
Abstract
The aim of this trial was to investigate on the growth and meat quality of Padovana male chickens with different plumage varieties, chamois (PC - light brown feathers with white edge), silver (PS - white feathers with black edge), and their cross. The body weight of PC during the growth period was higher (p < .01) than PS, and it was 1.7 and 1.5 kg, respectively, at 126 d of age. At slaughter, PC showed higher weight of carcase (p < .05), breast and total fleshiness (breast, wings and legs) (p < .01), and thigh meat:bone ratio (p < .05). PS showed higher shanks weight on carcase weight (p < .01), Ilio tibialis a* value (p < .01), water losses (p < .01) and shear force (p < .05) in breast meat than PC. Crossing PC males to PS females gave birds with white (Cross-W) and silver (Cross-S) plumage (3:1 ratio, respectively). The offspring genotypes showed similar body weight, and almost all slaughtering, carcase and meat quality traits studied. Cross-W and Cross-S showed significantly higher final body weight, breast and leg weight, total fleshiness and thigh meat:bone ratio than PS. For the Padovana breed, the plumage colour can involve productive and slaughtering performance, and carcase and meat quality, throughout the growing period. At 18 weeks of age, the Padovana male chickens show body weight and carcase fleshiness similar to that of a hybrid laying hen belonging to a light strain.
Padovana is an historic Italian chicken breed with a slow growth rate; it is slaughtered broadly later than hybrid strains and it is important to know the age for achieving favourable slaughtering performance and good carcase quality.
Padovana chicken breed has been selected in some plumage colours and the growth rate, slaughtering performance and carcase quality can significantly differ according to them.
Crossing chickens with two different plumage colours can exert a positive heterosis effect on productive and carcase quality traits.
Highlights
Introduction
Northern Italy has a long tradition in poultry production, given the presence of a wide plain of alluvial origin crossed by Po river and a large production of cereals as wheat and corn. Padovana breed is a local Italian breed; its name origins from the city of Padova, in Veneto region in Northern Italy, where it was reared since 1500 (Aldrovandi Citation1599). It was initially reared in the houses of Venetian aristocracy that was attracted by its particular head morphology. Padovana head has a frontal bony protuberance with a variable number of openings as well as a variable openings size according to the plumage variety (Verdiglione and Rizzi Citation2018) with a big tuft that partially cover eyes. Sexual dimorphism involves many body traits, also the comb that can be absent or less developed in females and generally consists of two little horns or has an irregular shape in males. Different plumage varieties exist, single colour or a combination of two colours, as the golden, silver and chamois variety with brown, white and light brown feathers, respectively, with black (golden and silver varieties), and white (chamois variety) edge. Padovana breed has grey shanks and the eggshell is white (Veneto Agricoltura Citation2004). It can be used as an ornamental hen for these characteristics, but it is also used for eggs production and gastronomic preparations given the excellent quality of its meat (Verdiglione and Cassandro Citation2013).
It is a slow-growing breed, and the chickens are usually slaughtered not before the pubertal age, depending mainly on gender (Cassandro et al. Citation2015; Rizzi Citation2018; Tasoniero et al. Citation2018).
The goal of this trial was to compare the growth, the slaughtering performance, and the carcase fleshiness and meat quality of Padovana male chickens belonging to two plumage varieties and their cross in order to assess differences between the groups and the heterosis effect until 18 weeks of age.
Materials and methods
Animals and rearing conditions
The birds were handled according to the principles stated in EC Directive 86/609/EEC. The trial was carried out on Padovana male chickens belonging to different plumage varieties, chamois (Padovana camosciata – PC) and silver (Padovana argentata – PS) plumage and the cross between PC males and PS females. The cross birds showed two plumage colours, white plumage (Cross-W) and silver plumage (Cross-S) at a ratio of 3:1, respectively. PC has light brown feathers with white edge, PS has white feathers with black edge; the cross has a completely white plumage or a silver plumage similar to that of PS. The birds came from an institute of Padovana breed conservation located in Padua. The chicks of the four groups were reared from 1 d until 126 d (18 weeks) of age under the same environmental conditions and prophylaxis procedures (Newcastle disease, Marek and Infectious bronchitis vaccines). The birds were kept in a breeder house from the hatching until the end of the trial. One–day old chicks of each strain were sexed (by cloacal inspection) and reared separately on wooden litter (at a mean stocking density of a final body weight of 3.3 kg/m2) from the start until the end of the trial. The chicks were fed ad libitum the same feed (Table ) and water. Formulation and composition did not change throughout the trial; the feed form was administered as crumbled for the first 8 weeks of age, and then in pelleted form. The trial started in May and lasted until September. The birds (PC = 36 birds; PS = 23 birds; Cross-W = 26 birds; Cross-S = 18 birds) were weighed from the 3rd week of age (after the first two weeks of life, when the mortality rate was high for some genotypes, as indicated later in the discussion) every 2 weeks until 70 d of age, and then every month.
