
Abstract
Steer meat production is insufficient to meet market demand in northern Spain. This study aimed to compare steer production from two local beef breeds, Asturian Valley (AV) and Asturian Mountain (AM), and two management strategies, differing in yearling steers grazing summer mountain pastures (M treatment) or lowland grasslands (L treatment). A total of 83 calves born in winter-spring were studied during four whole life-cycles. During their first year of life, calves were managed with their dams on summer pastures and weaned when returned to lowlands. Once castrated, half of the yearlings of each breed were randomly assigned to treatment M or L. After summer grazing, all steers were managed at lowlands until they were slaughtered at an age of approximately 33 months after a finishing period of 3–4 months with maize silage and concentrate diet. In general, bodyweight gains were greater in AV than in AM breed, resulting in greater slaughter (714 versus 616 kg) and carcase weights. During the second summer season, AM steers gained more bodyweight than AV in summer pastures, resulting in a breed × management interaction. In the next periods (autumn grazing, winter and spring grazing), M steers showed a compensatory growth counterbalancing their previous lower performance, whereas no differences between treatments or breeds were found in the finishing period. As a result, no differences between managements were observed in final bodyweight at slaughter or carcase weight. Summer pastures offer opportunities to manage yearling steers without adversely affecting saleable product yields.
Steer meat production could be enhanced utilising local animal and pasture resources.
Although the larger local breed was more productive, the smaller breed was better suited to mountain pastures.
Grazing steers on summer mountain pastures as yearlings resulted in similar yields at slaughter compared with those using lowland grasslands.
Highlights
Introduction
Steer meat is a highly valued and sought after product by consumers and the restaurant industry in most regions of Spain (Vieira et al. Citation2007). However, beef production is almost completely focussed on veal and, to a much lesser extent, on bull beef production, so the market is deficient in steer meat. This shortage prevails throughout the humid northern region of Spain, including Asturias, with abundant pasture resources and where beef production is one of the bases of the regional economy. Furthermore, the extensive genetic variation in cattle of this region, with 14 anciently adapted autochthonous cattle breeds (11 of them in danger of extinction; MARM Citation2010), provides opportunities for extensive grazing systems and sustainable steer production.
Two native cattle breeds are present in Asturias: Asturian Valley (AV) and Asturian Mountain (AM) cattle. Both breeds are adapted to extensive systems and their management is mostly based on the use of semi-natural pasturelands as grazing resources. Asturian Valley cattle are widespread throughout the region. This breed is specialised in beef production, with animals having a good conformation, large capacity for food processing and a great growth rate (Cañón et al. Citation1994; Piedrafita et al. Citation2003; Albertí et al. Citation2008). The AV breed includes a double-muscled phenotype, a genetically determined muscular hypertrophy (Grobet et al. Citation1998) that produces heavier and leaner carcases from concentrate-finished yearling bulls than other genotypes (Martínez et al. Citation2010). On the other hand, the AM breed is declared endangered and is mostly restricted to the mountain areas of eastern Asturias (Picos de Europa and surroundings). The animals from this breed present a smaller body size and are more rustic and better adapted to harsh conditions than AV animals (Cañón et al. Citation1994; Osoro et al. Citation1999).
In Asturias, as in many other mountain areas, the traditional beef cattle management is the valley-mountain system. In this system, suckler cows and their calves are moved from lowlands to summer mountain pastures from May–June to September–October. The rest of the year animals graze in lowland grasslands, while the housing period is usually restricted to winter, coinciding with the calving season. Currently, both lowland and mountain pastures are increasingly underutilised, endangering their productivity and biodiversity. The integration of lowland and mountain pastures promotes an efficient use of the available grazing resources, both economically and environmentally (Hadjigeorgiou et al. Citation2005). Although cow-calf operations under this management have been studied with both AV and AM breeds (Osoro et al. Citation1998, Citation1999), there is very little information on the use of mountain pastures by yearling steers.
