
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Population-level phenotyping is, for some traits, hardly achievable due to limitation of reference analyses. Mid-infrared spectroscopy (MIRS) is a quick and cost-effective laboratory technique allowing the prediction of innovative milk quality features on a large scale. The present study aimed at investigating factors associated with milk Ca, K, Mg, Na and P contents predicted by MIRS models on a large multi-breed spectral data set of 123,240 test-day records. Two dairy (Holstein–Friesian and Brown Swiss) and two dual-purpose (Simmental and Alpine Grey) cattle breeds were considered. Sources of variation of predicted milk mineral contents were studied using linear mixed models, including the fixed effects of breed, month of sampling, stage of lactation, parity and interactions between the main effects. Random factors were herd nested within breed, cow nested within breed and the residual. Calcium content was greater in milk of dual-purpose than dairy cattle breeds. Simmental cows produced milk with the greatest content of Ca and Na, and milk of Holstein-Friesian had the lowest P content. Variation of content of Ca, Mg, Na and P throughout lactations exhibited an opposite trend to that of milk yield, with the lowest values around the peak of lactation. On the other hand, K content mirrored the trend of milk yield. Multiparous cows had lower content of milk minerals, with the exception of Na, compared with first parity animals. Environmental factors identified in the present study can be considered for within-breed genetic evaluation to adjust records of milk mineral contents for these effects.
Introduction
The acquisition of individual-level phenotypic information using rapid and cost-effective methodologies is becoming an extremely relevant challenge, since genomic information is nowadays easily available at low-cost. However, large-scale collection of economically important traits is often hampered by high recording costs, thus preventing their inclusion in breeding programmes and in quality-based payment systems. Mid-infrared spectroscopy (MIRS) is worldwide recognised as one of the most accessible and efficient technologies to collect phenotypes at individual level (De Marchi et al. Citation2014). In national milk recording systems, individual cow milk samples are routinely collected and analysed through MIRS to determine their chemical composition (fat, protein, casein and lactose percentages). The output of milk MIRS analysis (i.e. spectrum) is a representation of the energy absorbed by the sample itself, when hit by the light, at a specific wavenumber in the region between 900 and 5000 cm−1. Storage of this information is feasible and once new MIRS prediction models are developed, spectra can be used to predict new phenotypes at no additional costs. Mid-infrared spectroscopy prediction models for milk mineral content (Ca, K, Mg, Na and P) have been developed by Visentin et al. (Citation2016) using an algorithm of variable selection (Gottardo et al. Citation2015) to improve the accuracy of prediction. The use of variable selection algorithms, coupled with partial least squares (PLS) regression, is fundamental for data reduction and for an improvement of data interpretability (Mehmood et al. Citation2012).
An adequate daily intake of minerals is important to maintain both infant and adult people at a good health (Cashman Citation2006; Haug et al. Citation2007). Moreover, milk minerals, particularly Ca and P, are important indicators of milk quality at the dairy industry level because they play a central role in the coagulation process of milk and thus on cheese-making efficiency (Lucey and Fox Citation1993). There is a paucity of studies that have investigated phenotypic variation of milk mineral content at population level, mainly because its determination through reference analysis is costly and time-demanding. Therefore, the objective of the present research was to investigate factors associated with milk minerals predicted by MIRS in a large database of dairy and dual-purpose cattle breeds.
