
Abstract
The study was conducted to analyse the effect of sheep body weight (BW) at mating, mid-pregnancy, lambing, early lactation, mid-lactation and late lactation on milk yield and patterns of milk production. Also, the effects of environmental factors such as number of lambing (NL) and type of lambing (TL) on BW and milk production were analysed. A total of 52 multiparous East Friesian ewes from an experimental flock were used. Ewes were assigned to three different groups according to their BW at each productive stage: low (LBW), moderate (MBW) and high BW (HBW). Lactations were fitted using the mechanistic model described by Pollott. Total milk yield (TMY), peak yield (PY) and time at peak yield (TPY) were also calculated. HBW ewes had consistently higher TMY (p < .001) and PY (p < .05) values, than LBW and MBW in most of productive stage measured. There was a positive linear relationship (p < .05) between TMY and BW in all-productive stage, except at mid-gestation where the relationship was quadratic. HBW ewes weighted at mid-pregnancy showed the highest values of maximum secretion parameter (p = .04) of Pollott model, which could partially explain the better milk yield of HBW ewes. A significant effect of NL on BW (p = .007) and TMY (p = .007) was observed. The BW ewe’s in pregnancy and early lactation is a useful indicator at farm level to improve the milk yield performance in dairy sheep.
Introduction
The East Friesian (EF) is the most popular dairy sheep at worldwide; it has been introduced to many countries to promote the development of the local dairy sheep or the industry using pure breed flocks and to improve milk production of native sheep breeds through crossbreeding (Haenlein and Wendorff Citation2006). The introduction of EF has showed variable and inconsistent results. Several authors have referred to the fact that EF sheep not shown good milk yields under dry and hot conditions (Fahmy and Shrestha Citation2011) even without nutritional deficiencies; for instance, Gootwine and Goot (Citation1996) referred that EF sheep have low milk production under Mediterranean conditions (161 L/lactation) compared with the productions in their natural environment (550 L/lactation). Similar results have been referred by Angeles-Hernandez et al. (Citation2014) with 76 kg per lactation of EF under semi-arid conditions.
In specialised dairy sheep animals, like EF sheep, high priority is given to functions of pregnancy and milk production (Bauman and Currie Citation1980). Adequate nutrient availability during pregnancy and lactation is key point in the development of the mammary gland and milk synthesis. Neville et al. (Citation2013) reported that ewes showed a decrease of mammary growth during pregnancy when they were fed below of their requirements, this finding is in line with various early reports (Dimauro et al. Citation2011; Paten et al. Citation2014) which point out that cell differentiation, cell turnover and epithelial cell activity are affected by nutrition during pregnancy in adult sheep. Also, the availability of nutrients during lactation determines an adequate milk yield with a better persistence of lactation curve.
During the late pregnancy, the utilisation of nutrients is less efficient and the feed intake is limited by the compression of the rumen by the growing foetus, passage rate of the feed through the digestive tract (Haenlein and Wendorff Citation2006), the underdevelopment of the rumen papillae (Mayer et al. Citation1986) and physiological factors associated mainly with the presence and mobilisation of body reserves (Ingvartsen et al. Citation1999); furthermore, the metabolic reserves are very important to successful lactation.
Although all nutrients are mostly derived from feed intake, the nutrients stored in body tissues are fundamental in specific production stages such as late gestation and early lactation, mainly supporting adequate development and performance of mammary gland. Throughout the year, sheep often show large variation of body reserves, with higher weight at the end of pregnancy, the last part of lactation and the dry period, and with lower weight in the first part of lactation (Cannas Citation2004a).
The measuring and recording the body weight (BW) in each stage of the productive cycle of dairy sheep is very important to evaluate the body condition and corporal reserves in preparation to high demand nutrient stages like final gestation and early lactation. The body condition score (BCS) technique has been recognised as an available option to estimate body fat variations. However, these estimations are some subjective and the values of this scale can be different among breeds and stages of the productive cycle (Cannas Citation2004b), hence the measurement of BW in animals of the same breed and living on a group under the same management is a useful tool to estimate the mobilisation of reserves during productive stages. Previous research has also shown a relationship between live BW and milk yield within or between breeds (Hansen et al. Citation1999; van der Linden et al. Citation2009), and BW has been used in selection programmes in cows (Hansen et al. Citation1999) and sheep (Mavrogenis and Papachristoforou Citation2000).
The aim of the present study was to analyse the effect of sheep BW at mating, mid-pregnancy, lambing, early, mid- and late lactation on milk yield and patterns of milk production in a dairy sheep farm of Chilean Patagonia. Also, the effects of environmental factors such as number of lambing (NL) and type of lambing (TL) on live weights and milk production were analysed.