The aim of this study was to evaluate the potential of mountain pastures for steer production comparing the two Asturian local breeds, based on the integrated use of lowland grasslands with summer mountain pastures. We compare two production systems: one consisting of yearling steers utilising summer mountain pastures in their second year of life, and another consisting of yearling steers grazing on lowland improved pastures. In the third year, all steers grazed on lowland pastures, and were offered maize silage and concentrate during a finishing period of 3–4 months to be slaughtered at an age of approximately 33 months.
Materials and methods
Study sites and available pastures
The study was carried out from 2010 to 2015 at four different locations in Asturias (northern Spain): (1) Villanueva (municipality of Villaviciosa, 43° 27′ N, 5° 25′ W, 50–80 m a.s.l.), in the coastal area, with old-sown grasslands that predominantly consisted of perennial ryegrass (Lolium perenne) and white clover (Trifolium repens); (2) Priesca (Villaviciosa, 43° 28′ N, 5° 21′ W, 85–140 m a.s.l.), in the coastal area, with semi-abandoned grasslands with incipient invasion of shrubs and bracken (Pteridium aquilinum); (3) La Mata (municipality of Grado, 43° 22′ N, 6° 4′ W, 65 m a.s.l.), in an inner valley, with improved perennial ryegrass-white clover grasslands that are occasionally reseeded; (4) Puertos de Agüeria (nature park of Las Ubiñas-La Mesa, municipality of Quirós, 43° 2′ N, 5° 55′ W, 1600–1800 m a.s.l.), with mountain pastures composed of semi-natural grasslands dominated by Festuca rubra and Agrostis capillaris (70% of the area), and Calluna vulgaris heathlands including broom formations of Genista florida (30% of the area).
The lowland old-sown grasslands at Villanueva were annually fertilised with 35, 100 and 65 kg/ha of nitrogen (N), phosphate (P2O5) and potash (K2O), respectively, sometimes substituted by manure, while the respective amounts at La Mata improved grasslands were 120, 120 and 80 kg/ha. In the semi-abandoned grasslands at Priesca, apart from fertilisation with NPK (same application rates as in Villanueva) and manure, additional improvements during the first two years of the study included clearing of shrub- and bracken-invaded areas and scattered sowing of perennial ryegrass and white clover. No management except grazing was performed in the mountain pastures of Agüeria.
Experimental design and animal management
A factorial design of 2 breeds × 2 management strategies was established over four whole life-cycles of the experimental animals. The subjects of the study were 83 male calves from the two Asturian breeds, 39 AV and 44 AM, born during winter and early spring (December–April) in four consecutive years: 2010 (8 AV, 10 AM), 2011 (11 AV, 14 AM), 2012 (12 AV, 12 AM) and 2013 (8 AV, 8 AM). None of the AV animals were phenotypically double-muscled. During spring, calves were reared with their mothers at the coastal old-sown grasslands until mid-June, when they were moved to the summer pastures at Puertos de Agüeria. Cows and calves spent the summer grazing mountain pastures until the end of season in late September or early October. Once returned to lowlands, calves were weaned and taken to Priesca semi-abandoned pastures for autumn grazing until December–January, when they were housed for wintering and fed with beet pulp, concentrate and grass hay (2.5, 1.5 and 1.5 kg/day per animal, respectively). In January–February, weanlings, approximately 1 year of age, were castrated by surgical removal of the testicles. Prior to castration, weanlings were sedated with 0.5 mL/100 kg bodyweight (BW) of 2% xylazine and locally anaesthetised using lidocaine (60 mL/animal).
