
Abstract
Previous grazing experience on high alpine pastures may help cows to optimise feed selection and minimise impairments in milk production. Eight inexperienced and eight experienced Aosta Red Pied cows were compared (primiparous:multiparous = 1:1). Measurements were performed when cows grazed sites at 600, 1800 and 2100 m a.s.l. The statistical model included experience, parity and site as fixed effects. The longest ingestion time was recorded for primiparous inexperienced cows at 1800 m a.s.l. Milk yield remained unaffected by experience, but the main milking time was shorter in inexperienced cows in the lowlands. Milk of inexperienced cows had a higher urea content than experienced cows. Somatic cell score remained unaffected by experience, but inexperienced cows had more incidences of clinical mastitis on alpine pastures. Compared with experienced cows, proportions of the fatty acids ΣC16:1 and ΣC17:1 in milk fat of inexperienced cows were higher and proportions of C18:1 t9 and C18:1 t6–8 lower. Overall, alpine grazing had substantial effects on milk yield and composition coinciding with results from previous studies. There were interactions between experience and site in milking characteristics and milk composition, and for the latter also between experience and parity. In conclusion, experience had effects on several of the variables tested, effects which were sometimes exhibited already on lowland pasture. Still, the effects of experience were much lower than those of the transhumance system.
The advantages of previous site-specific and age experience of cows were studied on alpine pasture.
Lack of experience had mostly minor effects on economically relevant traits.
Experience helped to maintain udder health in this alpine environment.
Highlights
Introduction
Transhumance for exploiting high-altitude pastures for milk production during summer is a traditional practice in alpine regions. Effects of alpine conditions on the cow’s physiological status and links to milk and cheese properties were investigated in various previous studies (e.g. Leiber et al. Citation2006; Sturaro et al. Citation2013; Farruggia et al. Citation2014; Zendri et al. Citation2016). Conditions in the hypoxic environment of alpine pastures differ from those in the lowlands and therefore require metabolic and behavioural adaptation of cows. In addition, alpine grazing regularly results in a milk yield decline as a consequence of moving to and staying on these pastures (Kreuzer et al. Citation1998; Leiber et al. Citation2006; Zendri et al. Citation2016).
Experience affects feeding behaviour and physiological status of ruminants. Accordingly, grazing experience acquired at a juvenile age affects diet selection years later in cows (Lopes et al. Citation2013). Early exposure of lambs to dietary diversity affected their acceptance of new flavours in new environments and reduced stress (Villalba et al. Citation2012). Primiparous cows, exposed to a high-quality pasture early in life, exhibited higher milk yields during the first days on pasture than cows unfamiliar with grazing (Lopes et al. Citation2013). Alpine animal caretakers also report about behavioural differences between cows experienced with the specific conditions of a distinct alp and others arriving there for the first time (Silbernagel Citation2002). This included that experienced cows remembered the places on alpine pastures where the most palatable feed grows from the previous year. However, it remains unclear whether this knowledge is helpful for feed selection and maintaining milk yield over the whole season. A different feeding behaviour might also affect the alpine-specific fatty acid (FA) profile of the milk, as cows might ingest herbage with a different botanical composition (Elgersma Citation2015). Low-input systems practised on biodiverse alpine pastures allow such selection and, concomitantly, affect amount and profile of plant secondary compounds (PSC) ingested (Willems et al. Citation2014). Accordingly, body lipids of lambs and milk fatty acid profiles of goats varied between different alpine pastures (Willems et al. Citation2014; Iussig et al. Citation2015).
The aim of the present study was to test the possible advantages of previous site-specific alpine grazing experience on feeding behaviour, performance, milking characteristics and milk composition. It was further tested if multiparous cows, due to the experience obtained as lactating cows before, had an advantage over primiparous cows. The investigation was performed in an established transhumant mountain system in the Aosta Valley (Northwestern Italy) with autochthonous Aosta Red Pied (ARP) cows.
Materials and methods
Experimental design
All animal-related procedures were in compliance with EU Directive 2010/63/E.U. Supplementary Table S1 gives an overview of the experimental schedule. Sixteen ARP cows were used where half were ‘experienced’ and originated from the herd of the Institut Agricole Régional (IAR, Aosta, Italy). They had experience on the experimental alpine pastures (>800 m a.s.l.) as cows and on comparable plots in the same valley as heifers before. The other half, obtained from a private herd (Pollein, Aosta Valley, Italy), had never experienced alpine pastures on comparable sites, either as heifers or cows. Animals were randomly selected from these two herds in a way that in both groups, (1) half of the cows were primiparous and half multiparous, and (2) mean values for DIM, milk yield, fat and protein contents were similar during the pre-experimental period. Accordingly, pre-experimental milk yields were 16.3 ± 3.5 (mean ± standard deviation) and 15.3 ± 3.2 kg/day in experienced and inexperienced cows, respectively. The corresponding milk composition was 32.2 ± 3.2 and 32.4 ± 2.8 g fat/kg, and 31.5 ± 1.4 and 35.6 ± 1.9 g protein/kg, and cows weighed 514 ± 25 and 522 ± 21 kg. Experienced and inexperienced cows were 121 ± 33 and 114 ± 30 days in milk (DIM), respectively. Individual milk yield, fat and protein contents had been obtained from milk control (one sample/cow/month during 5 months) in their respective home-tied stalls, where they had received hay from their respective lowland pastures and 3 kg/day of concentrate (Mangime Settebello Ma. Co. Pa., Mareine & Cie, Bosconero, Italy) containing, per kg of original substance, 925 g organic matter, 208 g crude protein, 85 g crude fibre and 38 g ether extract. Data on feeding behaviour and milking characteristics were individually collected two times during the weeks before the start of the experiment for their use as covariates.