Materials and methods
Study area and animal management
The study sample consisted of 52 multiparous EF sheep from an experimental flock of the University of Magallanes, Puerto Natales, Chile. The farm is located in the Magallanes region with a mean annual temperature of 6.5 °C (min: −2.1 °C and max: 15.6 °C); the average monthly maximum temperatures occur in January and February (11 °C and 10.6 °C, respectively) and average monthly minimum temperatures in June and July (2.2 °C and 1.8 °C, respectively). Annual average rainfall is 512.4 mm, with the rainy season lasting from March to July, although there low rainfall occurs throughout the year (Butorovic Citation2015).
The nutritional flushing began 2 weeks prior to breeding and continued during the breeding period; the sheep were supplemented with 200 g/d of commercial concentrate (15% CP; 2.6 Mcal EM/kg DM). During the first 100 days of pregnancy, the sheep were fed under a grazing system on mixed swards of mainly rye grass, cocksfoot and white clover. During the last 50 days of pregnancy, the ewes were supplemented with 400 g/d as DM of alfalfa hay. The feed management of milked ewes was strip grazing plus alfalfa hay (400 g DM/d) and commercial concentrate (400 g DM/d).
The ewes’ oestrus was synchronised using intravaginal sponges (medroxyprogesterone acetate) at the second week of May; after 48 hours, the sponges were removed and teaser rams were introduced to the flock to detect oestrus. Ewes in oestrus were hand-mated; PM-AM method of breeding was used, ewes detected in oestrus in the afternoon were mated early the next morning and those detected in oestrus in the morning were mated in the afternoon of the same day. Fifteen days after teaser rams were placed with the ewes again to detect oestrus activity and ewes were hand-mated again. The lambing occurred between the 9th and 26th of October.
Measures of milk yield and live BW
Sheep were milked mechanically once daily. Lambs suckled their dams freely during the first month of age. Although in Chile there is not an official milk recording regulation of sheep milk production, the milk yield was recorded every four weeks (B4) according to ICAR rules (ICAR Citation2016) with the first milk recording in the day 34 ± 5.6 post-lambing, approximately; the average of the lactation length was around 148.9 ± 8.6 days. The total milk yield (TMY) was calculated using the Fleischmann’s method (Ruiz et al. Citation2000). Sheep were weighed using a digital scale in the morning prior to feeding; the weighing were carried out at mating (WM), mid-pregnancy (WS), 48 h after lambing (WL), early lactation (WEL), mid-lactation (WML) and late lactation (WLL). According with their BW in each productive stage, sheep were assigned to one of three groups: low (LBW, n = 16), moderate (MBW, n = 18) and high body weight (HBW, n = 18).
Statistical analysis
Data were analysed using the mixed model analysis (PROC MIXED, SAS Institute Citation2002) to repeated measures data structure. The mathematical model for the analysis of milk production and live BW, included fixed effects for month of lactation, TL (single, n = 16; twins, n = 31; and triplets, n = 5) and NL (1st, n= 5; 2nd, n = 17; 3rd, n = 15; 4th, n = 10 and 5th, n = 5); the ewe was included as a random factor. Post hoc pairwise comparison was carried out using the Tukey’s HSD test.
Akaike information criteria and Bayesian information criteria were used as indicators to select the most appropriate variance–covariance matrix, which was first-order autoregressive AR (1); the other covariance structures tested were Toeplitz, unstructured, compound symmetry and first-order ante-dependence ANTE (1). Significance was declared with p < .05, and trends were declared at p < .10.
Milk production and live BW of sheep in different stages of the productive cycle were taken as continuous variables. Higher-order polynomials were used to analyse their relationship (PROC REG, SAS Institute Citation2002). To analyse the effect of studied factors (NL, TL, WM, WS, WL, WEL and WLL) in lactation curves, we used the mechanistic model described by Pollott (Citation2000).(1)
Where: Mt=milk production; MS = maximum potential of secretion; GR = relative proliferation rate of secretory cells during early lactation; NOD = proportion of secretory cell at start of lactation; DR = relative decline in cell numbers as lactation progressed. The parameters of the Pollott model were estimated individually for each lactation curve using a nonlinear regression analysis (PROC NLIN, SAS 2002).
Results
Environmental factors that affect milk production
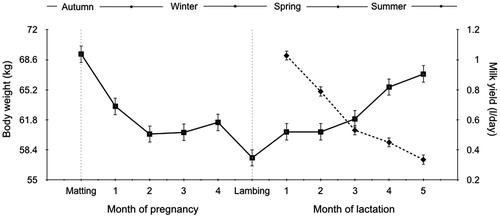
The effect of month of lactation, NL, TL on milk production and BW was analysed in the current work; the results are shown in Table . The milk production was affected significantly (p = .001) by the month of lactation with the highest yields in the first month. It had a gradual decrease until the fifth month of lactation. In contrast, the BW was significantly lower (p < .0001) in the first and second months and rose gradually with the higher values in the last month of lactation; BW decreases from mating to lambing (Figure ).