In the second spring, from March to June, the yearlings grazed on the semi-abandoned grasslands. In mid-June, half of the steers (balanced by breed and BW) were randomly assigned to one of two groups: one group was taken to Agüeria mountain pastures (M treatment), while the other was moved to La Mata to utilise lowland improved pastures (L treatment). In October, all steers were regrouped and managed together at La Mata, where they stayed until their slaughter. The period includes autumn grazing, winter housing, spring–summer grazing in the third year of life and the finishing period. During winter steers were fed with rationed concentrate (3 kg/day per steer, increasing to 4 kg/day in the last month) and barley straw ad libitum. During the finishing period, which lasted 3–4 months (99–114 days) from August–September to November–December, steers were fed with concentrate feed (5 kg/day) and maize silage ad libitum, including barley straw in the initial days. The commercial concentrate was composed of 40% maize, 27% barley, 15% soybean meal, 8% wheat bran, 4% soybean husks, 3% soy oil and 3% minerals (calcium carbonate, sodium bicarbonate, magnesium oxide and sodium chloride). Its nutritional content on a dry matter (DM) basis was 14% crude protein (CP), 5% crude fibre, 6% crude fat and 6% crude ash, in addition to added vitamins (A, D3, E) and trace elements (Zn, Mn, Cu, Co, Se, I). The nutritional content of the maize silage was (on a DM basis) 8.1% CP, 47.4% neutral detergent fibre (NDF), 29.8% starch and 4.1% ash.
Animals were drenched against gastrointestinal nematodes with albendazole (Albendex®, S.P. Veterinaria, Riudoms, Tarragona, Spain) and closantel (Endoex®, S.P. Veterinaria) before spring grazing, with deltametrine (Deltavex pour-on®, S.P. Veterinaria) before summer grazing, and with ivermectin (Virbamec®, Virbac, Esplugues de LLobregat, Barcelona, Spain) during autumn. Before summer, steers were vaccinated against Clostridium spp. (Miloxan®, Merial, Lyon, France). When steers reached target BW of around 700 kg and 600 kg for AV and AM breeds, respectively, they were slaughtered in a commercial abattoir at an age of approximately 33 months. All procedures were performed in accordance with the EU Directive 2010/63/EU on the protection of animals used for scientific purposes.
Measurements
Botanical composition of the grazed pastures was measured in spring (March–April) at the lowland old-sown (2011–2014), semi-abandoned (2012–2014) and improved grasslands (2010–2014) by recording 1000–1200 vertical hits at random at each site with a point quadrat (Grant Citation1981). In the mountain pastures, botanical composition was assessed at the end of June of 2011, 2013 and 2014 with the same method, recording 1000 and 500 contacts on grasslands and heathlands, respectively.
Herbage samples from lowland sites were collected during spring grazing seasons (May) to analyse their nutritive quality. Semi-abandoned grasslands utilised after weaning were also sampled in October, while at mountain pastures, herbage and heather samples were collected in July and September. Samples were stored at −20 °C, freeze-dried and milled to 1 mm before chemical analyses. Organic matter (OM) and CP contents were determined following the procedures of AOAC (Citation2006). Acid detergent fibre (ADF), acid detergent lignin (ADL) and NDF were analysed by the methods of Van Soest et al. (Citation1991).
Green herbage availability was assessed fortnightly during the grazing seasons at lowland pastures by measuring the sward height at 250–500 random points at each site with a swardstick (Barthram Citation1986). In the mountain grasslands sward height was measured at monthly intervals at 300–500 random points.
Animals were weighed at the start and at the end of each grazing or housing season. Daily BW gains were calculated for each season. At slaughter, carcases were weighed and carcase yield was calculated as dressing percentage (100 × hot carcase weight/final BW).