Table 1. Formulation and chemical composition of the feed.
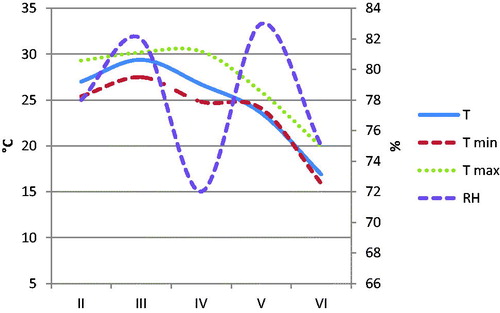
Indoor temperature and relative humidity were checked during the experimental period (Figure ); the photoperiod was natural, and it decreased during the trial according to the season. Throughout the first 4 weeks of life, the chicks were kept under infra-red lamps and the temperature under them was gradually lowered from 33 to 24 °C, and then it changed according to the seasonal values.
Figure 1. Environmental temperature (T) and relative humidity (RH) patterns according to the age of the chickens. Age: is referred to the weeks of life; II (5–7th week); III (8–10th weeks); IV (11–13th week); V (14–16th week); VI (17–18th week).
Slaughtering procedures
At 126 d of age, the birds (7–10 birds/group) were weighed and brought to the slaughter-house. Feed was withdrawn 12 h before slaughtering; the birds, previously weighed, were electronically stunned, killed by exsanguination, plucked, and eviscerated (intestines; perivisceral, perineal and abdominal fat; gall bladder; oesophagus; full crop; proventriculus; and spleen). Cold carcase (after air chilling at 0 °C for 90 min and then stored at 4 °C) weights were recorded. From the refrigerated carcase, the head and neck, shanks, and giblets (heart, liver and gizzard) were removed to obtain the ready-to-cook carcase (WPSA Citation1984). Successively, breasts, thighs, drumsticks and wings were weighed. The shanks, and the head and neck were also weighed. The wing (drumette - humerus, surrounding musculature and skin; wingette – radius and ulna, surrounding musculature and skin; tip – carpal bones, cartilage and skin) and the leg (thigh and drumstick), after weighing, were dissected and the weight of skin, muscles and bones were recorded.
pH and physical parameters of breast, wing and leg
On the breast muscle, Pectoralis major, at 48 h post mortem, the ultimate pH by a Delta Ohm HI-8314 pHmeter (Delta Ohm, Padova, Italy), and Crison electrode (Crison, Barcelona, Spain) was recorded. Colour measurements on breast skin and muscle, Pectoralis major, and on thigh skin and muscle, Ilio tibialis, were recorded by a Minolta Chroma metre CR-300 tristimulus analyzer (Minolta Corp., Ramsey, NJ, USA) (CIELAB colour space model, Commission International de l’Eclairage Citation1978). The CIELAB colour space model was chosen to numerically describe the colour parameters. Lightness (L*) is the amount of incident light that a surface reflects; −a* values represent green and + a* values represent red; −b* values represent blue and + b* values represent yellow.
The right skinless breasts were weighed and frozen at −20 °C for further analyses concerning some rheological traits. Given that Padovana chickens for meat production are usually reared from the spring until the autumn months, the carcases are often frozen to ensure culinary preparations all through the year. After 12 d, the breasts were removed from the freezer, placed on trays, and thawed in a commercial refrigerator at 3–4 °C for 24 h. The breast muscles were blotted dry, and then cooked in water at 75 °C for 60 min inside sealed bags. After 15 min of cooling under running water, the fillets were dried with paper.