Materials and methods
Data
The initial data set comprised 132,380 spectra of individual milk samples from 15,173 cows collected during the monthly test-day recording in Bolzano province (Italy) between January 2012 and December 2013 (Visentin et al. Citation2015). The study area is mostly characterised by small farms with traditional feeding (forage or hay and concentrates), and usually during the summer season, cows are moved to highland pastures. The data set included records of four cattle breeds reared in single-breed herds: Holstein–Friesian (HF, n = 2020 cows), Brown Swiss (BS, n = 5475 cows), Alpine Grey (AG, n = 3154 cows) and Simmental (SI, n = 4524 cows). Animals were from 6 to 450 days in milk and from parity 1 to 15. Preservative was added to milk samples (Bronysolv; ANA.LI.TIK Austria, Vienna, Austria) immediately after collection and processed according to International Committee for Animal Recording (ICAR) recommendations at the milk laboratory of the South Tyrol Dairy Association (Sennereiverband Südtirol, Bolzano, Italy). For each sample, protein (%), casein (%) and fat (%) were determined, and spectral information, containing 1060 infra-red transmittance data in the region between 900 and 5000 cm−1, were stored using a MilkoScan FT6000 (Foss Electric A/S, Hillerød, Denmark). Values of somatic cell count (SCC) were assessed by Fossomatic (Foss Electric A/S, Hillerød, Denmark) and transformed to somatic cell score (SCS) through the formula SCS = 3 + log2(SCC/100,000).
Reference analyses and MIRS prediction models
Detailed information about samples collection, reference analyses and development of MIRS prediction models is available from Visentin et al. (Citation2016). Briefly, in 2014, a total of 251 individual bovine milk samples were collected in the same province and herds contributing to the data of the present study. Reference analysis for the determination of Ca, K, Mg, Na and P was conducted in the laboratory of the Department of Agronomy, Food, Natural Resources, Animals and Environment of the University of Padova (Legnaro, Italy) using Inductively Coupled Plasma Optical Emission Spectrometry, Ciros Vision EOP (SPECTRO Analytical Instruments GmbH, Kleve, Germany) after mineralisation.
Prediction models were developed using PLS regression after uninformative variable elimination procedure as described by Visentin et al. (Citation2016). Coefficients of determination (root mean square errors) in validation were 0.67 (122.00 mg/kg), 0.69 (120.00 mg/kg), 0.65 (12.50 mg/kg), 0.40 (70.00 mg/kg) and 0.68 (88.12 mg/kg) for Ca, K, Mg, Na and P, respectively.
Phenotypic characterisation
Statistical analyses were performed using SAS software ver. 9.3 (SAS Institute Inc., Cary, NC). Spectral information expressed in transmittance was transformed into absorbance by taking the log10 of the reciprocal of the transmittance. Prior to multivariate analysis, two high noise level regions (1600–1700 cm−1 and 3040–3660 cm−1), which are known to be related to water absorption (Hewavitharana and van Brakel Citation1997) were discarded. Principal component analysis was carried out in order to identify the spectra (n = 132,380) similar to those used to develop the MIRS prediction models (n = 251). The first six principal components were retained as explaining the 97.02% of the total variance. The Mahalanobis distance from the centroid of the cluster of the spectra used to develop the MIRS models was calculated to identify the outlier spectra. Instrumental or temporal drifts did not represent an issue given that the laboratory equipment was the same for the whole duration of the study. Moreover, the principal component analysis did not identify two evident clusters based on the different years of data collection (2012–2013). Following this approach, 9140 spectra were discarded from further analysis. Mid-infrared spectroscopy prediction models were then applied on the remaining 123,240 spectra from 14,389 cows to predict major milk minerals. For each milk composition trait (i.e. protein, casein and fat percentages, and SCS), as well as for each predicted milk mineral, values that deviated more than 3 standard deviations from the respective mean were discarded.