Statistical analysis
Data on chemical composition of spring herbage from lowland sites were analysed by one-way analysis of variance (ANOVA) to examine differences among sites. For semi-abandoned grasslands, differences between spring and autumn were also examined by the same method. Data from mountain pastures were subjected to factorial ANOVA to test the effects of plant component (herbage and heather), season (July and September) and their interaction. Animal performance data were analysed by factorial ANOVA with individual animals as experimental units (N = 83). Individuals were considered as independent replicates for the productive traits measured (BW changes) and the significance of effects was tested against random variation between animals (Iason and Elston Citation2002). This statistical approach is widely adopted in grazing system studies (e.g. Yarrow et al. Citation1996; Fraser et al. Citation2009), even when measurements may be correlated among individuals (e.g. behavioural responses due to social interactions), because of the problem of replicating herds of sufficient size in these complex systems (Connolly Citation2018). For BW changes until treatments were imposed, the model included the effects of breed (AV versus AM), birth year (2010, 2011, 2012 and 2013) and their interaction, leaving 75 degrees of freedom (d.f.) for the error term. For BW changes during the following phases of the life cycle, the effects of breed, treatment (M versus L), birth year and the full interactions were examined (67 d.f. for the error term). Data on slaughter BW, carcase weight and yield were examined with the preceding model including the age of the animal (in days) as covariate (66 d.f. for the error term). Tukey’s test was used for multiple comparisons. All analyses were performed with Statistica 8.0 (StatSoft Inc., Tulsa, OK).
Results
Pastures
In the coastal old-sown grasslands, sown species accounted for 36% cover (26–28% ryegrass and 5–10% clover; Table ). Dominant grasses were Agrostis spp. and Holcus lanatus, with Poa spp., Festuca spp. and Dactylis glomerata also being relatively abundant. Forbs accounted for 6–8% cover, being Plantago lanceolata and Ranunculus spp. the most abundant. The nearby semi-abandoned pastures were dominated by grasses (56–63%), especially Agrostis spp., with a lower presence of D. glomerata and H. lanatus. Non-legume forbs and weeds were more abundant than in other lowland grasslands (16% cover), with Centaurea nigra and P. lanceolata as the most abundant, together with Ranunculus spp. and Rumex spp., among others. There was an incipient colonisation by woody species like Rubus spp. and Prunus spinosa. The presence of bracken was reduced from 7.4% in 2012 to 1.3% in 2014 due to clearing, while dead matter accounted for a mean of 8%. The improved grasslands were dominated by perennial ryegrass (51–56%), with white clover accounting for 3–10%. Pooled cover of unsown grasses (mostly Agrostis spp. and Poa spp.) varied between 27% and 34%, while the mean cover of forbs was 8%, with Capsella bursa-pastoris as the most abundant. Mountain grasslands were highly diverse, with F. rubra as the most abundant species (36%), together with other grasses such as A. capillaris (10%) and Nardus stricta (5%). Among the legumes (8–9%), T. repens and Lotus corniculatus were the most important. Among the many other species (forbs and monocots), Merendera montana, Plantago media, Carex caryophyllea, Plantago alpina and Hieracium pilosella were relatively abundant. Heathlands were dominated by C. vulgaris (40–48%), with bilberry (Vaccinium myrtillus) as the most abundant companion species (8–11%). Potentilla erecta and Succisa pratensis had the highest presence among the herbaceous species.
Table 1. Botanical composition (mean cover percentages ± SE) of the pastures grazed by steers at each study site.
The nutritive quality of spring herbage did not differ among lowland sites, averaging 899 ± 2.5 g OM/kg DM, 189 ± 4.0 g CP/kg DM, 469 ± 5.4 g NDF/kg DM, 256 ± 3.9 g ADF/kg DM and 26 ± 1.1 g ADL/kg DM (n = 36; Table ). In the semi-abandoned grasslands CP content tended to decrease from spring to autumn (p = .089), whereas ADF and ADL contents increased (p < .01 and p < .001, respectively). In mountain pastures, grassland herbage presented greater CP (178 versus 90 g/kg DM; SEM = 4.2; p < .001) and NDF contents (526 versus 408 g/kg DM; SEM = 20.7; p < .01), and lower OM (930 versus 968 g/kg DM; SEM = 2.3; p < .001), ADF (246 versus 319 g/kg DM; SEM = 15.4; p < .01) and ADL contents (34 versus 190 g/kg DM; SEM = 8.9; p < .001) than heather. Herbage CP content tended to increase from July to September (p = .07), whereas heather CP content decreased (p < .05; Table ).