For the evaluation of the thawing losses [(frozen weight – thawed weight)/(frozen weight) × 100] and cooking losses [(uncooked weight − cooked weight)/(uncooked weight) × 100] the breasts were weighed before and after thawing and cooking. Then, shear force was performed on three cylindrical cores (1.13 cm of diameter) of each breast-cooked sample, by using a TA-HDi Texture Analyzer (Stable Macro System, London, UK) with a Warner-Bratzler shear probe (10 N load cell, crosshead speed of 2 mm/s). The peak shear force, as average from the three replicates, was taken as the final shear force value.
Chemical analyses of feed and meat
The crumbled feed was ground (1-mm mesh screen, Grindomix ZM 200, Retsch Gmbh, Haan, Germany) and the raw meat of left breasts were homogenised in a homogeniser (Grindomix GM200, Retsch GmbH, Haan, Germany), stored at −20 °C for further analyses. After 12 d, they were chemically analysed for dry matter (method 950.46; AOAC Citation2000), crude protein (method 981.10; AOAC Citation2000), lipid (method 991.36; AOAC Citation2000) and ash (method 920.153; AOAC Citation2000). Total lipids, in the breast meat samples, were extracted according to the method of Folch et al. (Citation1957).
Statistical analysis
All the data were evaluated by using ANOVA according to a general linear model that considered genotype as fixed effect (PROC GLM) following procedure of SAS (Version 9.3, SAS Institute Inc., Cary, NC, USA). Differences among groups were detected by Tukey-Kramer’s test at a significance level of p < .05. Heterosis was calculated using the following formula: [Cross-W or Cross-S − (PC + PS)/2]/[(PC + PS)/2] × 100 for all variables (except the colour data) and it is discussed only for the variables with statistically significant differences.
Results and discussion
Growth, slaughtering performance and carcase yield
Crossing PC males to PS females produced chicks with two different plumage colours, one completely white and the other silver. The white:silver plumage ratio for the cross birds was more favourable for the first plumage colour: at hatching, a higher percentage of birds with white plumage (76%) than that of silver plumage (24%) was observed. The factors, of genetic or environmental origin, determining a different ratio of plumage colour, also in female chickens (Rizzi Citation2018), are still unknown, as well as it is not clear whether this ratio origins from the layed eggs or the hatched eggs (Collins and Hubbard Citation1958; Conway and Martin Citation2000; Yilmaz et al. Citation2011).
The daily gain is summarised in Table : the results indicate a lower (p < .01) growth for the PS chickens when compared to PC, until the 8th week of life, and then similar, whereas Cross-W did not differ from Cross-S at all the ages considered. A different growth was observed for Cross-W in comparison to PC at 5–6th (lower, p < .05) and 9–10th (higher, p < .01) weeks of age, and to PS (higher, p < .01) for the first period of life until 70 d of age. Cross-S showed a lower (p < .01) body gain than PC until 5–6th week of age, and then higher until the 18th week (p < .05) and it showed a growth rate higher than that of PS from 7–10th week (p < .05) of age. The heterosis was constantly positive for Cross-W with the highest values at 9–10th week of life (+18.8%), whereas for Cross-S it was negative until the 5–6th weeks of life (−9.0%) and then positive with values of +19.6% and +15.9% at 9–10th and 15–18th week of life, respectively.
Table 2. Daily growth and heterosis according to the age of the chickens.
In Table , the body weight of the chickens is reported. The two parental groups showed different body weights according to the plumage colour: throughout the rearing period the PS chickens showed lower (p < .01) body weights than those of PS. No difference was observed between Cross-W and Cross-S. A comparison between each offspring group and parental genotype, for the body weight, showed that Cross-W was similar to PC, but it was constantly higher (p < .01) than PS. The body weight of Cross-S was lower than PC until 10 weeks (p < .05) of age and then similar; it was higher than PS from the 8th week (p < .01). Crossing PC to PS exerted a positive heterosis on the two cross genotypes: the live body weight was positively affected throughout the growing period for Cross-W (on average +8.6%), whereas for Cross-S it was initially negative (−10.5%) and then increased until the slaughter age (+6.1%). It is well remembering that at 126 d of age (18 weeks) the PC birds had reached 80% adult body weight as indicated by Veneto Agricoltura (Citation2004), whereas for the PS genotype no comparison is possible because there is a lack of literature information on its adult body weight. Indications on gonads development can be useful when considering the body growth: the birds of this trial at 18 weeks of age showed a very low testicular development, with a testes incidence of 0.03–0.2% body weight, indicating that the males were distant from the pubertal age and had not reached the adult body weight. The body weight at 18 weeks of age was 1.7 kg for the PC and the cross birds, whereas PS showed a body weight of 1.5 kg. Nowadays, commercial hybrids for broiler production are slaughtered from 2.2 kg at 35 d of age (Petracci et al. Citation2015) and this indicates that the Padovana chickens can be slaughtered not before 4 months of age for achieving a commercial size and acceptable slaughtering performance, comparable to those of laying hen hybrid strains (Rizzi et al. Citation2007).