Sources of variation of milk mineral content were investigated using the following mixed linear model:
where yijklmno is MIRS-predicted content of Ca, K, Mg, Na or P, μ is the overall intercept of the model, Bi is the fixed effect of the ith breed (i= HF, BS, AG, SI), Mj is the fixed effect of the jth month of sampling (j = 1 to 12), Yk is the fixed effect of the kth year of sampling (k = 2012, 2013), Sl is the fixed effect of the lth class of stage of lactation of the cow (l = 1 to 45, with the first being a class from 6 to 10 d, followed by 10-d classes), Paritym is the fixed effect of the mth parity of the cow (m = 1 to 5, with class 5 including cows from parity 5 to 15), (B×M)ij is the fixed interaction effect between breed and month of sampling, (B×S)il is the fixed interaction effect between breed and stage of lactation, (B×Parity)im is the fixed interaction effect between breed and parity, (S×Parity)lm is the fixed interaction effect between stage of lactation and parity, Hn(Bi) is the random effect of the nth herd nested within the ith breed ∼N(0, σ2H(B)), Cowo is the random effect of the oth cow nested within the ith breed ∼N(0, σ2COW(B)), and eijklmno is the random residual ∼N(0, σ2e). Significance of breed effect was tested on the cow within breed variance. A multiple comparison of means was performed for breed effect using Bonferroni’s test (p < .05).
The same approach of data selection, application of MIRS prediction models and generation of MIRS-predicted phenotypes has been applied by Visentin et al. (Citation2015) to predict milk coagulation properties (MCP) of the same milk samples considered in the present study. Milk coagulation properties investigated by Visentin et al. (Citation2015) were rennet coagulation time (RCT, min; the time taken from rennet addition to the development of the gel), curd-firming time (k20, min; the time taken from gel development to the achievement of a 20 mm strong curd) and curd firmness at 30 min (a30, mm; the strength of the curd after 30 min from rennet addition). Pearson correlations between milk minerals were assessed, and also between such traits and MCP from Visentin et al. (Citation2015).
Results
Descriptive statistics and phenotypic correlations
Table summarises the descriptive statistics for milk yield, chemical composition, SCS and minerals of samples retained after removal of spectral outliers. Milk yield averaged 22.74 kg/d, and means for fat, protein and casein were 4.03%, 3.53% and 2.76%, respectively. Minerals predicted by MIRS were characterised by moderate to large coefficients of variation, which ranged from 13.13% (P) to 23.36% (Mg). The proportion of phenotypic variance of milk minerals accounted by the cow effect ranged from 2.01% (K) to 32.45% (P), and the ratio between herd and phenotypic variance was between 4.80% (Mg) and 12.92% (Ca).
Table 1. Descriptive statistics and variance accounted by cow (σ2C) and herd (σ2H) effects for milk yield, composition, somatic cell score (SCS), mineral content and coagulation properties.
Pearson correlations (r) between milk minerals (Table ) varied from −0.20 (Ca and K, and K and Na) to 0.66 (K and Mg). Moderate to low relationships were assessed between Ca and P (r = 0.53), Na and P (r = 0.32), Ca and Na (r = 0.29) and Mg and P (r = 0.24) (p < .05). Calcium and P were the minerals with the strongest association with milk protein and casein percentages, ranging from 0.31 (Ca with protein) to 0.37 (P with casein) (p < .05). Strong and unfavourable correlations were estimated between RCT and a30 (r = −0.87), and between k20 and a30 (r = −0.64) (p < .05). Curd-firming time was unfavourably associated with protein (r = −0.58) and casein percentages (r = −0.61) (p < .05), whereas a30 was favourably correlated with these constituents (r = 0.21 with protein and r = 0.23 with casein percentages; p < .05). Rennet coagulation time was weakly correlated with milk composition. Regarding correlations between MCP and minerals, Ca and P were the elements with the greatest association with milk coagulation ability, and correlations ranged from −0.35 and −0.39 with k20 to 0.37 and 0.28 with a30, respectively (p < .05). Finally, SCS was positively and moderately associated with Na (0.22; p < .05).
Table 2. Pearson correlations between milk chemical composition, somatic cell score (SCS), mineral content and coagulation traits predicted by mid-infrared spectroscopy.