At the beginning of the grazing season (calendar week 16), the 16 experimental cows were integrated into the IAR herd and grazed on lowland pastures. Cows were transported by truck from 600 m (Montfleury, Aosta) to 1800 m a.s.l. (Alp Chaudanne), and later walked on a steep track to Alp Entrelor (2100 m a.s.l.). On each site, samples were collected 9 and 28 days after moving cows to the respective pasture (equivalent to calendar weeks 17 and 20, 25 and 27, and 29 and 32 on the three pastures, Supplementary Table S1). We focussed on medium-term effects (9 days after arrival) of experience in grazing on alpine pastures and therefore sampling started at a time where there were no longer carry-over effects of the transhumance. In the lowlands, cows started grazing outside during daytime, were tethered at night and milked at 05:00 and 16:00 h (AfiMilk, TDM, Brescia, Italy). They received local hay and 3 kg/day of the same concentration as that used in the pre-experimental period and had free access to water. At high altitude, cows had permanent access to pasture and water resources. Throughout the grazing period, a new portion of the pasture was offered daily to the animals (strip grazing technique). Milking was accomplished outside with a mobile milking parlour (Eliar 4, Eli IAR, equipped with AfiMilk machines) at 04:00 and 16:00 h, where cows received exclusively 2 kg/day of the same concentrate manually.
Sampling protocols
Botanical composition of the pastures was recorded using the point-quadrat method (Daget and Poissonet Citation1971). Samples of fresh grass were collected two times per sampling period from 10 cm × 10 m strips dispersed randomly across the area offered to the cows. Compositional analysis was performed with NIRSystems 5000 (Foss, Hillerød, Denmark; Niero et al. Citation2018). Feeding behaviour of the cows was recorded using chewing sensors and data analysis software (MSR Electronics, Henggart, Switzerland) (Nydegger et al. Citation2011). Each of the 16 cows was equipped with these sensors during 2 × 24 h on each site.
Individual milk-related measurements were conducted during two morning and evening milkings per period. A Lactocorder® (WMB AG, Balgach, Switzerland) provided data on milk yield, milking characteristics and electrical conductivity. Milking time was separated into time to reach the plateau (incline phase, flow <0.500 kg/min), duration of plateau and decline phases, and main milking time (start to a decline to 0.200 kg/min). Two 50-ml milk samples/day were taken with the Lactocorder equipment. The morning samples were stored at 4 °C and the evening samples at −20 °C. For the baseline winter barn period, an average between data on milk yield and composition during the 5 months before the experiment was obtained along with the official milk control analyses of the breeders’ association of Aosta. Milk samples then were obtained with the sampling device MPC (AfiMilk, TDM, Brescia, Italy).
In the morning milk, mid-infrared spectroscopy (MilkoScan FT6000, Foss Electric A/S, Hillerød, Denmark) was used to determine milk gross composition. Somatic cells were counted by a fluorimetric method (Fossomatic 5000, Foss Electric, Hillerød, Denmark) and converted into somatic cell scores (SCS; Ali and Shook Citation1980). All devices were calibrated monthly using reference samples (Regione Autonoma Valle Citation2006). Clinical mastitis was assumed when milk conductivity was >9 µS/cm and milk reacted positively with Leucocytest solution (Sacco System, Cadorago, Italy). The percentage of cows with clinical mastitis was calculated. Milk compositional data of these cows were removed from the dataset. The milk FA profile was determined by gas chromatography (Hewlett-Packard HP, Waldbronn, Germany) following Kälber et al. (Citation2011), but with a CP7421 column (200 m × 0.25 mm × 0.25 μm, Varian Inc., Darmstadt, Germany). Individual FA were grouped into saturated FA (SFA), mono-unsaturated FA (MUFA), poly-unsaturated FA (PUFA), n-3 FA, n-6 FA and conjugated linoleic acids (CLA), and the iso-to-anteiso FA. The C14:1 c9 to C14:0 ratio was calculated.