Table 3. Body weight and heterosis according to the age of the chickens.
For the Padovana chickens, the more critic period of growth seems to be that concerning the feathers growing. The reason of this response in this physiological phase is not clear as different factors can affect the daily growth of the animals such as genetic assessment or interaction with environmental conditions or feeding. Developing and growing feathers need nutritional requirements (especially tyrosine and carotenoids, and microelements) according to the plumage colour, and a variety of genes and mutations are involved (Mills and Patterson Citation2009). Birds have only one pigment cell type, the melanocyte; these cells can produce either eumelanin (black/brown) or pheomelanin (yellow/red) and can switch rapidly between the syntheses of these two pigment types. Despite the colours produced by birds, the feather pattern resulting in bars, stripes or spots is controlled exclusively by genes and melanin production, but bird and mammalian models of stripe formation are lacking, whereas good fish and amphibian models exist (Mills and Patterson Citation2009). The feather lacing in silver plumage is due to the presence of gene that affects the distribution of pigment within the feather, thus showing a black border (Nesheim et al. Citation1979). Furthermore, in chickens whose mating occurs in polygamy, it is important iridescence, the most common type of structural colour, important as a source of UV reflectance. Changes of a structural colour depend on the angle from which it is viewed, and they are produced by myriad patterns of alternating layers of keratin, air and melanin in barbules (Mills and Patterson Citation2009). Feather growth with such a structure may be longer than those of feathers with more simple structure so it is likely that silver males showed a lower daily growth until 70 d of life also for this reason. Not recent indications stated that some genes for plumage colour and feathering can be lethal influencing the hatching ratio (Collins and Hubbard Citation1958) and can lower the growth rate of chickens with silver plumage (Jaap and Grimes Citation1956). In this trial the mortality rate until 18 weeks of age averaged 5% for PC, 7% for cross-W and 18% for cross-S, whereas for PS was 35%, with a great number of deaths during the first weeks of life, as previously indicated. It is not clear which endogenous factors could have induced such a high mortality rate in PS even if there is a reasonable hypothesis that a genetic component could have been involved as stated by Collins and Hubbard (Citation1958). These authors observed in chickens with barred plumage a lethal effect due to the presence of gene for black plumage and a possible its interaction with unknown factors. A similar mortality rate was observed in females belonging to the same genetic groups (Rizzi Citation2018).
The daily growth of the four groups is a growth typical of slow-growing strains; the maximum value, under 20 g/d, was reached between 77 and 97 d and then it decreased. It is worth remembering that the environmental temperature was high throughout the trial (Figure ), so this condition could have induced a lower growth. The daily growth of Padovana breed is well modelled by the Gompertz equation (Rizzi et al. Citation2013). As previously stated, PS and PC showed a different body weight until 22 weeks of age, but their asymptotic final body weight was not statistically different, even if in PC was 13% higher than PS. Furthermore, in PS the inflection point was reached 4–9 d later than PC and at a lower body weight, but without any statistical significance (Rizzi et al. Citation2013). In the same previous study (Rizzi et al. Citation2013) the Padovana cross bird group (PC × PS), including chickens with white and silver plumage at the same ratio of this trial, did not statistically differ from the parental lines for the body weight, even if with values more favourable in comparison to PS.
The slaughtering performance of the four groups is summarised in Table . The PC carcase weight was higher (p < .05) than PS, but the dressing percentage was similar. The commercial cuts differed between the two groups for the breast (p < .01) and legs (p < .05) weights, that were higher in PC, whereas the wings weight was similar. The PS birds showed higher percentage of shanks (p < .01) on carcase weight, but similar percentage of head and neck.