Breed effect
Least squares means of milk yield, chemical composition and mineral content for HF, BS, AG and SI breeds are in Table . Milk production ranged from 14.67 kg/d (AG) to 25.28 kg/d (HF). Fat, protein and casein percentages of milk from BS cows were greater (p < .05) than those of the other breeds, and AG exhibited the poorest fat percentage (3.93%; p < .05). Regarding the nitrogen fraction, AG and SI breeds had intermediate levels of both protein (3.57% and 3.58%, respectively) and casein (2.79% and 2.80%, respectively), and HF cows had the poorest percentage of both these constituents (p < .05). The greatest (2.77) and lowest (2.50) values of SCS were estimated for BS and SI, respectively.
Table 3. Least squares means (with standard errors) of milk yield, composition, somatic cell score (SCS) and mineral content of cows of different breeds.
Overall, mineral content varied significantly across breeds. In particular, milk of SI cows had the greatest content of Ca and Na (1426.44 mg/kg and 460.69 mg/kg, respectively; p < .05) and that of HF exhibited the lowest content of Ca and P (1306.60 mg/kg and 972.68 mg/kg, respectively; p < .05). Milk of AG breed had significantly (p < .05) greater Ca (1370.06 mg/kg) than that of HF (1306.60 mg/kg) and BS (1350.42 mg/kg).
Lactation, parity, and temporal effects
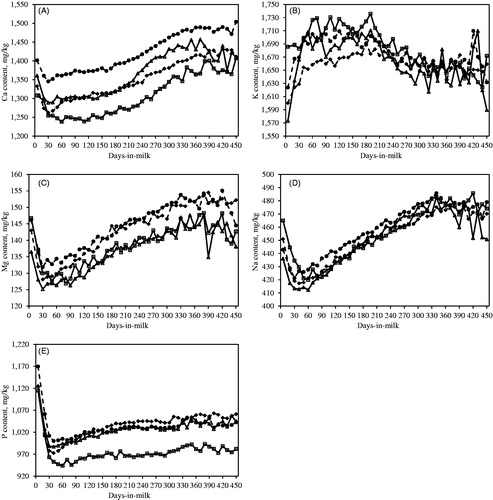
Figure depicts the least squares means of predicted milk mineral composition across lactation for HF, BS, AG and SI cows. Calcium, Mg, Na and P contents exhibited an opposite trend to that of milk yield across days in milk, having the minimum peak of content at the maximum peak for milk yield, whereas K was the only mineral which resembled the trend of milk yield across lactation.
Figure 1. Least squares means of (A) Ca, (B) K, (C) Mg, (D) Na and (E) P content across lactation for Holstein-Friesian (—□—), Brown Swiss (- -♦- -), Alpine Grey (—△—) and Simmental (- -•- -) cows.
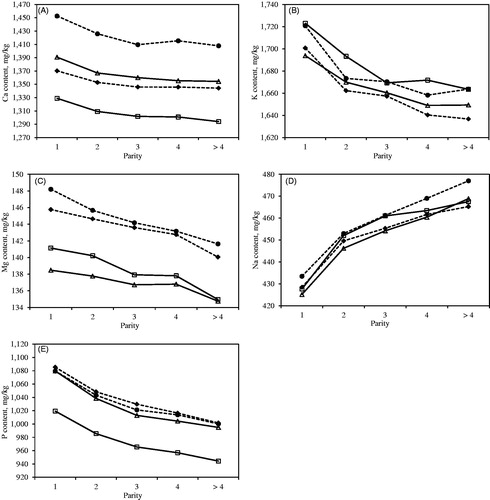
Trend of predicted mineral composition across parity is depicted in Figure . With the exception of Na, minerals contents were greater in milk of primiparous than multiparous cows, and the lowest values were observed for cows at fifth and later parities. Milk minerals content also varied across the calendar month of the year with similar trends for all the four cattle breeds considered in the present study (data not shown). The decrease in all minerals content, which occurred in the first part of the year, halted during the summer season (June to September).
Figure 2. Least squares means of (A) Ca, (B) K, (C) Mg, (D) Na and (E) P content across parity for Holstein-Friesian (—□—), Brown Swiss (- -♦- -), Alpine Grey (—△—) and Simmental (- -•- -) cows.