Statistical analysis
After the normality of the data was checked using Shapiro-Wilk’s test, Repeated Time Mixed Procedure was run with SAS version 9.4 (SAS Institute, Cary, NC, USA). Individual cow data obtained in the pre-experimental lowland barn period were used as a parity-centred covariate, as statistical differences were observed for milk yield and protein content already in the barn. As no significant differences between sampling days occurred within the period (9 and 28), these data were averaged. The model included previous site-specific alpine grazing experience (Yes vs. No), parity (1 vs. >1), site (600, 1800 vs. 2100 m a.s.l.) and all their interactions as fixed effects. Site was used as repeated factor with a compound symmetry covariance structure, with the cow (experimental unit) being the subject nested within group. For milk yield, DIM was used as covariate to differentiate between DIM and site effect. Multiple comparisons among means were adjusted with Tukey’s method. Effects were considered significant at p < .05 and as trend at .05 ≤ p < .10. The tables give least square means and standard errors of the mean.
Results
Composition of the grass
Grass from alpine pastures at 1800 and 2100 m a.s.l. was generally richer in acid detergent fibre and lignin than lowland grass (Table ). All other constituents analysed were quite similar at all three sites, with generally moderate contents of crude protein. Details of the botanical composition of the experimental pastures are listed in Supplementary Table S2.
Table 1. Chemical composition of the pasture grass, g/kg dry matter. (n = 2; means ± standard deviation).
Feeding behaviour
Inexperienced cows tended (p < .10) towards a longer rumination time (342 ± 21.7 min/day) compared to experienced cows (286 ± 18.7 min/day) (Table ). Ingestion time (min/day) increased (p < .05) from lowland (287 ± 28.3) to 1800 m a.s.l. (416 ± 33.7) and declined thereafter to 335 ± 34.6. Ingestion time doubled for inexperienced primiparous cows (p < .05) between lowland and 1800 m a.s.l. (p < .05), but not in experienced primiparous cows (three-way interaction, p < .10). The ingestion-to-rumination ratio changed like ingestion time. The multiple comparisons among means using Tukey’s method showed that the total number of ingestion chews (×103/day) was higher (p < .05) at 1800 m (27.6 ± 2.43) than at 600 m (20.1 ± 2.07) and at 2100 m a.s.l. (21.5 ± 2.50), independent of parity and experience, even though the three-way interaction was significant. The number of rumination boli/day was higher (p < .05) at 2100 m (470 ± 21.0) than at 600 m a.s.l. (403 ± 15.5).
Table 2. Effect of experience, site and parity on feeding behaviour.
Milking characteristics and milk properties
The DIM-adjusted milk yield was not affected by the transfer from lowland to 1800 m a.s.l. but declined (p < .05, Table ) from 12.5 to 11.3 kg/day at 2100 m a.s.l. Milking characteristics were mainly influenced by experience and site; changes within sites occurred for experienced cows only. Main milking time declined between lowland (7.27 ± 0.292 min) and alpine sites (5.65 ± 0.284 and 5.39 ± 0.284 min for 1800 and 2100 m a.s.l., respectively) only for experienced cows. The milking plateau phase of experienced cows did not decline at 1800 m a.s.l. (3.27 ± 0.284 min) whereas it was lower (p < .05) at 2100 m a.s.l. (2.57 ± 0.284 min) than at 600 m a.s.l. (3.78 ± 0.292 min). However, the experienced cows’ decline phase was already reduced at 1800 m a.s.l. (2.06 ± 0.284 min) compared to 600 m a.s.l. (3.19 ± 0.292 min) and did not decline between 1800 and 2100 m a.s.l. (2.55 min ± 0.284 min) (Figure ). Maximum milk flow decreased (p < .05) from 1.70 ± 0.101 kg/min to 1.53 ± 0.103 min when cows went from 1800 m to 2100 m a.s.l. Multiparous cows had a longer (p < .05) decline phase and a reduced (p < .05) maximal milk conductivity compared to primiparous cows.
Figure 1. Effect of experience at the sites on durations of main milking time, plateau and decline phase of milking (interactions of experience × site, p < .05, p < .05 and p < .01, respectively). Error bars reflect standard errors. Within the same variable, columns (Least Square means) marked without a common superscript differ (p < .05).
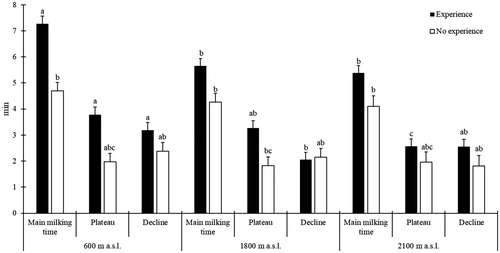
Table 3. Effect of experience, site and parity on milk yield, milking characteristics and milk properties.
There were some effects of experience (without interaction with site or parity) on milk composition, and a number of significant interactions, but not always the same in different traits (Table ). Milk casein content decreased (p < .05) in inexperienced cows at 1800 m a.s.l. but remained unchanged in experienced cows. Milk urea content was 1.4 times higher (p < .01) and, along with a general increase in SCS (p < .05) from lowland to alpine pastures, SCS was 2.4 times higher (p < .05) in inexperienced cows compared to experienced cows, respectively. The latter difference was numerically highest at 600 m a.s.l. and declined thereafter (experience × site, p < .001). More inexperienced than experienced cows exhibited clinical mastitis on alpine pastures (37.5 and 12.5% vs. 12.5 and 0% of inexperienced and experienced cows at 1800 m and 2100 m a.s.l., respectively; data not shown). Milk protein content was higher in primiparous than multiparous experienced cows (36.7 ± 0.76 and 33.5 ± 0.92 g/kg, respectively, p < .05). Milk fat increased (p < .05) from 29.0 ± 1.42 g/kg at 600 m a.s.l. to 34.9 ± 1.56 g/kg at 1800 m a.s.l. and 40.0 ± 1.85 g/kg at 2100 m a.s.l. Milk lactose declined (p < .05) from 600 to 1800 m a.s.l. Site effects on contents of fat were recovered in total solids and on lactose in solids-non-fat.