Table 4. Slaughtering performance.
The two offspring genotypes showed similar data, with an exception for the percentage of head and neck on carcase that was higher (p < .05) in Cross-W than in Cross-S. The two offspring groups showed a lower (p < .05) dressing percentage than PS, but higher breast (p < .01) and legs (p < .05; p < .01) weight. Cross-S showed a lower incidence on carcase of head and neck (p < .05) than PC and of shanks (p < .05) than PS. Heterosis was positive for the cold carcase weight (+3.7% and +2.9% in Cross-W and Cross-S, respectively) and negative for the dressing percentage (−3%). The heterosis effect was more considerable in Cross-S than in Cross-W for the breast weight (+14.2% versus +5.1%) and the head and neck % cold carcase (−11.2% versus +4.8%), whereas for the legs weight (+7.7% versus +7.0%) and shanks % cold carcase (−3.9% versus −3.5%) it was similar.
The carcase yield is shown in Table . As far as the ready-to-cook carcase conformation is concerned, the incidences of breast, wings and legs were similar between PC and PS and between the two offspring genotypes. A significant difference between Cross-S and PS was detected for the breast incidence that was higher (p < .05) in the cross birds. An evaluation of the fleshiness of the commercial cuts indicated a significant superiority of PC than PS for the legs (p < .05) and for the total weight (breast, wings and thighs and drumsticks) (p < .01), but not for their incidence on the ready-to-cook carcase. The two offspring groups were similar for all the fleshiness traits considered. Furthermore, Cross-W and Cross-S showed higher legs fleshiness (p < .05 and p < .01, respectively) and total fleshiness (p < .01 and p < .01, respectively) than PS; Cross-S showed a leg fleshiness higher (p < .01) than PC and a higher (p < .01) percentage of total fleshiness on ready-to-cook carcase than PS. Cross-S showed a higher heterosis value for the breast percentage on ready-to-cook carcase (+9.2% versus +1.7%). The heterosis for the fleshiness was constantly positive for all the variables considered: it was more relevant for the legs (+9.8% versus +8.1%) for Cross-S and Cross-W, respectively, and the total fleshiness, concerning the three commercial cuts and its percentage on the ready-to-cook carcase, weight showed heterosis values of +10.4%, +5.9% in Cross-S and +6.1%, +2.6%, in Cross-W, respectively. The slaughtering performance data showed typical values of a medium size purebred, with breast weighing less than those of hybrids (Petracci et al. Citation2015), and a lower fleshiness. The breast weight of the Padovana chickens of this trial and the breast incidence on body weight is similar to those of hybrids of 1957, slaughtered at 8 weeks of age (Petracci et al. Citation2015); the Padovana fleshiness (breast and legs) is slightly higher than that of Hy-Line White, a typical egg laying strain, of 1.7 kg body weight (Rizzi et al. Citation2007).
Table 5. Carcase yield.
In Table some characteristics of the wing are shown. The weight of the three portions of the wing, drumette, wingette and tip, did not differ between the groups as well as their meat:bone ratio. The leg traits are summarised in Table . As far as the leg portions are concerned, the two parental genotypes showed significant differences only for the thigh weight, with higher (p < .05) values in PC. No difference was seen between the offspring groups. The Cross-W and Cross-S groups showed a higher (p < .05) thigh weight in comparison to PS and Cross-W was higher (p < .05) also for the drumstick weight. The meat:bone ratio of the leg showed higher (p < .05) values in PC than in PS for the thigh and total leg, and it was similar between the two crosses. The Cross-W and Cross-S showed a higher meat: bone ratio for thigh (Cross-W: p < .05; Cross-S: p < .01) and for the total leg (Cross-W: p < .05; Cross-S: p < .01) than PS, as a consequence of a higher (Cross-S: p < .01) muscle incidence and a lower (Cross-W: p < .05, Cross-S: p < .01) bone incidence in thigh; the muscle and bone % on drumstick was similar for all the groups. In the offspring genotypes the heterosis effects were +7.8% and +6.9% for the weight of thigh and +3.9% and +7.0% for the weight of drumstick in Cross-S and Cross-W, respectively, and they showed the highest values for the meat:bone ratio of thigh (Cross-S: +24.8%, Cross-W: +15.1%) and leg (Cross-S: +19.2%, Cross-W: +13.6%). The heterosis effects involved, for the thigh, the muscle incidence and the femur incidence, particularly in the Cross-S group (+6.7% and −16.5% versus +3.8% and −11.5%, respectively). The meat:bone ratio of the leg of these birds is lower than those showed by hens belonging to Italian dual-purpose purebreds (Ermellinata di Rovigo and Robusta maculata) and to laying hen hybrids (Hy-Line white and Hy-Line brown) (Rizzi et al. Citation2007). The lower incidence of muscles in Padovana thigh, and especially in drumstick, can be due to the early age of the animals when compared to the adult birds, besides the genotype and rearing conditions.