Discussion
In order to generate robust prediction models, large variability of both spectral and trait(s) of interest is desired in the calibration data set. Mean mineral contents were consistent with findings of Soyeurt et al. (Citation2009) on a data set (n = 100) of HF, Jersey, Belgian Blue, Montbéliarde and Normande breeds and Toffanin et al. (Citation2015) on 208 HF cows. Moreover, coefficients of variation of milk minerals calculated in the present study were in accordance with the results of Soyeurt et al. (Citation2009). The variability of the spectral data used to develop the MIRS prediction models was sufficient to represent the 93% of the total spectra variation. This was somewhat expected given that the dataset used to develop the prediction models by Visentin et al. (Citation2016) included samples of the same province and breeds considered in the present study.
The importance of milk minerals on MCP has been reported by Malacarne et al. (Citation2014) and Toffanin et al. (Citation2015), who demonstrated that milk samples with optimal coagulation profile were those with the greatest content of Ca (and P), particularly colloidal and total Mg, and the lowest values of chloride. The correlations between milk minerals, and between milk minerals and milk composition were expected since about two-third of Ca and P, and one-third of Mg measured in milk, are bonded to form calcium phosphate, which plays an essential role for the casein micelle structure (Holt et al. Citation2013). Finally, the positive association between Na and SCS was probably due to physiological reason; indeed, an infection to the mammary gland, which results in higher SCS, increases the permeability of blood capillaries. Therefore, Na and chloride, which are more concentrated into the extracellular liquid, are poured into the mammary lumen (Summer et al. Citation2009).
Effects of cow breed on predicted traits
The differences of milk chemical composition across cow breeds have been well documented in literature (Barlowska et al. Citation2006; Carroll et al. Citation2006; Malacarne et al. Citation2006; De Marchi et al. Citation2007; Penasa et al. Citation2014) and they were consistent with the results of the present study. Milk mineral characterisation of HF was investigated by Carroll et al. (Citation2006) and van Hulzen et al. (Citation2009). Also, milk mineral profile of SI and BS breeds was studied by Barlowska et al. (Citation2006) and Carroll et al. (Citation2006), respectively. To our knowledge, the present study is the first dealing with the determination of milk minerals content of AG cows, a local cattle breed reared in the Alps. Alpine local breeds are part of cultural and social traditions and, although characterised by lower productivity compared to HF cows, they are more adapted to marginal areas. Moreover, De Marchi et al. (Citation2007), in agreement with results of the present study, demonstrated that local breeds, including AG, produced milk with more favourable processing characteristics (i.e. shorter RCT and greater a30) than HF and BS cows. Recently Pretto et al. (Citation2013) estimated a positive association between a30 and cheese yield, and such a conclusion could have serious implications for the valorisation of local Alpine breeds.
Effects of stage of lactation and parity on predicted traits
Variation of Ca, Mg, Na and P contents throughout lactation (Figure ) was consistent with Carroll et al. (Citation2006) and van Hulzen et al. (Citation2009), following an opposite trend than milk yield. Therefore, a dilution effect could be assumed to explain this variation. Moreover, the increase of the aforementioned elements could be associated with the higher casein level of milk from cows in late lactation, as reported by Holt et al. (Citation2013). Particularly, at the onset of lactation, an excess of dietary cations is a risk factor for milk fever, and forages normally offered to dry cows have high K content (Goff and Horst Citation1997). Therefore, the increment of milk K content observed in the present study in early lactation might be due to a dietary excess of this mineral, although no data on feeding rations were available to justify this hypothesis. Van Hulzen et al. (Citation2009) estimated that herd variance for K was, on average, 50% greater than herd variance for other minerals, such as Ca, Mg, P and Zn, and suggested that management strategies (such as feeding) could have a relatively high impact on K variation. Such a statement on K content was also supported by the results of the present study, as the proportion of phenotypic variance accounted by the herd effect was larger than the variance accounted by the cow effect. The variation across calendar month for milk minerals might be due to the change in the diet of animals reared in the Alpine area, since a quite common practice in Alpine regions is to move cows to highland pastures during summer (June to September).