Table 4. Effect of experience, site and parity on milk composition.
Proportions of some C18:1 trans isomers were influenced by experience, whereas this did not apply for C18:0, C18:1 n-9, C18:2 n-6, C18:2 c9t11 and C18:3 n-3 (Table ). Proportions of C18:1 t6–8 and t9 were higher (p < .05) by 18.2 and 24.3%, respectively, in milk of experienced compared to inexperienced cows. For C18:1 t10 and t12, the experience effect was less systematic. C18:1 t10 proportion was higher (p < .05) in experienced than inexperienced primiparous cows, but only at 600 m a.s.l., and higher (p < .05) than in experienced multiparous cows, but only at 2100 m a.s.l. (p < .05). Apart from an interaction with site for C18:2 c9t11 and C18:1 t11, parity had no effect on C18 FA proportions. Site increased (p < .05) proportions of C18:0, C18:1 t6-8, C18:1 t9, C18:1 t11, C18:2 n-6, C18:2 c9t11 and C18:3 n-3 by 13.9, 14.5, 17.9, 38.1, 19.6, 27.5 and 52.2%, respectively, between 600 and 1800 m a.s.l.
Table 5. Effect of experience, site and parity on selected individual C18 fatty acids in total milk fat (g/100 g).
Among the short- and medium-chain-length FA (C4 to C17) only C8:0, ΣC16:1 and ΣC17:1 were affected (p < .05) by experience and only one interaction (C15:1 c10; with site, p < .01) was found (Supplementary Table S3). Experience enhanced proportions of C8:0 and reduced those of ΣC16:1 and ΣC17:1. Almost every short- and medium-chain-length FA was affected by site. C4:0 proportion increased (p < .05) by 9.4 and 12.6% at 1800 and 2100 m a.s.l. compared to 600 m a.s.l. Proportions of C8:0, C10:0, C12:0, C15:0, C16:0, ΣC16:1 and ΣC17:1 decreased (p < .05) by alpine grazing by 8.8, 15.5, 17.2, 6.3, 9.0, 17.3 and 17.4%, respectively. Proportions of C14:0 and C14:1 c9 decreased by 15.6 and 10.3%, respectively, at 1800 m a.s.l., then increased at 2100 m a.s.l. (p < .05). Experience and parity did not affect proportions of FA > C18 (Supplementary Table S4). Proportions of C20:0, ΣC20:1, C20:2 n-6, C20:4 n-6, C20:4 n-3, C22:0 and C22:1 c13 increased (p < .05) by 12.9, 20.7, 13.7, 76.2, 65.9, 18.8 and 38.9% between 600 and 1800 m a.s.l., respectively. C20:3 n-3 proportion decreased (p < .05) between 1800 and 2100 m a.s.l. by 82.7%.
Site significantly affected every group of FA (Table ). Interactions found with total CLA resembled those in C18:2 c9t11. The iso-to-anteiso FA ratio was higher (p < .05) for experienced than inexperienced multiparous cows at 2100 m a.s.l. Moreover, it increased (p < .05) in experienced multiparous cows and inexperienced primiparous cows from 600 to 1800 m a.s.l. The SFA proportion (p < .05) decreased by 5.2% from lowland to alpine pastures. Proportions of PUFA, total CLA, n-3 and n-6 FA increased (p < .05) by 28.8, 23.2, 44.5 and 24.1% between 600 and 1800 m a.s.l., respectively, and returned to an intermediate level from 1800 to 2100 m a.s.l. for PUFA and total CLA (p < .05). Between 1800 and 2100 m a.s.l., n-6 FA proportion returned to its previous level at 600 m a.s.l. The n-6-to-n-3 FA ratio decreased (p < .05) by 32.4% between 600 and 2100 m a.s.l. The ratio C14:1 c9 to C14:0 was higher in milk of inexperienced cows (0.119 ± 0.002) than in that of experienced cows (0.111 ± 0.003, p < .05).
Table 6. Effect of experience, site and parity on the groups of fatty acids (FA) in total milk fat, g/100 g.
Discussion
Effects of alpine grazing
The present study involves a common type of alpine transhumance system, where cows are moved to progressively higher altitudes. This scheme offers grass of sufficient quality throughout the season due to the delayed vegetation growth at higher altitude. The nutrient composition of the forages was quite similar at the three sites except that the alpine grass contained more lignified fibre, consistent with the botanical composition found. The phenology of the plants may also have contributed, but this was not recorded in the present study. Indeed, highly digestible grasses make up a smaller proportion in alpine swards (Jeangros et al. Citation1999).