Table 2. Chemical composition (means ± SE in g/kg DM) of the herbage from the different pastures, including heather green shoots from the mountain heathlands.
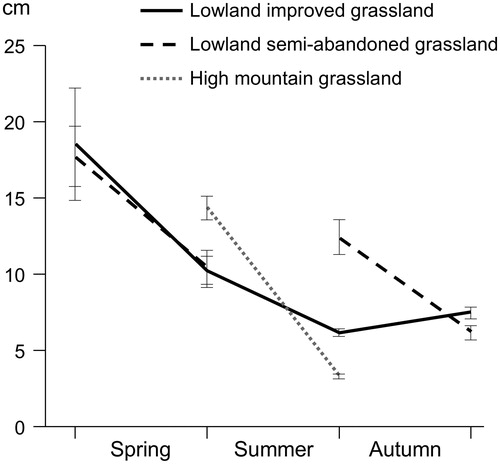
In general, green herbage availability decreased during each grazing season at all sites. Mean sward height during spring ranged between 12.6 cm and 14.4 cm in lowland grasslands, decreasing from 17.8–18.6 cm at the beginning to 10.2–10.5 cm at the end (Figure ). During summer, mean sward height of improved grasslands was 7.3 cm, decreasing from 10.2 cm in June to 6.2 cm in September–October. In mountain grasslands, mean sward height during summer was a little lower (6.1 cm), although a greater reduction rate was observed, from a mean of 14.4 cm to 3.3 cm at the end of the summer. During autumn, mean sward height of improved grasslands was 7.9 cm, fluctuating between 6.0 and 9.5 cm, while it was 9.7 cm in semi-abandoned grasslands, decreasing from 12.4 to 6.2 cm (Figure ).
Figure 1. Sward height of the pastures grazed by steers (means ± SE of 4–5 years at each site at the start and end of each grazing season).
Steer performance
Liveweight gains across the life cycle according to breed and treatment are shown in Figure . During their first year of life, BW gains were greater in AV than in AM calves, except in winter (Table ). In spring, suckling AV calves gained 21% more BW than AM calves (p < .01). In summer, when calves were in mountain pastures with their dams, daily BW gains were 15% higher in AV than in AM (p < .05). In autumn grazing, when animals returned to lowland pastures and were weaned, the gains were 37% higher for AV calves compared to AM (p < .01). In winter, calves fed in the stables achieved mean BW gains of 642 ± 28.3 g/day, with no differences between breeds, although being greater (p < .001) in those born in 2011 compared to the rest. During the next spring grazing season at semi-abandoned grasslands, AM steers tended to gain more BW than AV steers (p = .086; Table ).
Figure 2. Changes in bodyweight (mean ± SE) during the life cycle of steers from Asturian Valley (AV) and Asturian Mountain (AM) breeds managed at lowland (L treatment) or mountain (M treatment) pastures during the summer of the second year.
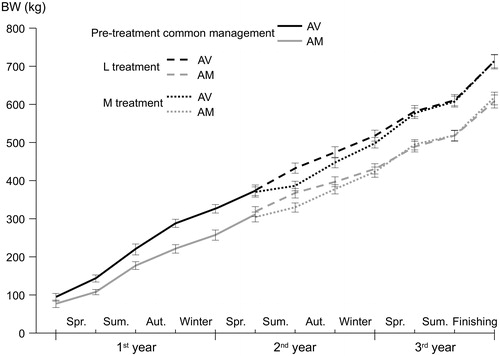
Table 3. Daily bodyweight gains (g/day) of male calves from two breeds (AV: Asturian Valley; AM: Asturian Mountain) during the early stages of their life: spring grazing at lowlands and summer grazing in the mountain as sucklers with their mothers, autumn grazing at lowlands after weaning, indoor wintering, and spring grazing after being castrated (LSmeans of 4 years, N = 83).
