
Abstract
Marismeña is a feral Spanish cattle breed reared in the Doñana National Park, whose meat quality is traditionally appreciated in the area of influence. We assessed male (n = 32) and female (n = 22) Marismeña calves raised in their natural habitat (n = 30) vs. feedlot conditions (n = 24) to test differences in the lipid profiles of the m. longissimus thoracis. Meat from the feral system displayed (p < .05) lower intramuscular fat (IMF) (0.93%), higher saturated fatty acids (SFA) (18:0) proportion, and lower monounsaturated fatty acids (MUFA) proportion compared with feedlot meat. Polyunsaturated fatty acids (PUFA) percentage was higher for feral meat compared with feedlot meat, especially the proportion of PUFA n-3, which was about 8.5 times greater. The PUFA/SFA (0.37) and PUFA n-6/n-3 (2.87) ratios showed that feral meat was presumably healthier, from a human dietary perspective, than feedlot meat (0.21 and 13.42, respectively). This study used a direct comparison of natural feral meat and conventional feedlot meat in cattle, and the results demonstrated a clear influence of natural production on the constituents of meat.
Introduction
Marismeña is a breed of cattle raised under feral conditions in the National Park of Doñana, which is a natural reserve located in southern Spain. Almost 95% of the individuals belonging to the breed live there, in an area comprising about 100,000 ha. The breed has a long historical tradition, with references to it dating back to the thirteenth century (Calderón Citation2008), but the present census of 2204 animals indicates that the Marismeña is an endangered breed. Marismeña cattle raised in Doñana Park are fed exclusively on natural pastures with almost no human interference, such that they are only herded together for official health check controls and for animal identification once a year (Lazo Citation1995). Natural mating takes place in the wild, and Marismeña calves are born under feral conditions, receiving maternal milk until they are naturally weaned. Occasionally, some young animals are bought by external meat managers, and fattened using a conventional feedlot system.
In recent years, increased awareness about the effect of ingestion of saturated fats in diet and its effect on human health has promoted a negative perception by consumers towards some animal products, such as beef (Daley et al. Citation2010). This is thought to be a consequence of the low levels of PUFA and high levels of some hypercholesterolemic SFA in beef, which raises total and low-density lipoprotein (LDL) cholesterol after ingestion (Scollan et al. Citation2006). However, the lipid profiles of cattle show variability, depending upon factors such as diet, breed, sex and age (Scollan et al. Citation2014; Sevane et al. Citation2014). In general, meat produced by animals under extensive conditions is leaner, with the IMF being rich in PUFA n-3, conjugated linoleic acid (CLA), and t11-18:1 fatty acids (FAs), which are related to the maintenance of health (Daley et al. Citation2010). The differences among breeds seem to be linked/associated to differences in the genome and expression of proteins, which intervene in the quantity and kinds of fat deposition (Shingfield et al. Citation2013).
Meat and meat products originating from different types of production systems are receiving increased scrutiny by consumers, especially when they are produced under circumstances that are perceived to be associated with animal welfare. The special conditions under which Marismeña cattle are produced may represent an important added value, thus contributing towards the conservation of the breed. In these terms, this study was conducted to characterise lipid profiles of male and female Marismeña cattle, reared in the wild (feral system; FS) or in captivity under conventional intensive conditions (IS).
Materials and methods
Sampling was adapted to take into account the special circumstances of this feral breed management, and recognising the fact that 95% of the animals belonging to the breed lives in the National Park of Doñana under feral conditions. This area is mostly marshland for the greater part of the year, and animal management is performed only from midsummer until the end of the summer. At this time, all the animals are herded together for sanitary checks and for the identification of new-born calves.
Animal sampling and rearing systems
For this study, 54 Marismeña calves, born and raised under feral conditions in Doñana National Park during the spring and with an estimated age of 147.31 ± 34.27 days and a mean live weight of 130.62 ± 45.64 kg, were random and proportionally chosen from the total number of male and female calves caught during the management season in the summer. These animals were assumed to be representative of the whole population raised in the Park, and were separated into two groups: the first group comprised 30 animals (10 females and 20 uncastrated males) which were marked and reintroduced into FS, and another group of 24 animals (12 females and 12 uncastrated males) which were moved to a nearby experimental feedlot where they were raised intensively (IS) with grain supplementation, during 14 months. Sampling was proportional to the total number of females and males caught in the season, which is unbalanced because more females are kept for replacement than males.
Animals raised in the Park under feral conditions had continuous access to native pastures, being Anthoxanthum ovatum, Vulpia membranacea, Chaetopogon fasciculatus, Anthemis cotufa, Tolpis barbata, Ornithopus sativus, Cynodon dactylon, Panicum repens, Juncus maritimus, Eleocharis palustris and Cressa cretica the most abundant and consumed species in the pasture (Lazo & Soriguer Citation1993). The IS group of animals received, in addition to alfalfa hay supplied ad libitum, a concentrate supplement composed of 39.8% corn, 17.8% barley, 12.1% soybean meal, 6.6% DDGS, 6.4% wheat bran, 6% gluten feed, 3% soybean hulls, 2.7% sorghum, and 5.4% vitamins and minerals. The amount of feed supplied was adjusted with live weight, such that, over the finishing period considered, animals received an average of 7.5 kg of concentrate per day, administered twice daily. The chemical composition and fatty acid profile of the finishing diet used in the feedlot are presented in Table .