Most variables measured in the cows changed between sites. Ingestion time was lower than previously observed by O’Driscoll et al. (Citation2010) with Holstein-Friesian cows, which most probably had higher intake capacity and energy requirements than Aosta Red Pied in the present study. It increase when cows were moved from 600 to 1800 m a.s.l. likely resulted from the extension of the time on pasture, as rearing system changed between these sites. Indeed, with cows milked once a day, the same authors showed that the extra time allowed on pasture resulted in an increased grazing time. Romanzin et al. (Citation2018) showed that a low level of supplementation increased the grazing time on low-energetic heterogeneous pastures, which could be another possible explanation. The decline in ingestion time at 2100 m a.s.l. might indicate that cows spent less time searching for the more palatable grasses. Moreover, the cows’ energy requirements declined with the advancing stage of lactation, which probably diminished their ingestion time, too. Differences between pastures in lignified fibre were obviously too small to cause rumination time to differ. Cows with higher milk yield typically express a substantial decline in milk yield when moved to high altitude (Leiber et al. Citation2006). Causes are the limited nutrient supply by the grass and the extra energy needed to cope with hypoxia and climbing on steep slopes (Leiber et al. Citation2006). Also in the present study, milk yield declined to some extent from 1800 to 2100 m a.s.l. However, this decrease was smaller than that reported from other studies with ARP (Renna et al. Citation2010; Gorlier et al. Citation2012). The transfer from 600 and 1800 m a.s.l. was tolerated by this robust breed, even without milk yield decline going beyond that caused by the progressing lactation, as DIM-adjusted values show. At 2100 m a.s.l. the harsher conditions (higher temperature difference between night and day, hypoxic environment, topography) may have prevented the full exhibition of the milk yield potential. Under stress like changing the milking environment, shorter milking times and changes in milk fat content were expected, as milk ejection may be inhibited (Wellnitz and Bruckmaier Citation2001). In the present study, this was only the case for inexperienced cows at the lowland site. A confounding effect by changing milking equipment with different properties (e.g. vacuum strength) when moving from lowland to alpine pastures could not be excluded.
In the lowland, milk fat content was unexpectedly low compared to Renna et al. (Citation2014). However, to our knowledge, previous measurements only featured hay. In the present study, cows in the lowland grazed on young and less fibrous grass, with fibre being the substrate for milk fat synthesis (Chilliard et al. Citation2001). The milk composition found on alpine pastures was consistent with previous studies using ARP (Battaglini et al. Citation2005; Renna et al. Citation2010), even though grazing system and location were not the same. The milk fat content continuously increased during the alpine sojourn, because of the increasing need for body fat mobilisation (Kreuzer et al. Citation1998) and the elevated fibre content of the grass. The continuous decrease in lactose was likely only an effect of the progressive lactation (Dillon et al. Citation2003). Different from previous studies (Leiber et al. Citation2005, Citation2006), protein and urea contents remained unaffected by the alpine sojourn in the present experiment. This may be related to the comparably low requirements of the ARP cows for energy, the most limiting factor for milk protein synthesis. The effect of change from lowland to alpine grazing in FA profile was the subject of several previous studies (e.g. Bugaud et al. Citation2001; Leiber et al. Citation2005), and, more specifically, studies with ARP cows (Battaglini et al. Citation2003, Citation2005; Renna et al. Citation2010). Consistent with that, the alpine milk fat was characterised by low SFA and high MUFA, PUFA and CLA proportions. The n-6-to-n-3 FA ratio was also changed towards n-3 FA as expected (Leiber et al. Citation2005), although long-chain n-3 FA were present in minutes amounts. Elevated CLA, trans C18:1 FA (especially C18:1 t11) and n-3 FA, and less C18:0 suggest that PSC in alpine forage might have inhibited ruminal biohydrogenation of part of the PUFA (Buccioni et al. Citation2012). Proportions of branched-chain iso- and anteiso-FA, indicative of rumen microbial activity, were higher in milk fat in the alpine period, a shift which is further enhanced by diets poor in starch and rich in fibre (Zhang et al. Citation2017).
Effect of experience
Cows start learning how to graze as young animals, mostly by observing older or more experienced animals (Costa et al. Citation2016). However, this opportunity was not given to calves in the current ARP husbandry system. Cows have a long memory, leading them to remember where older animals were grazing and on what (Fraser and Broom Citation1997). Touching the grass and exploring are important for feeding behaviour, too (Distel and Provenza Citation1991; Currie Citation1995). It was reported that older cattle spend less time grazing than younger ones because of their eating experience, but still ingest for a longer time when arriving on a new pasture because they spend more time exploring (Krysl and Hess Citation1993). However, this was not the case in the present study. Eventually, feeding behaviour depends on prior bad eating experience and recognition of the presentation of plants as forages (Distel et al. Citation1995; Provenza et al. Citation2015). Moreover, cows have cognitive and physiologic abilities to determine whether they achieved their nutritional and energetic needs (Provenza et al. Citation2015). This can also affect their grazing behaviour.