Table 1. Least square means of milk yield (MY) and live body weight (BW) in relation with month of lactation, number of lambing and type of lambing of East Friesian sheep.
Figure 1. Milk yield (dotted line) and body live weight (solid line) in pregnancy and lactation period of East Friesian sheep. The lactation month in both milk yield and live body weight had significant effect (p < .001).
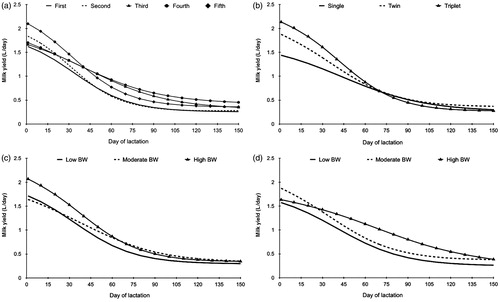
The NL significantly affected the TMY (p = .007); first and second lambing ewes had lower milk yields than ewes of other lambing. There was an effect of the NL on milk production patterns; all parities show an atypical shape of the lactation curve (continuously decreasing), with the fifth lambing ewes showing the highest milk yields on the 1st day (Figure ). Ewes of first and second lambing showed significant (p = .001) lower BW (55.62 and 56.48 kg, respectively); the higher BW was reported for ewes of third, fourth and fifth lambing (63.15, 62.36 and 65.22 kg, respectively) (Table ).
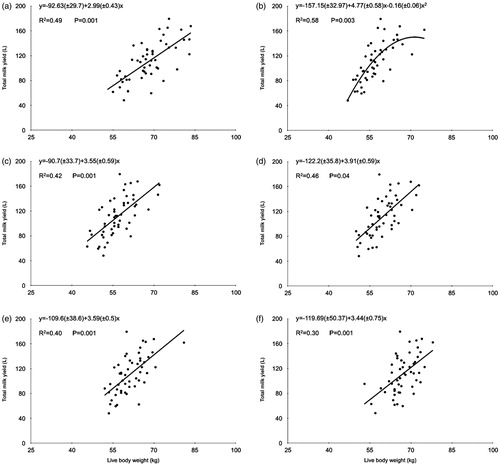
Figure 2. Relationships between total milk yield (TMY) and body live weight (BW) at mating (a), mid-pregnancy (b), lambing (c), early lactation (d), mid-lactation (e) and ending lactation (f) of East Friesian sheep, n = 52.
Litter size does not have an effect (p > .05) on milk production and BW of the studied sheep. A high number of animals might be needed to find significant effects of this factor. However, a trend is shown in TMY in analysed data (p = .09); ewes bearing twins and triplets had greater milk yields, by 6.9% and 32.7%, respectively, in comparison with single bearing ewes (Table ). Also, lactation curves show differences in the first weeks of lactation in relation with the ewe’s litter size (Figure ).
Figure 3. Lactation curves (L/d) according with the number of lambing (a), type of lambing (b) and live body weight (BW) at mating (c) and lambing (d) of East Friesian sheep. Lactation curves fitted using the Pollott (Citation2000) model; a, number of lambing; b, litter size; c, BW at mating; d, BW at lambing.
Effect of live body weight on milk production
Mean squares and test of significance of milk production and parameters of the Pollott (Citation2000) model in relation with the BW are presented in Table . The mean of TMY was 111.08 L with an average lactation length of 149 days. The BW in all analysed productive stages affected the TMY (p < .01). HBW ewes weighed at mating and mid-pregnancy showed the highest values (p < .001) of TMY followed by MLW and LBW ewes. MLW and HBW ewes had higher values (p < .01) of TMY than LBW when was weighed at lambing, early and mid-lactation. There was an effect of BW (p < .05) on the lactation length when the ewes were weighted at mating, mid-pregnancy, early and mid-lactation.
Table 2. Effect of live body weight of dairy sheep on milk production characteristics and parameters of Pollott model.
Maximum secretion was higher in HBW ewes in comparison with LBW and MBW ewes at mid-pregnancy. HBW ewes tended to have higher maximum secretion values at early lactation compared with LBW and MBW ewes (1.92 vs. 1.66 and 1.73 L; p = .09; for HBW vs. LBW and MBW-ewes, respectively). No dam-weight effect (p > .05) was found on GR and DR parameters; however, LBW ewes showed numerically higher rates of growth and death in most of measured stages. Ewes with LBW and MBW in most of productive stages had smaller peak yield (p < .05) than those with HBW; also, HBW ewes reached peak time before at mid-pregnancy (33.44 and 37.97 vs. 29.62 d; p = .04) and mid-lactation (36.89 and 37.66 vs. 29 d; p = .05; for LBW and MBW vs. HBW, respectively).