Table 1. Chemical composition of concentrate supplied in intensive system and fatty acid profile (in % of total fatty acids) of finishing diet (concentrate and alfalfa hay).
Sample collection
Given the absence of systematic management, experimental animals were very heterogeneous. They were slaughtered at an age of 552.40 ± 34.65 days, at a live weight of 329.86 ± 99.32 kg, in a slaughter plant, following EU regulations on the protection of animals during transport and related operations, and at the time of killing. Meat samples from each carcase were collected from the m. longissimus thoracis between the fifth and seventh rib, at 24 h post mortem. Samples were individually packaged, identified, fast-frozen, and stored at −20 °C until required for further analysis, at which time they were placed at 4 °C for thawing. The samples were trimmed to remove visible adipose and connective tissues, and minced in a commercial mixer/blender until total homogenisation of the sample was achieved.
Laboratory analysis
Chemical analyses were carried out following AOAC recommendations (AOAC Citation2005) to determine the proximate composition of meat, including moisture and IMF. To determinate IMF, duplicate samples of fresh meat were boiled with a 4 N HCl solution and subsequently washed and filtered to obtain a residue on which a Soxhlet extraction was performed using petroleum ether as solvent. In all cases, the results were expressed in percentage of the meat sample. For the analyses of FAs, total lipids were extracted from 0.5 g of freeze-dried sample, using chloroform/methanol (2/1; v/v) as described by Folch et al. (Citation1957). The preparation of FA methyl esters (FAMEs) was carried out as described by Raes et al. (Citation2001), using NaOH/MeOH and HCl/MeOH. The FAMEs were submitted to gas–liquid chromatography (GLC) on an Agilent HP 6890 chromatograph (Agilent Technologies Inc., Palo Alto, CA) equipped with a flame ionisation detector, using a split injector ratio of 1:10 and a 100 m capillary column (100 m × 0.25 mm internal diameter ×0.20 μm film thickness; CP-Sil 88; Varian Inc., Walnut Creek, CA). The temperature and other chromatograph settings were as follows: initial column temperature of 100 °C for 1 min, which was increased to 150 °C at 1 °C/min and maintained for 20 min; increased by 1 °C/min up to 190 °C and maintained for 5 min; and finally, increased by 1 °C/min up to 200 °C and maintained for 35 min. The temperature was kept at 250 °C for the injector and 280 °C for the detector. Helium was the carrier gas, at a constant pressure of 32.78 psi. FAs were identified by comparison of retention times with those of a FAME standard mix (Supelco37 standard FAME Mix, Supelco Inc., Bellefonte, PA), and quantification was obtained by normalisation. The FA composition was expressed both as a proportion of total FA identified and as mg/100 g of fresh muscle.
Data processing and statistical analysis
Statistical analyses were carried out by analysis of variance (ANOVA) of chemical composition variables, FA composition, and groups and ratios of FAs, using the General Lineal Model procedure of the SAS software version 9.0 (SAS Citation2002). In all cases, the animal was considered to be the experimental unit, and two-way ANOVA was performed, including the fixed effects of sex and management system, together with their interaction. In addition, age at slaughter was included as a covariate, to account for the differences between individuals resulting from their random selection from the herds. Regression functions of MUFA, PUFA, and SFA with respect to IMF were obtained using the REG procedure of SAS software.
Results
IMF and moisture content
The means and results obtained using the two-way ANOVA analysis for IMF and moisture content are presented in Table. Statistically significant differences were found for both factors: management system and sex. The R2 was high (>.70), demonstrating high efficiency for the model. Overall, the contents of the two meats differed substantially, with the FS meat having a lower IMF, and a higher moisture content than the IS meat. In addition, meat from males showed a lower IMF and a higher moisture content, compared with that from females.
Table 2. IMF and moisture content (g/100 g of fresh muscle) in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
Saturated fatty acids
Compared with IS meat, FS meat had approximately 6% higher proportion of SFA (p < .01) (Table ). All individual SFA were affected by the production system (p < .05), except for the proportions of 14:0, 16:0, 17:0 anteiso, and 22:0. In particular, FS had 5% higher proportions of 18:0. Although the SFA means were similar between the sexes, meat from females had higher relative values of 14:0 and 16:0 than meat from males.
Table 3. SFA composition (% of total identified fatty acids) in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
When the comparison was based on the amount of SFA (Table ), it was nearly twice higher in females than in males and in animals finished intensively than in the feral group. These differences were mostly a consequence of increased levels of 16:0 and 18:0 in females and in the group finished intensively.