Taken together, previous experience on specific alpine pastures could, therefore, result in more efficient feeding behaviour. Animals originated from two different farms, but their genetic background was similar. Thus, differences in milk yield and composition can be attributed to the experience with experimental conditions and associated behavioural differences. Cows of both groups were found to be similar in feeding behaviour and milk yield on alpine pastures. The only direct effect of experience was a tendency towards a reduced rumination time. This could indicate the ability of experienced cows to select plants with higher digestibility. However, this needs to be confirmed by a more in-depth analysis of the feeding behaviour. Finally, the results indicate that when using robust breeds in extensive alpine farming systems, previous experience would not provide a general medium-term advantage. Experience could be more important directly after transhumance, but its effect would be confounded with that of the walk or transport to the highland.
Milking dynamics were affected by experience, but this was already the case in the lowlands. Since the inexperienced cows, different from the experienced cows, did not know milking equipment and staff in the farming system investigated, the effects observed were likely due to the novel milking procedure and not to the lack of pasture-related experience, even though this was determined after an adaptation period. Overall, milk flow data indicate that milking time of experienced cows was longer compared to inexperienced cows at the lowland site. As all milking machines were left on each cow for the same length of time, this could have led to overmilking of the inexperienced cows. The breeder of inexperienced animals may have selected cows with short milking time, which might have intensified the problem. Overmilking is detrimental to udder health (Edwards et al. Citation2013). This could also explain why SCS and incidence of clinical mastitis were subsequently higher in inexperienced cows in both alpine periods. Mastitis and high SCS can also be linked to the stress of animals arriving on a new pasture, as alpine grazing promotes somatic cell count (Lamarche et al. Citation2000; Leiber et al. Citation2005). Furthermore, Coulon et al. (Citation1998) demonstrated that physical motion increases SCC in milk of untrained cows. However, as strip grazing was applied, only small effects of the latter were expected.
Data on milk composition indicate that the feed of inexperienced and experienced cows differed in composition. The lower casein content in milk of inexperienced compared to experienced cows at 1800 m a.s.l. points towards a lower energy supply and, thus, metabolisable protein (Leiber et al. Citation2005). The higher urea content in milk of inexperienced cows was unexpected. Possibly their previous grazing experience could have had an effect, as they were used to grazing lowland pastures rich in protein-rich plants. Although the strip grazing technique reduces the possibility to select among plant species, cows still may have been able to recognise and select species-rich in protein through ‘feed hedonics’ (Villalba et al. Citation2015). Further studies on ingested feed composition would be needed to confirm this assumption. Effects of experience on milk FA profile consisted of three categories: experience enhanced proportions of some short- and medium-chain FA, reduced proportions of mono-unsaturated medium-chain FA, and enhanced those of several C18:1 trans isomers. The latter points towards a specific selection by experienced cows towards forages rich in PSC. Relationships between phenolic compounds and ruminal biohydrogenation were repeatedly shown (Vasta et al. Citation2009; Kälber et al. Citation2011; Willems et al. Citation2014). However, herb selection was not monitored in the present experiment. Apart from that, higher ΣC16:1 and ΣC17:1 proportions in milk fat of inexperienced cows, together with the higher C14:1 c9-to-C14:0 ratios, point towards a higher Δ9-desaturase activity in the mammary gland (Rutkowska et al. Citation2012).
Distinction between site-specific and age experience
Cows are able to learn through observation of older animals (Costa et al. Citation2016), which is a different experience from that acquired by grazing a specific site. In the present study, primiparous cows could be considered less experienced than multiparous cows because of their young age and the fact that calves and heifers are not kept together with more experienced animals before first calving. We aimed at testing if the age experience was more important than the site-specific one. Interactions of experience with parity (and together with site) were rare, suggesting that the site-specific experience obtained before first calving was equally valuable as that obtained as a dairy cow. One exception was that only inexperienced primiparous cows responded to the new situation at 1800 m a.s.l. They increased their ingestion time in response to the new pasture. However, it is unclear if they increased it more than what would have been needed to cover the extra energy and nutrients necessary to maintain milk yield at this altitude.
Conclusions
No clear advantage of previous experience with distinct alpine pastures used in a traditional extensive transhumance system and those associated with age (multiparous vs. primiparous cows) was found on feeding behaviour. Several medium-term effects on milking characteristics and milk composition were observed. However, these effects were much lower in general than those caused by the transhumance to alpine pastures. In addition, the new farming environment rather than the lack of habituating to the specific alpine pastures caused part of the stress for inexperienced cows. As the milking characteristics demonstrate, familiarisation to new staff and milking equipment already on lowland pastures could be effective. In the transhumance system investigated, the dispersal of feed across inhomogeneous alpine pastures had been minimised by strip grazing. In all other approaches, experience may have a greater importance. Future studies are needed to determine the importance of experience during transport and the actual days of transition and to compare autochthonous breeds with high-yielding genotypes.