Least squares means of Ca and P contents across parity (Figure ) were consistent with the findings of Kume et al. (Citation1998), who reported the same decreasing trend in Holstein dairy cows. Indeed, it has been hypothesised that the utilisation of these minerals at the mammary gland level is lower in multiparous cows (Kume and Tanabe Citation1993). In particular, the mobilisation of Ca from bones to blood is less efficient due to a lower activity of parathyroid hormone, which is involved in osteoplastic activities. In the present study, a reduction of casein percentage in older cows was observed, and this evidence could explain the decreasing trend of Ca, P and Mg contents, which are known to be mainly bonded to casein micelle. The lowest K and the highest Na contents were observed in multiparous cows, and this was in association with the increasing trend of SCS across parity. Great Na and low K levels are associated with high SCS (Summer et al. Citation2009), which is an indicator of (sub)clinical mastitis. Overall, the decreasing trend for Ca, K, Mg and P might be also explained by a dilution effect of these minerals due to higher milk yield across parity observed in the present study.
Conclusions
Mid-infrared spectroscopy prediction models can be successfully applied to large spectral datasets for phenotypic characterisation of major minerals in milk. Breed, stage of lactation and parity were important sources of variation of major milk mineral contents at the population level, and significant differences among breeds were detected for all traits. Dual-purpose breeds were characterised by greater mineral contents compared to dairy breeds. Factors identified in the present study will be useful for within-breed genetic analyses to adjust records for significant environmental effects.
Acknowledgments
The authors would like to thank Luis Kerschbaumer and Thomas Kerschbamer (South Tyrol Dairy Association – Sennereiverband Südtirol, Bolzano, Italy), Christian Plitzner (Breeders Association of Bolzano province – Vereinigung der Südtiroler Tierzuchtverbände, Bolzano, Italy) and Claudio Stecchi (CTS informatica srl, Bolzano, Italy) for providing data used in this study and for technical support.
Disclosure statement
The authors declare that there is no conflict of interest associated with the paper. The authors alone are responsible for the content and writing of this article.
Funding
This study was supported by the University of Padova (Ricerca Scientifica fondi quota EX 60% - 2014, project 60A08-7955/14, Italy).
References
- Barlowska J, Litwińczuk Z, Król J, Kędzierska-Matysek M. 2006. Fatty acid profile and minerals content in milk from cows of various breeds over spring-summer feeding period. Pol J Food Nutr Sci. 15:13–16.
- Carroll SM, DePeters EJ, Taylor SJ, Rosenberg M, Perez-Monti H, Capps VA. 2006. Milk composition of Holstein, Jersey, and Brown Swiss cows in response to increasing levels of dietary fat. Anim Feed Sci Technol. 131:451–473.
- Cashman KD. 2006. Milk minerals (including trace elements) and bone health. Int Dairy J. 16:1389–1398.
- De Marchi M, Dal Zotto R, Cassandro M, Bittante G. 2007. Milk coagulation ability of five dairy cattle breeds. J Dairy Sci. 90:3986–3992.
- De Marchi M, Toffanin V, Cassandro M, Penasa M. 2014. Invited review: mid-infrared spectroscopy as phenotyping tool for milk traits. J Dairy Sci. 97:1171–1186.
- Goff JP, Horst RL. 1997. Effects of the addition of potassium or sodium, but not calcium, to prepartum rations on milk fever in dairy cows. J Dairy Sci. 80:176–186.
- Gottardo P, De Marchi M, Cassandro M, Penasa M. 2015. Technical note: Improving the accuracy of mid-infrared prediction models by selecting the most informative wavelengths. J Dairy Sci. 98:4168–4173.