Table 4. Fatty acid composition (mg/100 g of fresh muscle) in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
Monounsaturated fatty acids
More than half of the identified MUFA were affected by the production system (Table ), such that IS meat had higher proportions of total MUFA and cis-MUFA (nearly 1.4 times more than FS), and higher mean proportions of 18:1 c-9, 16:1 c-9, 20:1 c-11, 18:1 c-13, 18:1 c-11, and 18:1 t-9. In contrast, FS meat had higher relative values of 16:1 c-7, 16:1 t-9, and 18:1 t-11 (1.5 times higher). Higher proportions of MUFA and cis-MUFA were also found in females, due to the larger percentages of 18:1 c-15, 18:1 c-13, 16:1 c-9, and 18:1 c-9 in their meat. No differences were observed for proportion of trans-MUFA between sexes.
Table 5. MUFA composition (% of total identified fatty acids) in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
When the comparison is based on the amount of FA (Table ), animals in IS had nearly four times more total MUFA than those in FS, and females had twice the amount found in male. This was mostly due to increased amounts of 18:1 c-9 and 16:1 c-9 in IS and in females, while for the rest of MUFA the differences were smaller.
Polyunsaturated fatty acids
The proportion of total PUFA was higher for FS animals, being almost twice that observed in IS animals (Table ). The Relative value of PUFA n-6 was 1.5 times greater and PUFA n-3 was 8.5 times greater in FS than in IS meat. Overall, all individual proportions of PUFA were affected by the production system (p < .05), with the exception of 18:2 c-9, t-11, and 22:5 n-6. The 18:3 n-6, 20:5 n-3 (EPA), 22:5 n-3, and 22:6 n-3 (DHA) had proportions which were six times higher and 18:2 n-6 was 50% higher in FS than in IS. Males showed greater proportions of PUFA, PUFA n-6, and PUFA n-3 than females.
Table 6. PUFA composition (% of total identified fatty acids) in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
When the amount of FA in meat was considered (Table ), the total PUFA, n-6 and n-3 differed between finishing systems, with IS having higher means for total PUFA and n-6 and lower means for n-3. However, the various individual PUFA differed when compared among finishing systems, such that 18:2 n-6, 20:2 n-6, 20:4 n-6 and 22:4 n-6 were higher (p < .05) in IS when compared with FS, and the opposite pattern was observed for 18:3 n-3, 20:5 n-3, 22:5 n-3 and 20:6 n-3. Only total n-3 differed between sexes, with higher means in females (p < .05), but differences in individual FA were minor.
Ratios and indexes
Estimated means for the PUFA/SFA and n-6/n-3 PUFA ratios are shown in Table . In the two cases, both the finishing system and sex effects, as well as their interaction, gave significant results. The FS meat had higher PUFA/SFA levels than in IS, whereas the opposite was true for n-6/n-3. Males showed lower PUFA/SFA and n-6/n-3 ratios than females. Desaturation indexes for 16 and 18 were significantly affected by both system and sex, being greater for IS and females.
Table 7. FAs ratios and indices in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
Significant interactions between system and sex effects
The production system influenced 39 of the 55 studied variables, whereas the effect of sex influenced 24 variables. The variables not affected by production system were: 14:0, 16:0, 17:0 anteiso, 22:0, 14:1 c-9, 17:1 c-9, 18:1 c-12, 18:2 c-15, 18:1 t-6, t-7, t-8, 18:1 t-10, 18:1 t-12, 18:2 c-9, t-11 (CLA), 22:5 n-6, MUFA trans, and elongase (16 to 18).
The interaction of both factors was not important, but it was significant in the following variables: IMF content, moisture content, 16:0, 18:0, 18:1 c-11, 20:2 n-6, 18:3 n-3, 20:5 n-3, 22:5 n-3, SFA, PUFA, PUFA n-3, PUFA/SFA ratio, and n-6/n-3 ratio (Table ). Chemical composition of the m. longissimus thoracis showed that although both males and females in the FS system gave similar results for moisture and IMF, IS males had lower amounts of IMF and higher amounts of moisture when compared with IS females (4.7 times), while IMF in IS males was almost half that of IS females. Significant interactions were found for SFA and PUFA such that animals raised in FS had higher percentages of SFA (p < .05), which were also higher in females than in males, due to the higher proportion of 18:0 in those groups. However, a low percentage of 16:0 was found for FS males compared with other groups. Elevated proportions of PUFA (p < .05) were observed in meat from FS males, and these were 2–3 times higher than for the other combinations. This resulted in greater percentages of PUFA n-3 (18:3 n-3; 20:5 n-3, 22:5 n-3) and 20:2 n-6. Compared with males from FS, females showed nearly half the level of PUFA n-3.
Table 8. IMF and moisture content, individuals FAs proportions, summations and indices resulted from the interaction of factors System x Sex in m. longissimus thoracis of female and male Marismeña cattle finished under feral or intensive systems.
Relationship between proportions of fatty acid groups and intramuscular fat content
The correlation between IMF content and total SFA results showed a different behaviour for FS and for IS meats (Figure ). For FS meat there was a significant slope of SFA on IMF, whereas for IS meat the slope was nearly horizontal and the sum of SFA proportions was almost constant, even if the total fat increased. In this case, the behaviour of the slopes was mostly defined by the individual percentages of C16:0 and C18:0.