There was a positive linear relationship between TMY and BW at the most productive stages measured (Figure ). The TMY increased at a rate between 2.99 and 3.91 L of TMY per kilogram of increase in BW in all productive stages, except at mid-gestation. There was a quadratic relationship between TMY and BW at mid-gestation. The strength of the relationship between TMY and BW was least to BW at mating and lambing (R2 = 0.49 and 0.42, respectively) and highest to BW at mid-gestation (R2 = 0.58) in measures of BW before lactation.
Discussion
The EF sheep is recognised for its superior dairy performance; however, the ewes of current study had lower milk yields than reported by Fahmy and Shrestha (Citation2011) in their native environment (540–650 kg per lactation). Several reports indicate that the EF breed has a low ability to adapt to exotic climatic conditions, resulting in lower milk yields mainly under dry or hot conditions (Gootwine and Goot Citation1996; Angeles-Hernandez et al. Citation2014). Hunter et al. (Citation2015) report that the variability of response of the East Frisian breed and its crosses is maybe due to insufficient nutrition to maximise milk yield under grazing condition, as in the current study, or inadequate milking pressure; although the heat, humidity and health problems are environmental factors that have been strongly associated with the low and moderate milk performance of the EF genotype outside the native conditions (Gootwine and Goot Citation1996).
The analysed sheep showed an atypical shape of lactation curve (continuously decreasing) with the peak of milk production in the first milk yield record and gradually declining until the end of the trial. Similar patterns of lactation curve have been reported in the EF (Mckusick et al. Citation2001; Angeles-Hernandez et al. Citation2014) and other breeds (Carta et al. Citation2001). The atypical shape of lactation curves has been associated with meat and wool breeds (Godfrey et al. Citation1997) or low genetic merit of animals; however, dairy sheep breeds, like the EF, can also show atypical curves due to environmental factors and nutritional management that play a dominant role in the shape of the lactation curves. Also, the lack of milk yield records in the first stage of lactation is another likely cause of atypical curves in the current work, since the first milk yield was recorded ∼35 days post-lambing.
Environmental factors that affect milk production
Several previous studies (Gootwine and Pollott Citation2000; Angeles-Hernandez et al. Citation2013) reported a quadratic effect of the NL on milk production, point out that the lowest milk yields and more persistent lactation curves occur in the first lactation due to the mammary gland not being fully developed at the beginning of the first lactation (Abdelsayed et al. Citation2015); afterwards, milk yields increase with the NL, showing the highest milk yields in the third and fourth lactation and a consequent decline until culling; this is in agreement with the results of the current study. An early study analysed the effect of the NL in milk production and BW of Chios sheep, stating higher milk yields in third lambing and maximum BWs in the fifth parity (Mavrogenis and Papachristoforou Citation2000); again, that coincides with our results.
The BW decreased from mating to lambing in average by 11.75 kg. Similar findings have been reported by Louca et al. (Citation1974) in Chios sheep under a low plane of feeding (barley straw ad libitum). They showed a considerable weight loss and also lower milk yields (p < .05) than sheep under medium (0.5 kg/day of concentrate) and high plane (1 kg/day of concentrate) of feeding. Mavrogenis et al. (Citation1980), who expected similar BW changes to the present study during the pregnancy in sheep fed on stubble grazing and/or cereal straw during early and late pregnancy.
In the current study, the pregnancy period coincides with the coldest season and lowest availability and quality of pastures; hence, sheep did not cover their feed requirements and were forced to mobilise body reserves. The results were consistent with (Laporte-Broux et al. Citation2011) who had analysed the effect of restricted feeding at late pregnancy on dairy goats. They suggest that the metabolic status at the lactation can result in the redirection of nutrients toward the foetus and therefore dam BW gain at the end of the pregnancy may be physiologically difficult to obtain.
The sheep’s BW decreased by 4.01 kg from fourth month of pregnancy to lambing; the sheep was weighed one day after lambing; hence, the loss of BW can be due to the expulsion of lamb or lambs, amniotic fluid and placental mass (Sahlu et al. Citation1995) and the decline of feed intake during the days previous to lambing (Charismiadou et al. Citation2000). The increase of the ewes’ BW during lactation was similar to the weight gain observed by Godfrey et al. (Citation1997) and Hunter et al. (Citation2015). After the ewes lost weight in early lactation, they began to gain weight again during the first month of lactation approximately; in contrast, milk production declined over time.