Ethical approval
For severity zero, only a communication at the local veterinarian office was necessary. All procedures of the experiment were excluded from further authorization of the animal welfare committee.
Acknowledgements
Authors wish to thank the technical staff from the IAR for their help and assistance. Thanks to D. André for allowing his cows to participate in the experiment.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Ali AKA, Shook GE. 1980. An optimum transformation for somatic cell concentration in milk. J Dairy Sci. 63:487–490.
- Battaglini L, Mimosi A, Malfatto V, Lussiana C, Bianchi M. 2005. Milk yield and quality of Aosta cattle breeds in Alpine pasture. Ital J Anim Sci. 4:224–226.
- Battaglini L, Mimosi A, Malfatto V, Robin Preillan A, Bianchi M. 2003. Management and productive aspects of a dairy herd in a Valle d’Aosta alpine pasture. Ital J Anim Sci. 2:293–295.
- Buccioni A, Decandia M, Minieri S, Molle G, Cabiddu A. 2012. Lipid metabolism in the rumen: new insights on lipolysis and biohydrogenation with an emphasis on the role of endogenous plant factors. Anim Feed Sci Tech. 174:1–25.
- Bugaud C, Buchin S, Coulon JB, Hauwuy A, Dupont D. 2001. Influence of the nature of alpine pastures on plasmin activity, fatty acid and volatile compound composition of milk. Lait. 81:401–414.
- Chilliard Y, Ferlay A, Doreau M. 2001. Effect of different types of forages, animal fat or marine oils in cow’s diet on milk fat secretion and composition, especially conjugated linoleic acid (CLA) and polyunsaturated fatty acids. Livest Prod Sci. 70:31–48.
- Costa JHC, Costa WG, Weary DM, Machado Filho LCP, von Keyserlingk MAG. 2016. Dairy heifers benefit from the presence of an experienced companion when learning how to graze. J Dairy Sci. 99:562–568.
- Coulon JB, Pradel P, Cochard T, Poutrel B. 1998. Effect of extreme walking conditions for dairy cows on milk yield, chemical composition, and somatic cell count. J Dairy Sci. 81:994–1003.
- Currie WB. 1995. Structure and Function of Domestic Animals. New York, USA: CRC Press.
- Daget P, Poissonet J. 1971. Une méthode d’analyse phytosociologique des prairies. Ann Agron. 22:5–41.
- Dillon P, Buckley F, O’Connor P, Hegarty D, Rath M. 2003. A comparison of different dairy cow breeds on a seasonal grass-based system of milk production: 1. Milk production, live weight, body condition score and DM intake. Livest Prod Sci. 83:21–33.
- Distel RA, Laca EA, Griggs TC, Demment MW. 1995. Patch selection by cattle: maximization of intake rate in horizontally heterogeneous pastures. Appl Anim Behav Sci. 45:11–21.
- Distel R, Provenza F. 1991. Experience early in life affects voluntary intake of blackbrush by goats. J Chem Ecol. 17:431–450.
- Edwards JP, O'Brien B, Lopez-Villalobos N, Jago JG. 2013. Overmilking causes deterioration in teat-end condition of dairy cows in late lactation. J Dairy Res. 80:344–348.
- Elgersma A. 2015. Grazing increases the unsaturated fatty acid concentration of milk from grass-fed cows: a review of the contributing factors, challenges and future perspectives. Eur J Lipid Sci Technol. 117:1345–1369.
- Farruggia A, Pomiès D, Coppa M, Ferlay A, Verdier-Metz I, Le Morvan A, Bethier A, Pompanon F, Troquier O, Martin B. 2014. Animal performances, pasture biodiversity and dairy product quality: how it works in contrasted mountain grazing systems. Agr Ecosyst Environ. 185:231–244.
- Fraser AF, Broom DM. 1997. Farm Animal Behaviour and Welfare. Wallingford, UK: CAB International.
- Gorlier A, Lonati M, Renna M, Lussiana C, Lombardi G, Battaglini L. 2012. Changes in pasture and cow milk compositions during a summer transhumance in the western Italian Alps. J Appl Bot Food Qual. 85:216–223.
- Iussig G, Renna M, Gorlier A, Lonati M, Lussiana C, Battaglini LM, Lombardi G. 2015. Browsing ratio, species intake, and milk fatty acid composition of goats foraging on alpine open grassland and grazable forestland. Small Rum Res. 132:12–24.
- Jeangros B, Scehovic J, Troxler J, Bachmann HJ, Bosset JO. 1999. Comparison of the botanical and chemical characteristics of grazed pastures, in lowlands and in the mountains. Fourrages. 159:277–292.
- Kälber T, Meier JS, Kreuzer M, Leiber F. 2011. Flowering catch crops used as forage plants for dairy cows: influence on fatty acids and tocopherols in milk. J Dairy Sci. 94:1477–1489.
- Kreuzer M, Langhans W, Sutter F, Christen RE, Leuenberger H, Kunz PL. 1998. Metabolic response of early-lactating cows exposed to transport and high altitude grazing conditions. Anim Sci. 67:237–248.
