, Stella JC, Whitham TG. 2016. Climate change perils for dioecious plant species. Nature Plants. 2(8):16109. doi:https://doi.org/10.1038/nplants.2016.109.
- Karl I, Bauer U. 2020. Inside the trap: biology and behavior of the pitcher-dwelling crab spider, Misumenops nepenthicola. Plants, People, Planet. 2(4):290–293. doi:https://doi.org/10.1002/ppp3.10104.
- Lam WN, Lai HR, Lee CC, Tan HTW. 2018. Evidence for pitcher trait-mediated coexistence between sympatric Nepenthes pitcher plant species across geographical scales. Plant Ecol Divers. 11(3):283–294. doi:https://doi.org/10.1080/17550874.2018.1517831.
- Lee CC. 2009. A preliminary conservation assessment of Nepenthes clipeata (Nepenthaceae). In: Lee CC, Clarke C, editors. Proceedings of the 2007 Sarawak Nepenthes Summit; Kuching (Malaysia). Sarawak Forestry Corporation. p. 96–100.
- Lovadi I 2010. Conservation of critically endangered Nepenthes clipeata on Mt. Kelam, West Kalimantan, Indonesia. London (UK): The Rufford Foundation; [Accessed 2020 May 18]. https://www.rufford.org/files/01.09.09%20Final%20Report.pdf
- Manning DAC. 2010. Mineral sources of potassium for plant nutrition. A review. Agron Sustain Dev. 30(2):281–294. doi:https://doi.org/10.1051/agro/2009023.
- Mansur M. 2006. Nepenthes: kantong Semar yang Unik. Jakarta (Indonesia): Penebar Swadaya.
- Mansur M. 2017. Potensi serapan CO2 pada beberapa jenis kantong semar (Nepenthes spp.) dataran rendah. Berita Biologi. 16(1):47–57. doi:https://doi.org/10.14203/beritabiologi.v16i1.2269.
- Mansur M, Brearley FQ, Jakalalana S, Rachmat F. Gustamansyah. In Review. Preliminary plant inventory of Mount Kelam, West Kalimantan, Indonesia. Edinb J Bot.
- Moran JA, Gray LK, Clarke C, Chin L. 2013. Capture mechanism in Palaeotropical pitcher plants (Nepenthaceae) is constrained by climate. Ann Bot. 112(7):1279–1291. doi:https://doi.org/10.1093/aob/mct195.
- Morisita M. 1959. Measuring of the dispersion and analysis of distribution patterns. Mem Fac Sci Kyushu Univ Ser E Biol. 2:215–235.
- Munné-Bosch S. 2015. Sex ratios in dioecious plants in the framework of global change. Env Exp Bot. 109:99–102.
- Olde Venterink H, Wassen MJ, Verkroost AWM, de Ruiter PC. 2003. Species richness-productivity patterns differ between N-, P-, and K-limited wetlands. Ecology. 84(8):2191–2199. doi:https://doi.org/10.1890/01-0639.
- Osunkoya OO, Daud SD, Di-Giusto B, Wimmer FL, Holige TM. 2007. Construction costs and physico-chemical properties of the assimilatory organs of Nepenthes species in northern Borneo. Ann Bot. 99(5):895–906. doi:https://doi.org/10.1093/aob/mcm023.
- Osunkoya OO, Daud SD, Wimmer FL. 2008. Longevity, lignin content and construction cost of the assimilatory organs of Nepenthes species. Ann Bot. 102(5):845–853. doi:https://doi.org/10.1093/aob/mcn162.
- Pontianak Tribune . 2020. Aksi pencurian tumbuhan langka kantong semar dilakukan sejak tahun 2017, 1 pokok dijual Rp 500 ribu. [Accessed 2021 Feb 22].https://pontianak.tribunnews.com/2020/05/28/aksi-pencurian-tumbuhan-langka-kantong-semar-dilakukan-sejak-tahun-2017-1-pokok-dijual-rp-500-ribu
- Porder S, Ramachandran S. 2013. The phosphorus concentration of common rocks—a potential driver of ecosystem P status. Plant Soil. 367(1–2):41–55. doi:https://doi.org/10.1007/s11104-012-1490-2.
- Rembold K, Fischer E, Striffler BF, Barthlott W. 2012. Crab spider association with the Malagasy pitcher plant Nepenthes madagascariensis. Afr J Ecol. 51(1):188–191. doi:https://doi.org/10.1111/aje.12037.
- Sellan G, Thompson J, Majalap N, Brearley FQ. in revision. Influence of species functional strategy on leaf stoichiometric responses to fertilization in a Bornean heath forest. J Ecol.
- Simpson RB.1995. Nepenthes and conservation. Curtis’s Bot Mag. 12:111–118.
- Thorogood CJ, Bauer U, Hiscock SJ. 2018. Convergent and divergent evolution in carnivorous pitcher plant traps. New Phytol. 217(3):1035–1041. doi:https://doi.org/10.1111/nph.14879.
- Trethowan LA, Blonder B, Kintamani E, Girmansyah D, Utteridge TMA, Brearley FQ. 2021. Metal-rich soils influence tropical tree stoichiometric distinctiveness. Plant Soil. 461(1–2):579–589. doi:https://doi.org/10.1007/s11104-021-04839-7.
- van der Ent A, Sumail S, Clarke CM. 2015. Habitat differentiation of obligate ultramafic Nepenthes endemic to Mount Kinabalu and Mount Tambuyukon (Sabah, Malaysia). Plant Ecol. 216(6):789–807. doi:https://doi.org/10.1007/s11258-015-0468-6.
- White PJ, Broadley MR. 2003. Calcium in plants. Ann Bot. 92(4):487–511. doi:https://doi.org/10.1093/aob/mcg164.