In the summer of the second year, once treatments were imposed, L steers had higher BW gains than M steers (485 versus 193 g/day; SEM = 26.8; p < .001), although the extent of such differences varied among years (p < .01 treatment × year). There was an interaction (p < .05) between breed and management, as in L treatment AV steers gained more BW than AM steers (512 versus 459 g/day), whereas in M treatment AM steers showed a better performance than AV ones (252 versus 133 g/day; Table ). The differences between managements increased from the first half of the summer grazing season (until early August; 672 versus 424 g/day in L and M treatments, respectively; SEM = 26.0; p < .001) to the second half, when a general poor performance was observed in mountain pastures (402 versus 54 g/day; SEM = 42.0; p < .001).
Table 4. Daily body weight gains (g/day) of steers from two breeds (AV: Asturian Valley; AM: Asturian Mountain) managed during summer grazing (second year of life) at lowlands (L) or mountain pastures (M), and thereafter managed at lowlands until their slaughter (LSmeans of 4 years, N = 83).
In the following autumn, steers grazing lowland improved grasslands from M treatment showed greater daily BW gains than those from L treatment (642 versus 350 g/day; SEM = 30.3; p < .001). These differences increased year after year from non-significant 70–140 g/day in those born in 2010 and 2011 to 371–590 g/day (p < .001) in those born in 2012 and 2013 (p < .001 treatment × year). In this season, AV steers presented greater gains than AM ones (556 versus 436 g/day; SEM = 30.6; p < .01). During the second winter, AV steers tended to gain more BW than AM (569 versus 503 g/day; SEM = 26.2; p = .074), with M steers having greater BW gains than L steers (597 versus 475 g/day; SEM = 25.9; p < .01). In the following spring grazing season (third year of life), differences in performance followed a similar pattern to those found in the preceding periods, with AV steers gaining more BW than AM (905 versus 787 g/day; SEM = 28.1; p < .01), and M steers still showing greater gains than L steers (931 versus 762 g/day; SEM = 27.9; p < .001). In the summer grazing preceding the finishing period, there were no differences between breeds or managements, with steers achieving mean BW gains of 591 ± 38.0 g/day. During the finishing period, steers gained an average of 922 ± 28.1 g/day, with no significant differences between breeds, managements or years (Table ).
Yields at slaughter
Steers were slaughtered at an average age of 1002 ± 6.7 days, although there were differences between years (p < .05), ranging from a minimum of 972 days in the steers born in 2012 to a maximum of 1024 days in those born in 2011. Steers from M treatment tended to be older than those from L treatment (1013 versus 990 days; SEM = 8.9; p = .076), with no differences between breeds (Table ). Age, introduced as a covariate, influenced (p < .01) slaughter and carcase weight with positive coefficients (i.e. greater weights the older the age), but not carcase yield. After subtracting age effect, BW at slaughter was greater in AV than in AM steers (714 versus 616 kg; SEM = 10.0; p < .001). In addition, AV steers were found to have higher carcase weights than AM steers (387 versus 317 kg; SEM = 5.6; p < .001), whereas no significant management effects were found in either slaughter or carcase weights (Table ). The dressing percentage was greater in AV than in AM steers (54.1% versus 51.5%; SEM = 0.33; p < .001). Although in general L steers tended to have carcase yields 1% higher than M steers (53.2% versus 52.3%; SEM = 0.34; p = .065), this effect was not constant across years, resulting in an interaction between management and year (p < .05).
Table 5. Age and yields at slaughter of steers from two breeds (AV: Asturian Valley; AM: Asturian Mountain) managed during summer grazing of their second year of life at lowlands (L) or mountain pastures (M), and thereafter managed at lowlands until their slaughter (covariate-adjusted –except for age– LSmeans of 4 years, N = 83).