- Krysl LJ, Hess BW. 1993. Influence of supplementation on behavior of grazing cattle. J Anim Sci. 71:2546–2555.
- Lamarche A, Martin B, Hauwuy A, Coulon JB, Poutrel B. 2000. Evolution of milk somatic cell count of cows grazing an alpine pasture according to the infection of udder by pathogens. Ann Zootech. 49:45–54.
- Leiber F, Kreuzer M, Leuenberger H, Wettstein H-R. 2006. Contribution of diet type and pasture conditions to the influence of high altitude grazing on intake, performance and composition and renneting properties of the milk of cows. Anim Res. 55:37–53.
- Leiber F, Kreuzer M, Nigg D, Wettstein H-R, Scheeder MRL. 2005. A study on the causes for the elevated n-3 fatty acids in cows’ milk of alpine origin. Lipids. 40:191–202.
- Lopes F, Coblentz W, Hoffman P, Combs D. 2013. Assessment of heifer grazing experience on short-term adaptation to pasture and performance as lactating cows. J Dairy Sci. 96:3138–3152.
- Niero G, Koczura M, De Marchi M, Currò S, Kreuzer M, Turille G, Berard J. 2018. Are cheese-making properties of dual-purpose cattle impaired by highland grazing? A case study using Aosta Red Pied cows. Ital J Anim Sci. 1. doi:10.1080/1828051X.2018.1443289
- Nydegger F, Keller M, Gygax L, Egli W. 2011. Capteur de mastication pour vaches laitières: Saisie automatique de l'activité de mastication et d'alimentation pour le contrôle sanitaire. ART-Bericht. 748:1–8.
- O’Driscoll K, O’Brien B, Gleeson D, Boyle L. 2010. Milking frequency and nutritional level affect grazing behaviour of dairy cows: a case study. Appl Anim Behav Sci. 122:77–83.
- Provenza FD, Gregorini P, Carvalho PCF. 2015. Synthesis: foraging decisions link plants, herbivores and human beings. Anim Prod Sci. 55:411–425.
- Renna M, Cornale P, Lussiana C, Battaglini LM, Turille G, Mimosi A. 2014. Milk yield, gross composition and fatty acid profile of dual-purpose Aosta Red Pied cows fed separate concentrate-forage versus total mixed ration. Anim Sci J. 85:37–45.
- Renna M, Lussiana C, Malfatto V, Mimosi A, Battaglini LM. 2010. Effect of exposure to heat stress conditions on milk yield and quality of dairy cows grazing on Alpine pasture. Proc. 9th European IFSA Symposium, Vienna, Austria, 4-7/07/2010, pp. 1338–1348.
- Regione Autonoma Valle dAosta. 2006. Elenco regionale dei laboratori che effettuano analisi ai fini dell’autocontrollo per le industrie alimentari. Report No. DGR N. 1025. Available at www.regione.vda.it
- Romanzin A, Corazzin M, Piasentier E, Bovolenta S. 2018. Concentrate supplement modifies the feeding behavior of Simmental cows grazing in two high mountain pastures. Animals. 8:76.
- Rutkowska J, Adamska A, Bialek M. 2012. Fatty acid profile of the milk of cows reared in the mountain region of Poland. J Dairy Res. 79:469–476.
- Silbernagel H. 2002. Alpsommer. Miesbach, Germany: Bergemann and Mayr.
- Sturaro E, Marchiori E, Cocca G, Penasa M, Ramanzin M, Bittante G. 2013. Dairy systems in mountainous areas: farm animal biodiversity, milk production and destination, and land use. Livest Sci. 158:157–168.
- Vasta V, Mele M, Serra A, Scerra M, Luciano G, Lanza M, Priolo A. 2009. Metabolic fate of fatty acids involved in ruminal biohydrogenation in sheep fed concentrate or herbage with or without tannins. J Anim Sci. 87:2674–2684.
- Villalba JJ, Catanese F, Provenza FD, Distel RA. 2012. Relationships between early experience to dietary diversity, acceptance of novel flavors, and open field behavior in sheep. Physiol Behav. 105:181–187.
- Villalba JJ, Provenza FD, Catanese F, Distel RA. 2015. Understanding and manipulating diet choice in grazing animals. Anim Prod Sci. 55:261–271.
- Wellnitz O, Bruckmaier RM. 2001. Central inhibition of milk ejection. Livest Prod Sci. 70:135–140.
- Willems H, Kreuzer M, Leiber F. 2014. Alpha-linolenic and linoleic acid in meat and adipose tissue of grazing lambs differ among alpine pasture types with contrasting plant species and phenolic compound composition. Small Rumin Res. 116:153–164.
- Zendri F, Ramanzin M, Bittante G, Sturaro E. 2016. Transhumance of dairy cows to highland summer pastures interacts with breed to influence body condition, milk yield and quality. Ital J Anim Sci. 15:481–491.
- Zhang Y, Liu K, Hao X, Xin H. 2017. The relationships between odd- and branched-chain fatty acids to ruminal fermentation parameters and bacterial populations with different dietary ratios of forage and concentrate. J Anim Physiol An N. 101:1103–1114.