Effect of live body weight on milk production
Pregnancy, mainly foetal and mammary gland development, induces adaptive responses in energetic metabolism and peripheral tissue mobilisation of glucose, which can be more pronounced under a regime of moderate-under nutrition. Dramatic metabolism changes also happened during the transition from late pregnancy to establishing lactation with the goal to meet the highest nutritional requirements of adult sheep without any increase of voluntary feed intake, this is a major motivation to analyse the effect of BW during the pregnancy and lactation on milk production in dairy animals.
The effect of BW measured in pregnancy and lactation on milk yields has been demonstrated in some previous studies (Berry et al. Citation2007). In the present experiment there was a positive relationship between TMY and BW in the all-productive stage; the finding of a greater milk yield in heavier ewes is in general agreement with previous studies (Berry et al. Citation2007; van der Linden et al. Citation2009). Ewes weighed at mid-lactation showed a quadratic relationship between TMY and BW which has been reported previously by Berry et al. (Citation2007) in dairy cows weighed pre-calving (8 weeks), at calving and during early lactation; the study mentions that heavier cows produce more milk, although the effect per kg of BW tended to diminish with an increase of BW.
In previous studies, the relationship between BW and milk yield showed inconsistent results. Several authors report an increase of milk yields as the BW increases (Sieber et al. Citation1988); they associated this with a greater energy intake and high levels of body reserves, that determine greater availability of precursors of milk synthesis reaching the mammary gland and higher udder volume in heavy ewes (Godfrey et al. Citation1997). Also, a positive genetic and phenotypic correlation between milk yields and BW have been reported (Veerkamp Citation1998; Veerkamp and Thompson Citation1999).
Hence, the breeding selection based on milk production can mean the selection of larger and heavier animals (Harville and Henderson Citation1966). Nevertheless, this situation apparently occurs only in the initial stages of selection programmes, since in efficiently selected genetic programmes no differences in BW between animals with different milk yields have been shown. This was observed in Lacaune sheep from a selection programme with duration of 50 years, in which the low production line versus the high productive line did not show differences in BW; however, the latter produced 22% more milk (Barillet et al. Citation2001).
Another successful genetic selection programme of dairy sheep has been carried out for the Awassi breed. Unimproved Awassi sheep under extensive conditions produce between 70 L and 107 L of milk per lactation (Dag et al. Citation2005). In contrast, improved Awassi sheep reach milk yields of up to 506 l/lactation as an average; with better milk yield of improved Awassi inclusive in comparative studies under the same conditions (Alqaisi Citation2007). Gootwine (Citation2011) carried out an intensive genetic improvement management programme; this researcher reported that high milk production of improved Awassi lines is associated with an increase of the ewe’s BW. The improved Awassi line sheep showed a higher BW (22 kg) in comparison with unimproved Awassi (Gootwine Citation2011).
In contrast, Mavrogenis and Papachristoforou (Citation2000) suggest that BW is not a determinative and is independent of the volume of milk yield; they found very low genetic (r = 0.08) and phenotypic (r = 0.08) correlation between milk production and BW. Also, Sieber et al. (Citation1988) reported that lighter cows tended to show larger milk yields than heavier cows. Berry et al. (Citation2007) suggest that contrasting results of the BW’s effect on milk yield could be associated with the characteristics of feeding management. Berry et al. (Citation2007) did not identify a significant association between BW and milk production when the sheep were fed with total mixed rations; in contrast to studies with grazing system as the main energy source. As the BW in pregnancy and lactation is significantly important, cows with better body conditions produce more milk; this finding is in agreement with the present study.
It has been suggested that the variability in the magnitude of relationships between milk yields and BW is in relation to the stage of pregnancy or lactation in which the BW measures were made, with higher genetic correlations between milk yields and BW at early lactation (r = 0.37) in comparison to the BW at ending lactation (r = 0.04) (Veerkamp and Thompson Citation1999). The different values of R2 in the present study confirm the above findings, as BW at mid-pregnancy explained the change of TMY better in comparison with BW at mid- and ending lactation. This finding could be explained by the fact that stored body fat is an important contributor to live BW during some parts of lactation, and body tissue mobilisation is closely related to milk yield (Veerkamp Citation1998). Also, Mavrogenis et al. (Citation1980) reported in three dairy sheep breeds that the correlation between TMY and BW changed from 0.28 at lambing to −0.23 in the week 18 of lactation. They conclude that adequate BW at lambing predisposes sheep to higher milk production.