Figure 1. Relationships between intramuscular fat content (IMF) and percentage of (a) SFA, (b) MUFA, (c) PUFA, (d) PUFA n-6 and (e) PUFA n-3; of the longissimus thoracis muscle of Marismeña cattle finished under feral (•) or intensive (▴) systems.
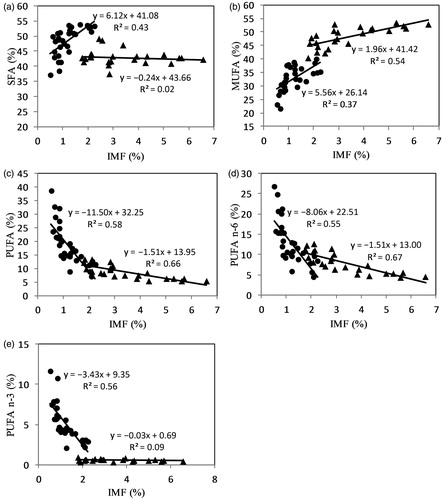
The behaviour of total MUFA proportions for regression on IMF in the IS and FS systems was distinct (Figure ). This total relationship was mostly made up of a combination of MUFA c-9 or C18:1 c-9 percentage, which were higher in IS animals. Overall, the trends of the curves for total SFA and total MUFA summations were opposing, and showed reciprocity when considering the feeding system.
With respect to PUFA, the curves for proportions of total PUFA, PUFA n-6, and PUFA n-3 showed inverse behaviour (Figure ) compared with the SFA and MUFA curves (Figure ). In these cases, when the IMF increased, the total PUFA and PUFA n-6 summations decreased in FS meat, and the same occurred for IS meat, but in a smoother way. The behaviour of PUFA n-3 was similar to that of n-6 for FS meat but not for IS meat, for which the slope of the PUFA n-3 was almost null, independently of the IMF content.
Discussion
To our knowledge, this is the first time that the lipid composition of beef cattle produced in feral conditions, without any human management, is compared with meat of the same genetic origin produced in conventional feedlot systems, which is a unique opportunity to test these two different scenarios.
Even though our interest was mostly in assessing the lipid profiles of meat produced in both systems, we first analysed their chemical composition, especially in terms of moisture and IMF, in order to test if the total amount of IMF affects the proportional lipid profile of meat. Our results for chemical composition were similar to values reported for other local Mediterranean breeds from Spain, Italy, and Portugal, but were lower than for non-native selected breeds reared in the same countries (Brugiapaglia et al. Citation2014; Costa et al. Citation2012; Indurain et al. Citation2010). Animals reared under intensive conditions showed a strong tendency towards higher fat content compared with feral animals, due to the higher nutritional value of concentrate feed (Wood et al. Citation2008). Additionally, despite the fact that females generally tend to deposit more fat than males, in our study differences in IMF were not found between sexes under feral conditions, probably due to the extremely low energy level of grass to which animals had access to. However, our results disagree with those from other researchers such as Daza et al. (Citation2014), who found differences between sexes in animals fattened under free-range conditions, with lower values of IMF for uncastrated males. However, free-range conditions are not the same as feral conditions, as the latter are almost completely natural without any human management.
It is well known (Wood et al. Citation2008) that fat content of meat has a fundamental effect on its FA composition. In our study, the relationship between the proportions of different FA groups and total IMF showed a diverse behaviour in the two management systems (Figure )). Our findings demonstrate that, when total fat increased, SFA proportion increased significantly in FS and remained nearly constant in IS animals, showing a different behaviour from what has been reported in the literature (De Smet et al. Citation2004). The proportion of MUFA increased with total fat in both FS and IS, but more pronouncedly in FS meat, indicating that an increase in the amount of fat, especially in FS calves, will produce an increase in the proportion of MUFA in their meat. The percentages of total PUFA and n-6 PUFA decreased as IMF increased, especially in FS animals, while the decline was very moderate in IS animals. On the other hand, in IS animals the PUFA n-3 remained nearly constant when IMF increased from 2 to 7%, so that the n-6/n-3 ratio decreased.
In our study, differences among sexes in the proportion of different fatty acids did not appear to be important, but a certain degree of interaction between system and sex was shown in our results.
Overall, meat produced by feral Marismeña cattle and by intensively finished animals differed substantially in the proportion of the various fatty acids, with FS meat having almost 6% more SFA than IS meat. Our findings do not agree with reports from other authors who assessed extensive vs. intensively produced meats, where the pattern generally found is that the proportion of SFA in meats produced extensively and semi-extensively is lower than in meats produced intensively (Alfaia et al. Citation2009; Bressan et al. Citation2011; Humada et al. Citation2012). In the present study, only four SFA had relative proportions not showing significant differences between the two finishing systems (14:0, 16:0, 17:0 anteiso, and 22:0). The other SFA proportions were significantly higher in the FS meat, which could indicate that this meat is hypercholesterolemic. However, according to Ulbricht and Southgate (Citation1991), the most harmful SFA are 12:0, 16:0, and 14:0, with the last one being the unhealthiest, showing a hypercholesterolemic power four times greater than the other two. In the current study, there was no difference in 14:0 proportions between the two types of meat, and the 16:0 percentage was higher in the IS meat. These findings, together with the fact that 18:0 is not hypercholesterolemic (Daley et al. Citation2010; Mensink Citation2005) and given that it was found at higher rates in feral system meat, suggest that, in general, FS meat has a healthier SFA profile.