Discussion
Available pastures
The pastures utilised in this study represent the typical grasslands used for livestock grazing in Green Spain, concerning both lowlands and high mountains. In spite of the observed differences in botanical composition, no differences in herbage nutritive quality were found between the lowland grasslands, having CP and fibre contents similar to those found in permanent grasslands in other Spanish Atlantic provinces (e.g. Salcedo and Sarmiento Citation1994; Brea et al. Citation1998). In addition, mountain grasslands presented a nutritive quality roughly similar to that found in other mesic grasslands from Cantabrian Mountains (e.g. Mandaluniz et al. 2009; Bedia and Busqué Citation2013), although the herbage in the current study had higher CP content and lower fibre contents compared to those studies (Mandaluniz et al. 2009; Bedia and Busqué Citation2013). In general, both lowland and mountain grasslands had a good nutritive quality to meet beef cattle protein and energy demands, so they could maintain acceptable levels of animal production. On the contrary, heather plants are known to have a much poorer nutritive quality than improved or semi-natural grasslands (Celaya et al. Citation2007; Mandaluniz et al. 2009), limiting animal performance.
Apart from its nutritive quality, sufficient herbage allowance is necessary to achieve adequate levels of nutrient intake by grazing cattle (Baker et al. Citation1981). Different studies have shown that cattle obtain maximum intake rates with sward heights around 8–10 cm in spring ryegrass-clover pastures, enhancing animal performance (Wright et al. Citation1990; Realini et al. Citation1999). In the current study, mean sward heights during spring were maintained above 10 cm at both improved and semi-abandoned lowland grasslands, and thus it is to be assumed that steers could obtain adequate herbage intakes to grow well in this season. However, at the end of summer grazing on improved grasslands, herbage allowance could be limiting steers’ intake, so concentrate supply at pasture is advisable to enhance BW gains, especially before the finishing period to facilitate diet change. On the other hand, mountain grasslands provided enough herbage amounts during the first half of the summer grazing season, but sward height was markedly reduced at the end of season. In the same experimental farm, similar low sward heights were measured at this time, negatively affecting cow-calf performance (Osoro et al. Citation1998, Citation1999). During autumn, when taller sward heights are recommended for bulls and steers (Morris et al. Citation1993), herbage shortage could limit animal performance at specific times, especially at the end of season when grazing on semi-abandoned grasslands.
Breed and management effects on steer performance
Throughout most of the life cycle, AV steers had greater BW gains than AM ones. This was an expected result as AV breed has a larger mature body size than AM (Cañón et al. Citation1994). Although both breeds have a common origin from the Cantabric trunk, AV animals are late maturing and show a higher growth potential, more capacity for protein deposition and less tendency to deposit fat compared to AM, which are intermediate maturing and accumulate more fat amounts (Piedrafita et al. Citation2003; Albertí et al. Citation2008). Our results show faster growth rates of AV than AM calves during the first year grazing seasons, i.e. both during pre-weaning (suckling their dams during spring-summer) and early post-weaning (autumn). However, previous studies at the same summer pastures (Osoro et al. Citation1999) showed similar calf BW gains between the same breeds (810–820 g/day), which could be partly attributed to the higher milk production of AM cows at the end of the grazing season (1.76 versus 1.32 kg/day) counterbalancing the higher growth potential of AV calves. The lower BW gains observed in the present study, especially in AM calves (618 g/day), could be due to a more extended grazing season, thus, relying on lower sward heights during the last weeks and negatively affecting herbage and milk intake by calves, in addition to worse weather conditions or animal genetics compared with the study of Osoro et al. (Citation1999).