Between and within breeds, sheep with higher BW in pregnancy showed better milk yields, maybe due to the high feed intake capacity and better body store reserves (Mavrogenis et al. Citation1980). The milk production depends on the potential milk production (number of active alveoli and milk secretion of each alveolus) and of the net energy available for lactation (Dimauro et al. Citation2011). In this context, adequate development of the mammary gland during pregnancy is a key factor to reach good milk production in the subsequent lactation. Adequate nutrient intake during late pregnancy is very important as the sheep’s mammary gland grows by 70% during the last 4 weeks of gestation (Swanson et al. Citation2008). Furthermore, the sheep’s mammary development is essentially complete by parturition, in contrast to the mammary gland development in rodents, pigs, some dairy cows and goats which continues during early lactation (Paten et al. Citation2015).
In the current study, there was a significant decrease of BW from mating to lambing, which can be associated with a maternal under nutrition during pregnancy. This low BW at lambing can be associated with the low quantity and quality of forage in the grassland during the pregnancy. During autumn and winter, temperatures are lowest and dry matter forage is least available in the region in which the study was conducted. Mellor and Murray (Citation1985) refer a significant reduction of udder measurements and milk yields in ewes with low nutrient intake during late pregnancy.
Also, a biological interpretation of the parameters as given in the Pollott model (Pollott Citation2000) can help to explain in part the positive effect of BW in milk production, as HBW ewes showed highest values of MS potential. These results concur with the findings in Gootwine and Pollott (Citation2000) for Awassi sheep. They stated that higher values of MS were associated with a greater number of secretory cells, a higher secretory rate per cell or both; Gootwine and Pollott (Citation2000) reported a significant positive correlation between MS and TMY (p < .001; r = 0.69) and PY (p < .001; r = 0.99). Swanson et al. (Citation2008) analysed the effect of nutrition level (restricted, control and high) on mammary development. Ewes with high nutrition level had the highest BW and showed high values of proliferation of mammary alveolar cells and weight of mammary gland.
In specialised dairy sheep breeds, the most common health problems (metabolic and infectious) occur close to lambing and in early lactation. This as consequence of that their elevated genetic potential of milk production determines high levels of energy requirements at ending of pregnancy, unfortunately also in this stage the sheep have the lowest flexibility and capacity to accommodate the food constrain; this situation is more important when the nutrition environment is limited, where the mobilisation of body reserves is a key factor (Ingvartsen et al. Citation1999). For this reason, although some early studies suggest that BW is not a determinant of milk production with a null relationship (Mavrogenis and Papachristoforou Citation2000), the importance of BW during pregnancy and lactation is in relation at the energy partition towards milk yield with a drastic body tissue mobilisation and less energy focussed to growth, fertility and immunity (Veerkamp and Koenen Citation1999) with consequences in animal health, well-being and welfare.
In the present study, the ewes with moderate BW at lambing and early lactation showed milk yields similar to high BW ewes can be associated with the potential of dairy animals to compensate for the low nutrient intake during the pre-parity period by a high intake in early lactation (Agenas et al. Citation2003) if they are fed with adequate energy-dense feed like it occurred in the current study. The ewes were supplemented at lambing and lactation with a good quality forage and concentrate. The ability to compensate is not enough for low BW ewes to reach milk yields of moderate and high BW, which is likely due to the low level of body reserves available that must be mobilised at early lactation. Dickerson (Citation1970) points out that the selection within a breed should not be focussed on higher weight per se, but rather aimed at higher yield relative to body size.
Conclusions
The availability of nutrients at the end of pregnancy and during lactation that are provided by the feed ration and body reserve mobilisation are a key point to reach adequate milk yields in dairy sheep. The BW showed an important relationship with the milk production, but the strength of association is according to the productive stage in which the BW was measured. The BW at pregnancy, lambing and lactation must be considered as indicators of animal nutritional status and level body reserves, mainly in systems that base their nutritional management on grazing.
Acknowledgements
Mr. Angeles Hernandez thank the National Council for Science and Technology (CONACyT, Mexico) for the scholarship for their studies in the Universidad Nacional Autonoma de Mexico, Dr. Gonzalez Ronquillo was granted with a Beca Alianza del Pacifico 2014 fellowship.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Abdelsayed M, Thomson PC, Raadsma HW. 2015. A review of the genetic and non-genetic factors affecting extended lactation in pasture-based dairy systems. Anim Prod Sci. 55:949–966.
- Agenas S, Burstedt E, Holtenius K. 2003. Effect of feeding intensity during the dry period. 1. Feed intake, body weight and milk production. J Dairy Sci. 86:870–882.
- Alqaisi O. 2007. Reproductive performance and lactation yield of local Awassi, Improved Awassi and Afec-Awassi ewes under contemporary comparison on Al-Khanasry station in Jordan [Thesis]. University of Hohenheim.