The general FA profile of meats in our study indicated that IS showed higher MUFA proportions (approximately 30% more), agreeing with the results reported by Daley et al. (Citation2010), who found that grain-fed beef consistently produced higher proportions of this FA group. However, Alfaia et al. (Citation2009) found similar results between feedlot and pasture grazing. In our study, MUFA differences between systems occurred with cis-MUFA rather than trans-MUFA, such that for the latter no significant differences were found, contradicting the assertion that pasture-based diets result in higher percentages of trans-MUFA (Scollan et al. Citation2014). This is supported by our results for the individual MUFA percentages, as all 18:1-trans isomers were barely or not at all significantly different between systems, while the proportion of 18:1 c-9 was largely responsible for the differences between finishing systems, because of its prevalence when compared with the other FAs of this group. Our findings do not fully agree with the results reported by Bressan et al. (Citation2011), who found proportions of MUFA to be equal or slightly higher with grass feeding, while the differences between the two systems in the relative values of 18:1 c-9 were not significant. However, other reports are in agreement with our findings in Mediterranean local breeds (Costa et al. Citation2012; Humada et al. Citation2012), and the results of Warren et al. (Citation2008) could suggest a good explanation for this diversity. These authors found that MUFA proportions were higher in young (14 month-old) Aberdeen Angus and Holstein Friesian calves fed on forage compared with calves fed on concentrate, but when the animals were older (18 months old) this pattern changed, with the animals fed on concentrate showing higher proportions of MUFA, and this situation stabilised for both breeds when the animals were 24 months old. The results found in our study were consistent with this pattern, as slaughtering of Marismeña males occurred at around 20 months of age, and the proportion of MUFA was lower in FS than in IS animals. The major proportions of MUFA, mostly oleic acid, in IS may be due to the higher energy content of the diet and to increased steaoryl-coenzyme A desaturase activity (Daniel et al. Citation2004). In our study, this activity was different between sexes, with females showing higher activity, in agreement with the results by Daza et al. (Citation2014). From recent studies, it is difficult to judge whether the proportional differences in MUFA found in Marismeña IMF are positive or negative, because it is known that MUFA can reduce LDL and total/HDL ratio when substituting those SFA considered hypercholesterolemic (FAO Citation2010) but, on the other hand, studies in species such as primates and mice have shown that MUFA is not better than SFA in protecting against coronary artery atherosclerosis (Baum et al. Citation2012).
Regarding PUFA, our findings demonstrated important proportional differences between sexes and management systems, with very few interactions between these two factors. There was a general trend towards higher PUFA percentages in meats originating from males and FS, both for total PUFA, and for PUFA n-3 and PUFA n-6 separately. The PUFA group contains the most important FAs in terms of prevention of several illnesses, as there is evidence that linoleic (18:2 n-6) and alpha-linolenic (18:3 n-6) acids are essential for humans, and that the risk of coronary heart disease is decreased when PUFA replaces SFA (FAO Citation2010). The greatest bioactivity is seen for the long chain PUFA n-3, such as eicosapentaenoic acid (EPA) (20:5 n-3), docosapentaenoic acid (DPA) (22:5 n-3), and docosahexaenoic acid (DHA) (22:5 n-3), which have recognised beneficial effects on the prevention of heart attack, depression, and cancer (Daley et al. Citation2010; McAfee et al. Citation2010). Alpha-linolenic acid is not as bioactive as the longer chain of n-3 FAs, but it is important because it is the major PUFA that occurs in the green tissues of plants present in grasslands, and in ruminants this FA is the precursor, after elongation and desaturation processes, of some of the most active FAs (Decker & Park Citation2010). In our study, the meat of feral Marismeña cattle had 2–4 times the proportion of PUFA when compared with intensively reared animals. These findings agree with those reported by Bressan et al. (Citation2011) in Bos taurus and Bos indicus, in which animals finished on pasture showed higher proportions of PUFA than intensively reared animals, but the differences between the two systems reported by these authors were much lower than in our study. Costa et al. (Citation2012) compared meats from animals from two local Portuguese breeds fed on low-forage and high-forage diets, and found conflicting results; in some cases, the animals on low-forage diets had higher PUFA percentages than those on high-forage diets, but they also found the reverse in other cases. This could be due to the low consumption of fresh grass by all animals, as preserved forage does not have the same positive effects as grass. Humada et al. (Citation2012) presented similar results to ours when they compared meats obtained in semi-extensive and intensive systems from a local Spanish breed at two different slaughter ages. However, their results were much more balanced between finishing systems when compared with ours, with the exception of PUFAn-3 proportions, which showed greater differences. In our study, the proportions of individual PUFA were clearly higher in FS meat, with the exception of CLA and 22:5 n-6 (no differences), and 22:4 n-6 (lower content). These findings correlated with a low level of IMF and therefore a high concentration of phospholipids in the case of PUFA n-6, whereas the individual PUFA n-3 were associated with a higher intake of α-linolenic acid from pasture (Humada et al. Citation2012). In the case of 22:4 n-6, similar results were observed by Nuernberg et al. (Citation2005) when comparing grass-based and concentrate feeding systems. In our study, males showed higher percentages of all n-3 and some PUFA n-6 compared with females, mostly due to the lower amount of IMF. There was also a clear differentiation when the interaction between system and sex was assessed, with noticeable sex differences in FS.