Table 6. Wing traits.
Table 7. Leg traits.
Meat quality
The Table summarises the colour traits of breast and thigh, measured on skin and meat. As well as the breast skin and meat colours are concerned, L* (lightness), a* (redness) and b* (yellowness) did not substantially differ both between the two parental and the two offspring groups. Cross-S showed lower (p < .05) b* values than PS in the breast skin.
Table 8. Colour traits of breast and thigh.
For the thigh skin, the two parental genotypes and the two offspring groups did not show any difference. Cross-S showed a lower (p < .05) a* value than PC and PS. In the Ilio tibialis muscle, the a* value was higher (p < .01) in PS than in PC, and the two offspring genotypes were similar for all traits. Cross-W and Cross-S showed a* values lower (p < .01) than PS. A higher (p < .05) value of b* was observed in Cross-S than in PC.
Table summarised the chemical and rheological traits of the breast meat. The final pH of Pectoralis major muscle did not differ between the genetic groups. The two parental and the two offspring groups showed similar crude protein, total lipids and ash content of breast muscle. The ash content was lower (p < .05) in Cross-S than in PC group. The water retention capacity, identified in the thawing losses and in cooking losses, differed (p < .01) between PC and PS, that showed higher water losses (p < .01) as a consequence of higher thawing (p < .01) and cooking (p < .05) losses; PS meat showed a lower (p < .05) tenderness than PC. Heterosis was negative for the ash content (−1.7% and −3.7%) in Cross-W and Cross-S. Higher heterosis effects on thawing and cooking losses in Cross-W (+7.3%; −3.0%) than in Cross-S (+1.1%; −4.8%) were observed, and then a positive effect on the shear force in Cross-W (+8.0%) and a decrease in cross-S (−10.8%). Anyway, several factors, besides water losses, such as fibre resistance, sarcomere length and collagen morphology can also affect meat tenderness (Lepetit Citation2007). It is well noting that the skin colour parameters were differentiated between breast and thigh as well as the yellowness and redness values of breast and thigh muscle. Padovana birds showed skin and muscle colours largely different by those of commercial hybrids mainly due to the different skin pigmentation (Veneto Agricoltura Citation2004) and kind of muscular fibres (Verdiglione and Cassandro Citation2013). Padovana meat showed a good microbiological stability due to the final pH, thus avoiding different types of spoilage microorganisms that impair taste, flavour and appearance of meat (Barbut et al. Citation2008; Petracci et al. Citation2015).
Table 9. Chemical and rheological traits of breast meat.
Conclusions
The Padovana crested purebred has been selected in some phenotypes, involving mainly the plumage colour, but also the carcase yield and some physical and chemical traits of muscles. Chickens with chamois and silver plumage belonging to a population selected in Veneto region showed differences in carcase and meat traits at a slaughter age of 126 d. At this age PS showed a lower body weight than PC, a lower breast and hindleg muscles development, and a higher incidence on carcase of shanks. For the Padovana breed, a slow-growing local purebred, the plumage colour may be considered not only under a point of view of a fancy purpose but also under a productive purpose, as it can affect the slaughtering age at least throughout the growing period before the pubertal age. Crossing PC males to PS females produced chickens with two plumage colours, in different proportions, but with similar final body weight and carcase weight, and meat quality responses. The most considerable heterosis effect was observed on the carcase muscle development: in cross-S it involved the breast and leg cuts and it was higher than in Cross-W. Cross-W showed a considerable heterosis effect on the leg muscle development, particularly in thigh. The results of this trial are an original contribution to the study on Padovana chicken populations with different plumage varieties and their cross, and further research is needed to increase knowledge regarding the relationships between the plumage colour and protein (in particular, feather and muscle) and lipid (in particular, abdominal and muscular fat) syntheses and their effect on slaughtering and carcase yield and meat quality.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Aldrovandi U. 1599. Ornithologiae: hoc est de avibus historia. Italy: University of Bologna.