- Angeles-Hernandez JC, Albarran-Portillo B, Gomez-Gonzalez AV, Pescador-Salas N, Gonzalez-Ronquillo M. 2013. Comparison of mathematical models applied to f1 dairy sheep lactations in organic farm and environmental factors affecting lactation curve parameter. Asian-Australasian J Anim Sci. 26:1119–1126.
- Angeles-Hernandez JC, Ortega OAC, Perez AHR, Gonzalez-Ronquillo M. 2014. Effects of crossbreeding on milk production and composition in dairy sheep under organic management. Anim Prod Sci. 54:1641–1645.
- Barillet F, Marie C, Jacquin M, Lagriffoul G, Astruc JM. 2001. The French Lacaune dairy sheep breed: use in France and abroad in the last 40 years. Livest Prod Sci. 71:17–29.
- Bauman DE, Currie WB. 1980. Partitioning of nutrients during pregnancy and lactation: a review of mechanisms involving homeostasis and homeorhesis. J Dairy Sci. 63:1514–1529.
- Berry DP, Buckley F, Dillon P. 2007. Body condition score and live-weight effects on milk production in Irish Holstein-Friesian dairy cows. Animal. 1:1351–1359.
- Butorovic N. 2015. Meteorological summary year 2014 “Jorge C. Schythe” Station. An del Inst la Patagon; p. 43.
- Cannas A. 2004a. Feeding of lactating ewes. In: Pulina G, Bencini R, editors. Dairy sheep nutrition. Oxford (UK): CABI Publishing; p. 79–108.
- Cannas A, 2004b. Energy and protein requirements. In: P, G, B. R editors. Dairy sheep nutrition. Oxford (UK): CABI Publishing; p. 31–49.
- Carta A, Macciotta N, Cappio-Borlino A, Sanna SR. 2001. Modelling phenotypic (co)variances of test day records in dairy ewe. Livest Prod Sci. 69:9–16.
- Charismiadou MA, Bizelis JA, Rogdakis E. 2000. Metabolic changes during the perinatal period in dairy sheep in relation to level of nutrition and breed. I. Late pregnancy. J Anim Physiol Anim Nutr. 84:61–72.
- Dag B, Keskin I, Mikailsoy F. 2005. Application of different models to the lactation curves of unimproved Awassi ewes in Turkey. S Afr J Anim Sci. 35:238–243.
- Dickerson G. 1970. Efficiency of animal production-molding the biological components. J Anim Sci. 30:848–859.
- Dimauro C, Atzori AS, Pulina G. 2011. Assessing and optimazing the performance of a mechanistic mathematical model of the sheep mammary gland. In: Sauvant D, M, JV, F, P, F. N, editors. Modelling nutrient digestion and utilisation in farm animals. Wageningen (The Netherlands): Wageningen Academic Publishers; p. 72–82.
- Fahmy MH, Shrestha JNB. 2011. Sheep breeds. In: Fuquay JW, Fox PF, McSweeney PLH, editors. Encyclopedia of dairy sciences. London (UK): Elsevier; p. 325–339.
- Godfrey RW, Gray ML, Collins JR. 1997. Lamb growth and milk production of hair and wool sheep in a semi-arid tropical environment. Small Rumin Res. 24:77–83.
- Gootwine E. 2011. Mini review: breeding Awassi and Assaf sheep for diverse management conditions. Trop Anim Health Prod. 43:1289–1296.
- Gootwine E, Goot H. 1996. Lamb and milk production of Awassi and East Friesian sheep and their crosses under a mediterranean environment. Small Rumin Res. 20:255–260.
- Gootwine E, Pollott GE. 2000. Factors affecting milk production in improved Awassi dairy ewes. Anim Sci. 71:807–815.
- Haenlein GFW, Wendorff WL. 2006. Sheep milk. In: Park YW, Haenlein GFW, editors. Handbook of Milk of Non-Bovine Mammals. Victoria: Blackwell Publishing; p. 137–194.
- Hansen LB, Cole JB, Marx GD, Seykora AJ. 1999. Productive life and reasons for disposal of Holstein cows selected for large versus small body size. J Dairy Sci. 82:795–801.
- Harville DA, Henderson CR. 1966. Interrelationships among age, body weight, and production traits during first lactations of dairy cattle. J Dairy Sci. 49:1254–1261.
- Hunter TE, Suster D, DiGiacomo K, Cummins LJ, Dunshea FR, Egan AR, Leury BJ. 2015. Milk production and body composition of single-bearing East Friesian × Romney and Border Leicester × Merino ewes. Small Rumin Res. 131:123–129.
- ICAR. 2016. Recording guidelines. Int Comm Anim Rec. 59–75.