Particular mention should be made of CLA, which is a collective term covering several conjugated isomers of linoleic acid, such as rumenic acid (18:2 c-9, t-11) and 18:1 t-10, c-11. This group is associated with prevention of inflammation, cancer, atherosclerosis, obesity, diabetes, and osteoporosis, among other benefits to human health (Decker & Park Citation2010; Dugan et al. Citation2011). CLA content in animal products (meat and milk) is strongly influenced by the consumption of fresh pasture (Kelly et al. Citation1998). However, no differences between system or sex were observed for 18:2 c-9, t-11 CLA percentages in our study, although some authors (Daley et al. Citation2010) have found such differences. We did find slight differences between systems for the proportion of the precursor 18:1 t-11, with FS meat having higher levels.
The goal of our work was to compare the impact of rearing system and sex on the relative importance of different fatty acids in Marismeña cattle, as has often been reported for many other breeds (Daza et al. Citation2014; Scerra et al. Citation2014; Bressan et al. Citation2016). However, it may also be of interest to assess the specific amount of the various fatty acids that are present in 100 g of fresh meat, which reflects the ingestion of fatty acids by consumers, and is highly dependent on the amount of IMF. In our study, IMF was nearly 1.7 higher in IS when compared with FS, such that, largely because of this, meat produced by cattle raised under IS had about 2.4 times more SFA, 4.2 times more MUFA and 1.4 times more PUFA than meat from FS. The pattern for individual fatty acids was mostly similar to the one found for the group that they belong to, but a few particular cases deserve to be mentioned. For example, the amount of CLA in 100 g of meat was nearly 3 times higher in IS, while the total amount of n-3 fatty acids, EPA and DHA were higher in FS by about 2.8, 6.6 and 1.9 times, respectively, when compared with IS.
When meat produced by females was compared with that obtained from males, the level of IMF was nearly 1.7 times higher. Therefore, the pattern found for the amount of the various fatty acids and groups of fatty acids in the two sexes was very much in line with the comparisons described for FS and IS, with higher amounts of the various FA and groups of FA in females, even though the differences between sexes were smaller than those observed when finishing systems were compared.
The various ratios computed among fatty acids and groups of fatty acids were the same, either when calculated from percentages or from the amount of fatty acids. The PUFA/SFA ratio is an important way to evaluate the nutritional value of red meat, and it is recommended that values should be above 0.5 (Scollan et al. Citation2006) but, with some exceptions, values below 0.1 are typical in beef production (Scollan et al. Citation2014). In the current study, this ratio was significantly different for meats produced in the two rearing systems, and it was almost twice in FS (0.37) when compared with IS (0.21) meat. Taking into account the results of the interaction, FS males had a ratio of 0.49, very close to the recommended value of >0.50, probably because of their lower IMF (0.93%).
The n-6/n-3 PUFA ratio is particularly important because an appropriate balance in this ratio in cell membranes is related to a significant reduction in the risk of coronary heart disease (Aldai et al. Citation2006). However, despite the beneficial effects of PUFA n-6, this FA group is related, owing to a major susceptibility to lipid peroxidation, with the inflammatory process, and some studies have also shown that a high n-6/n-3 ratio increases cancer risk (Lawrence Citation2013). In addition, both linoleic acid and alpha-linolenic acid are essential FAs, and are precursors of other compounds by two different methods, but enzymes such as desaturase are involved in both, so an excess of linoleic acid can interfere in the metabolism of alpha-linolenic acid (Daley et al. Citation2010). In our study, the n-6/n-3 ratio in FS meat was almost five times more balanced than that of IS meat, an important finding. Our results are in line with those of Humada et al. (Citation2012), but not with those of Bressan et al. (Citation2011), who reported closer results between the two finishing systems. Our results for key individual FAs, such as alpha-linolenic acid, EPA and DHA point towards recommending FS as a way to improve the healthy profile of FA in meat.
Moving on the estimated enzymatic activities studied in the present research, FS meats showed a significantly lower enzymatic activity as regards to the 16 and 18 desaturation indexes. The differences may be related to higher serum insulin concentration in animals fed on grain (Daniel et al. Citation2004), which supports our findings about the influence of a diet rich in concentrate on the enzymatic desaturation activities affecting the 16:0 and 18:0 isomers. However, interactions between breed type, slaughtering age, and diet on desaturase activity have been previously reported in the literature (Chung et al. Citation2007).