- AOAC. 2000. Official methods of analysis. 17th ed. Arlington, VA, USA: Association of Official Analytical Chemists.
- Barbut S, Sosnicki AA, Lonergan SM, Knapp T, Ciobanu DC, Gatcliffe LJ, Huff-Lonergan E, Wilson EW. 2008. Progress in reducing the pale, soft and exudative (PSE) problem in pork and poultry meat. Meat Sci. 79:46–63.
- Cassandro M, De Marchi M, Penasa M, Rizzi C. 2015. Carcass characteristics and meat quality traits of the Padovana chicken breed, a commercial line, and their cross. Ital J Anim Sci. 14:304–309.
- Collins WM, Hubbard W. 1958. Influence of plumage color on hatching ratio and growth rate in chickens. Poult Sci. 37:69.
- Commission International de l’Eclairage. 1978. Recommendations on uniform color spaces, color differences, equations. Psychometric color terms. Commission International de l’Eclairage (CIE), Colorimetry, Paris, France.
- Conway CJ, Martin TE. 2000. Effects of ambient temperature on avian incubation behavior. Behav Ecol. 11:178–188.
- Folch J, Lees M, Sloane-Stanley GH. 1957. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 226:497–507.
- Jaap RG, Grimes JF. 1956. Growth rate and plumage color in chickens. Poult Sci. 35:1264–1269.
- Lepetit J. 2007. A theoretical approach of the relationships between collagen content, collagen cross-links and meat tenderness . Meat Sci. 76:147–159.
- Mills MG, Patterson LB. 2009. Not just black and white: pigment pattern development and evolution in vertebrates. Semin. Cell Dev. Biol. 20:72–81.
- Nesheim MC, Austic RE, Card LE. 1979. Poultry production. 12th ed. London, UK: Bailliere Tindall. Chapter 3, Poultry breeding; p. 58–91.
- Petracci M, Mudalal S, Soglia F, Cavani C. 2015. Meat quality in fast-growing broiler chickens. Worlds Poult. Sci. J. 71:363–374.
- Rizzi C. 2018. Plumage colour in Padovana chicken breed: growth performance and carcass quality. Ital J Anim Sci. 17:797–803.
- Rizzi C, Contiero B, Cassandro M. 2013. Growth patterns of Italian local chicken populations. Poult Sci. 92:2226–2235.
- Rizzi C, Marangon A, Chiericato GM. 2007. Effect of genotype on slaughtering performance and meat physical and sensory characteristics of organic laying hens. Poult Sci. 86:128–135.
- Tasoniero G, Cullere M, Baldan G, Dalle Zotte A. 2018. Productive performances and carcase quality of male and female Italian Padovana and Polverara slow-growing chicken breeds. Ital J Anim Sci. 17:530–539.
- Veneto Agricoltura. 2004. Settore ricerca e Sperimentazione Agraria e Ittica. Progetto CO.VA. Interventi per la Conservazione e la Valorizzazione di razze avicole locali venete. Legnaro - Padova (I): Veneto Agricoltura.
- Verdiglione R, Cassandro M. 2013. Characterization of muscle fiber type in the pectoralis major muscle of slow-growing local and commercial chicken strains. Poult Sci. 92:2433–2437.
- Verdiglione R, Rizzi C. 2018. A morphometrical study on the skull of Padovana chicken. Ital J Anim Sci. 17:785–796.
- WPSA. 1984. World Poultry Science Association, Working group no. 5. Method of dissection of broiler carcases and description of parts. (ed. J. Fris Jensen), Frederiksberg Copenaghen, Denmark, Papworth Everard, Cambridge, UK.
- Yilmaz A, Tepeli C, Garip M, Caglayan T. 2011. The effects of incubation temperature on the sex of Japanese quail chicks. Poult Sci. 90:2402–2406.