- Ingvartsen KL, Friggens NC, Faverdin P. 1999. Food intake regulation in late pregnancy and early lactation. Metab Stress Dairy Cows. 24:37–54.
- Laporte-Broux B, Duvaux-Ponter C, Roussel S, Promp J, Chavatte-Palmer P, Ponter AA. 2011. Restricted feeding of goats during the last third of gestation modifies both metabolic parameters and behaviour. Livest Sci. 138:74–88.
- Louca A, Mavrogenis A, Lawlor MJ. 1974. Effects of plane of nutrition in late pregnancy on lamb birth weight and milk yield in early lactation of Chios and Awassi sheep. Anim Prod. 19:341–349.
- Mavrogenis AP, Hancock J, Louca A. 1980. The effect of body-weight changes during pregnancy and lactation on the performance of three breeds of sheep. J Agric Sci. 95:357–363.
- Mavrogenis AP, Papachristoforou C. 2000. Genetic and phenotypic relationships between milk production and body weight in Chios sheep and Damascus goats. Livest Prod Sci. 67:81–87.
- Mayer E, Liebich HG, Arbitman R, Hagemeister H, Dirksen G. 1986. Nutritionally-induced changes in the rumenal papillae and in their capacity to absorb short chain fatty acids in high producing dairy cows. Paper presented at Proceedings of the 14th World Congress on Diseases of Cattle; p. 806–817.
- Mckusick BC, Thomas DL, Berger YM. 2001. Effect of weaning system on commercial milk production and lamb growth of East Friesian dairy sheep. J Dairy Sci. 84:1660–1668.
- Mellor DJ, Murray L. 1985. Effects of maternal nutrition on udder development during late pregnancy and on colostrum production in Scottish Blackface ewes with twin lambs. Res Vet Sci. 39:230–234.
- Neville TL, Meyer AM, Reyaz A, Borowicz PB, Redmer DA, Reynolds LP, Caton JS, Vonnahme KA. 2013. Mammary gland growth and vascularity at parturition and during lactation in primiparous ewes fed differing levels of selenium and nutritional plane during gestation. J Anim Sci Biotechnol. 4:6.
- Paten AM, Duncan EJ, Pain SJ, Peterson SW, Kenyon PR, Blair HT, Dearden PK. 2015. Functional development of the adult ovine mammary gland-insights from gene expression profiling. BMC Genomics. 16:748.
- Paten AM, Pain SJ, Peterson SW, Blair HT, Kenyon PR, Dearden PK, Duncan EJ. 2014. Identification of reference genes for RT-qPCR in ovine mammary tissue during late pregnancy and lactation and in response to maternal nutritional programming. Physiol. Genomics. 46:560–570.
- Pollott GE. 2000. A biological approach to lactation curve analysis for milk yield. J Dairy Sci. 83:2448–2458.
- Ruiz R, Oregui LM, Herrero M. 2000. Comparison of models for describing the lactation curve of latxa sheep and an analysis of factors affecting milk yield. J Dairy Sci. 83:2709–2719.
- Sahlu T, Hart SP, Le-Trong T, Jia Z, Dawson L, Gipson T, Teh TH. 1995. Influence of prepartum protein and energy concentrations for dairy goats during pregnancy and early lactation. J Dairy Sci. 78:378–387.
- SAS Institute. 2002. Statistical analysis systems user’s guide. SAS Cary, NC.
- Sieber M, Freeman AE, Kelley DH. 1988. Relationships between body measurements, body weight, and productivity in Holstein dairy cows. J Dairy Sci. 71:3437–3445.
- Swanson TJ, Hammer CJ, Luther JS, Carlson DB, Taylor JB, Redmer DA, Neville TL, Reed JJ, Reynolds LP, Caton JS, et al. 2008. Effects of gestational plane of nutrition and selenium supplementation on mammary development and colostrum quality in pregnant ewe lambs. J Anim Sci. 86:2415–2423.
- van der Linden DS, Kenyon PR, Blair HT, Lopez-Villalobos N, Jenkinson CMC, Peterson SW, Mackenzie DDS. 2009. Effects of ewe size and nutrition on fetal mammary gland development and lactational performance of offspring at their first lactation. J Anim Sci. 87:3944–3954.
- Veerkamp RF. 1998. Selection for economic efficiency of dairy cattle using information on live weight and feed intake: a review. J Dairy Sci. 81:1109–1119.
- Veerkamp RF, Koenen EPC. 1999. Genetics of food intake, live weight, condition score and energy balance. Metab Stress Dairy Cows. 24:63–73.
- Veerkamp RF, Thompson R. 1999. A covariance function for feed intake, live weight, and milk yield estimated using a random regression model. J Dairy Sci. 82:1565–1573.