Conclusions
The main purpose of the present study was to assess the differences in fatty acid profiles in meat from feral and intensively reared Marismeña cattle. Our results indicate that meats obtained in the two systems differ considerably, but sex also plays an important role. Animals raised under feral conditions produce leaner meat, with a FA profile which is generally better. Even though the global SFA profile did not appear to be healthier for feral meat, the study of individual SFA showed that the largest proportion of SFA in feral meat was composed of 18:0, rather than other FAs that are considered hypercholesterolemic. PUFA are the FAs that have the largest protective bioactivity, and in our study, compared with meat obtained under intensive conditions, the meat from feral animals had far higher percentages of these FAs, particularly the most representative PUFA, such as EPA, DPA and DHA, and their precursor alpha-linolenic acid. The PUFA/SFA and n-6/n-3 ratios showed a much better balance in meat obtained under feral than in the intensive conditions. Our findings could be used as an indicator of the healthy FA profile of Marismeña meat obtained under feral conditions, which could be used as a way to recognise its unique properties and in support of a certification programme yielding a premium price, thus contributing to the conservation of this breed.
Acknowledgements
This research was partially financed through grants provide by Junta de Andalucía for the breed conservation programme. The authors are grateful to the Doñana Biological Station belonging to the Spanish Council for Scientific Research (CSIC), specially to Dr. Juan Calderón and Fernando Ibañez. We also like to thank Asociación Nacional de Criadores de Ganado Marismeño and Hermanos Ferrosa for their support.
Geolocation information section: 37°02'29.5"N 6°27'00.4"W.
Disclosure statement
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of this article.
Funding
This research was partially financed through grants provide by Junta de Andalucía for the breed conservation programme. The authors are grateful to the Doñana Biological Station belonging to the Spanish Council for Scientific Research (CSIC), specially to Dr. Juan Calderón and Fernando Ibañez. We also like to thank Asociación Nacional de Criadores de Ganado Marismeño and Hermanos Ferrosa for their support.
References
- Aldai N, Murray BE, Oliván M, Martínez A, Troy DJ, Osoro K, Nájera AI. 2006. The influence of breed and mh-genotype on carcass conformation, meat physico-chemical characteristics, and the fatty acid profile of muscle from yearling bulls. Meat Sci. 72:486–495.
- Alfaia CPM, Alves SP, Martins SIV, Costa ASH, Fontes CMGA, Lemos JPC, Bessa RJB, Prates JAM. 2009. Effect of the feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem. 114:939–946.
- AOAC. 2005. Official Methods of analysis of the AOAC INTERNATIONAL. Association of Official Analytical Chemists Publ. 18th ed. Gaithersburg (MD), USA.
- Baum SJ, Kris-Etherton PM, Willett WC, Lichtenstein AH, Rudel LL, Maki KC, Whelan J, Ramsden CE, Block RC. 2012. Fatty acids in cardiovascular health and disease: a comprehensive update. J Clin Lipidol. 6:216–234.
- Bressan MC, Rodrigues EC, Paula ML, Ramos EM, Portugal PV, Silva JS, Bessa RB, Telo da Gama L. 2016. Differences in intramuscular fatty acid profiles among Bos indicus and crossbred Bos taurus × Bos indicus bulls finished on pasture or with concentrate feed in Brazil. Ital J Anim Sci. 15:10–21.
- Bressan MC, Rossato LV, Rodrigues EC, Alves SP, Bessa RJB, Ramos EM, Gama LT. 2011. Genotype x environment interactions for fatty acid profiles in Bos indicus and Bos taurus finished on pasture or grain. J Anim Sci. 89:221–232.
- Brugiapaglia A, Lussiana C, Destefanis G. 2014. Fatty acid profile and cholesterol content of beef at retail of piemontese, limousin and friesian breeds. Meat Sci. 96:568–573.
- Calderón J. 2008. La vaca mostrenca de Doñana. Madrid: ICONA (Organismo Autónomo de Parques Nacionales).
- Chung KY, Lunt DK, Kawachi H, Yano H, Smith SB. 2007. Lipogenesis and stearoyl-coa desaturase gene expression and enzyme activity in adipose tissue of short- and long-fed angus and wagyu steers fed corn- or hay-based diets. J Anim Sci. 85:380–387.
- Costa P, Lemos JP, Lopes PA, Alfaia CM, Costa ASH, Bessa RJB, Prates JAM. 2012. Effect of low- and high-forage diets on meat quality and fatty acid composition of alentejana and barrosã Beef breeds. Animal. 6:1187–1197.
- Daley CA, Abbott A, Doyle PS, Nader GA, Larson S. 2010. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr J. 9:10. doi: 10.1186/1475-2891-9-10.
- Daniel ZCTR, Wynn RJ, Salter AM, Buttery PJ. 2004. Differing effects of forage and concentrate diets on the oleic acid and conjugated linoleic acid content of sheep tissues: the role of stearoyl-coa desaturase1,2. J Anim Sci. 82:747–758.