- Haug A, Høstmark AT, Harstad OM. 2007. Bovine milk in human nutrition-a review. Lipids Health Dis. 6:25.
- Hewavitharana AK, van Brakel B. 1997. Fourier transform infrared spectrometric method for the rapid determination of casein in raw milk. Analyst. 122:701–704.
- Holt C, Carver JA, Ecroyd H, Thorn DC. 2013. Invited review: caseins and the casein micelle: their biological functions, structures, and behavior in foods. J Dairy Sci. 96:6127–6146.
- Kume S, Tanabe S. 1993. Effect of parity on colostral mineral concentrations of Holstein cows and value of colostrum as a mineral source for newborn calves. J Dairy Sci. 76:1654–1660.
- Kume S, Yanamoto E, Kudo T, Toharmat T, Nonaka I. 1998. Effect of parity on mineral concentration in milk and plasma of Holstein cows during early lactation. Asian-Australas J Anim Sci. 11:133–138.
- Lucey JA, Fox PF. 1993. Importance of calcium and phosphate in cheese manufacture: a review. J Dairy Sci. 76:1714–1724.
- Malacarne M, Summer A, Fossa E, Formaggioni P, Franceschi P, Pecorari M, Mariani P. 2006. Composition, coagulation properties and Parmigiano-Reggiano cheese yield of Italian Brown and Italian Friesian herd milks. J Dairy Res. 73:171–177.
- Malacarne M, Franceschi P, Formaggioni P, Sandri S, Mariani P, Summer A. 2014. Influence of micellar calcium and phosphorus on rennet coagulation properties of cows milk. J Dairy Res. 81:129–136.
- Mehmood T, Liland KH, Snipen L, Saebø S. 2012. A review of variable selection methods in partial least squares regression. Chemometr Intell Lab. 118:62–69.
- Penasa M, Tiezzi F, Sturaro A, Cassandro M, De Marchi M. 2014. A comparison of the predicted coagulation characteristics and composition of milk from multi-breed herds of Holstein-Friesian, Brown Swiss and Simmental cows. Int Dairy J. 35:6–10.
- Pretto D, De Marchi M, Penasa M, Cassandro M. 2013. Effect of milk composition and coagulation traits on Grana Padano cheese yield under field conditions. J Dairy Res. 80:1–5.
- Soyeurt H, Bruwier D, Romnee J-M, Gengler N, Bertozzi C, Veselko D, Dardenne P. 2009. Potential estimation of major mineral contents in cow milk using mid-infrared spectrometry. J Dairy Sci. 92:2444–2454.
- Summer A, Franceschi P, Malacarne M, Formaggioni P, Tosi F, Tedeschi G, Mariani P. 2009. Influence of somatic cell count on mineral content and salt equilibria of milk. Ital J Anim Sci. 8:435–437.
- Toffanin V, De Marchi M, Lopez-Villalobos N, Cassandro M. 2015. Effectiveness of mid-infrared spectroscopy for prediction of the contents of calcium and phosphorus, and titratable acidity of milk and their relationship with milk quality and coagulation properties. Int Dairy J. 41:68–73.
- van Hulzen KJE, Sprong RC, van der Meer R, van Arendonk JAM. 2009. Genetic and nongenetic variation in concentration of selenium, calcium, potassium, zinc, magnesium, and phosphorus in milk of Dutch Holstein-Friesian cows. J Dairy Sci. 92:5754–5759.
- Visentin G, Penasa M, Gottardo P, Niero G, Isaia M, Cassandro M, De Marchi M. 2015. Milk coagulation properties of cattle breeds reared in the Alpine area. Poljoprivreda. 21:237–240.
- Visentin G, Penasa M, Gottardo P, Cassandro M, De Marchi M. 2016. Predictive ability of mid-infrared spectroscopy for major mineral composition and coagulation traits of bovine milk by using the uninformative variable selection algorithm. J Dairy Sci. 99:8137–8145.