- Daza A, Rey AI, Lopez-Carrasco C, Lopez-Bote CJ. 2014. Effect of gender on growth performance, carcass characteristics and meat and fat quality of calves of avileña-negra ibérica breed fattened under free-range conditions. Span J Agric Res. 12:683–693.
- De Smet S, Raes K, Demeyer D. 2004. Meat fatty acid composition as affected by fatness and genetic factors: a review. Anim Res. 53:81–98.
- Decker EA, Park Y. 2010. Healthier meat products as functional foods. Meat Sci. 86:49–55.
- Dugan M, Aldai N, Aalhus J, Rolland D, Kramer J. 2011. Review: trans-forming beef to provide healthier fatty acid profiles. Can J Anim Sci. 91:545–556.
- FAO. 2010. Fats and fatty acids in human nutrition. Report of an expert consultation. Food and Agriculture Organization of the United Nations. Geneva, Switzerland.
- Folch J, Lees M, Sloane Stanley GH. 1957. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 226:497–509.
- Humada MJ, Serrano E, Sañudo C, Rolland DC, Dugan MER. 2012. Production system and slaughter age effects on intramuscular fatty acids from young tudanca bulls. Meat Sci. 90:678–685.
- Indurain G, Beriain MJ, Sarries MV, Insausti K. 2010. Effect of weight at slaughter and breed on beef intramuscular lipid classes and fatty acid profile. Animal. 4:1771–1780.
- Kelly ML, Berry JR, Dwyer DA, Griinari JM, Chouinard PY, Van Amburgh ME, Bauman DE. 1998. Dietary fatty acid sources affect conjugated linoleic acid concentrations in milk from lactating dairy cows. J Nutr. 128:881–885.
- Lawrence GD. 2013. Dietary fats and health: dietary recommendations in the context of scientific evidence. Adv Nutr. 4:294–302.
- Lazo A, Soriguer RC. 1993. Size-biased foraging behaviour in feral cattle. Appl Anim Behav Sci. 36:99–110.
- Lazo A. 1995. Ranging behaviour of feral cattle (Bos taurus) in Doñana National Park, s.W. Spain. J Zool. 236:359–369.
- McAfee AJ, McSorley EM, Cuskelly GJ, Moss BW, Wallace JM, Bonham MP, Fearon AM. 2010. Red meat consumption: an overview of the risks and benefits. Meat Sci. 84:1–13.
- Mensink R. 2005. Effects of stearic acid on plasma lipid and lipoproteins in humans. Lipids. 40:1201–1205.
- Nuernberg K, Dannenberger D, Nuernberg G, Ender K, Voigt J, Scollan ND, Wood JD, Nute GR, Richardson RI. 2005. Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of longissimus muscle in different cattle breeds. Livest Prod Sci. 94:137–147.
- Pitchford WS, Deland MP, Siebert BD, Malau-Aduli AE, Bottema CD. 2002. Genetic variation in fatness and fatty acid composition of crossbred cattle. J Anim Sci. 80:2825–2832.
- Raes K, de Smet A S, Demeyer D. 2001. Effect of double-muscling in belgian blue young bulls on the intramuscular fatty acid composition with emphasis on conjugated linoleic acid and polyunsaturated fatty acids. Anim Sci. 73:253–260.
- SAS. 2002. User's guide: statistics, version, 9th ed. SAS Institute Inc, Cary (NC), USA.
- Scerra M, Foti F, Cilione C, Chies L, Scerra V, Caparra P. 2014. Influence of stall finishing of Podolian young bulls raised on pasture on fatty acid composition and oxidative status of meat. Ital J Anim Sci. 13:857–863.
- Scollan N, Hocquette J-F, Nuernberg K, Dannenberger D, Richardson I, Moloney A. 2006. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 74:17–33.
- Scollan ND, Dannenberger D, Nuernberg K, Richardson I, MacKintosh S, Hocquette J-F, Moloney AP. 2014. Enhancing the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 97:384–394.
- Sevane N, Nute G, Sañudo C, Cortes O, Cañon J, Williams JL, Dunner S. 2014. Muscle lipid composition in bulls from 15 European breeds. Livest Sci. 160:1–11.
- Shingfield KJ, Bonnet M, Scollan ND. 2013. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal. 7:132–162.
- Ulbricht TL, Southgate DA. 1991. Coronary heart disease: seven dietary factors. Lancet. 338:985–992.
- Warren HE, Scollan ND, Enser M, Hughes SI, Richardson RI, Wood JD. 2008. Effects of breed and a concentrate or grass silage diet on beef quality in cattle of 3 ages. I: animal performance, carcass quality and muscle fatty acid composition. Meat Sci. 78:256–269.
- Wood JD, Enser M, Fisher AV, Nute GR, Sheard PR, Richardson RI, Hughes SI, Whittington FM. 2008. Fat deposition, fatty acid composition and meat quality: a review. Meat Sci. 78:343–358.