The marginal difference between breeds observed in the first wintering period was possibly a consequence of the limited amounts of feed offered, intended to maintain acceptable BW gains with low feeding costs. Under these conditions calves could not fully express their growth potential. During the following spring, unlike the first year, AM steers obtained greater BW gains than AV steers, which showed an unexpectedly poor performance grazing on semi-abandoned grasslands. Although herbage nutritive quality and availability were similar to other lowland grasslands, other factors like the more rugged terrain and higher presence of unsown, less palatable plant species could increase animals’ ME requirements whilst reducing voluntary DM intake (Wright et al. Citation1994). These handicaps were better overcome by AM steers, as shown by their higher BW gains compared to AV steers.
When treatments were imposed during the second summer grazing season, important differences in steers’ BW gains were found between M and L managements. Different reasons explain the general poor performance observed in high mountain pastures. Environmental factors, such as inclement weather and rough terrain negatively affect animal performance (Casasús et al. Citation2002). Herbage nutritive quality was not much poorer compared to lowland grasslands, although moderately lower CP and higher NDF contents would reduce nutrient intake. More than this, the much shortened sward height in mountain grasslands in the second half of summer undoubtedly implied a reduction in herbage intake. As pointed above, herbage intake and performance are positively correlated with sward height (Wright et al. Citation1990; Osoro et al. Citation1998). Although steers could increase their grazing time on heathlands as preferred herbage was depleted (as observed in previous studies with cattle grazing grassland–heathland mosaics; Gordon Citation1989; Ferreira et al. Citation2013), those shrub communities do not provide enough nutrients for the maintenance of grazing cattle (Hodgson et al. Citation1991; Fraser et al. Citation2009; Celaya et al. Citation2011). In addition, a breed × management interaction was observed, as the lowering of performance under M treatment was much more marked in AV than in AM steers. Genotype (regarding breed body size)×environment (physical or nutritional) interactions have been previously observed with cattle (Ferrell et al. Citation1978; Fitzhugh Citation1978). It is generally assumed that smaller breeds can thrive better under harsher conditions of limited energy supply than more energy demanding animals (Morris et al. Citation1993; Wright et al. Citation1994). In summer pastures, AM cows showed less unfavourable BW changes than AV cows when heathlands prevailed over grasslands, leading to very limited availabilities of preferred herbage (Osoro et al. Citation1999). The results from the current study suggest that the smaller AM steers are better suited than the larger AV breed to utilise these mountain pastures. As pointed out by Vercoe and Frisch (Citation1992), BW gain in favourable environments would be a reflection of growth potential, but a reflection of resistance to environmental stresses in harsh ones.
In the following periods (autumn grazing, winter and spring grazing), regardless of the breed, steers from M treatment showed compensatory growth, achieving greater BW gains than those from L treatment. Numerous studies have observed such compensatory growth in beef cattle after a period of restricted feeding (e.g. Wright et al. Citation1989; Sainz et al. Citation1995; Yarrow et al. Citation1996). Different mechanisms have been proposed for it, such as increased intake, lowered maintenance requirements, increased net efficiency of tissue growth or altered metabolism (Hornick et al. Citation2000), and several factors can affect its magnitude, such as severity and duration of restriction, age or growth stage (Sainz et al. Citation1995). In our study, the effect was observed for a long time (around 250 days), although not significantly in the first two study years during the autumn grazing. During the same periods, AV steers had higher BW gains than AM ones but the differences were no longer apparent during the last summer grazing, possibly due to the limited sward height, in addition to the large between year differences observed.
Marked differences in animal performance among study years were noted in most phases of the life cycle of the steers. These differences would be mainly related to climatic conditions affecting animal welfare directly or indirectly through pasture conditions. As the differences among study years were not consistent throughout the different phases of the life cycle, animal factors such as genetics or age would be of minor importance. In the case of semi-abandoned grasslands during post-weaning autumn grazing, BW gains did not show an increasing pattern across study years, so the pasture improvements made were not enough to enhance animal performance.
During the finishing period, no breed or management effect on steer performance was observed. Under more intensive growth conditions with yearlings or young bulls, higher BW gains were found in AV (1300–1400 g/day) compared with AM (1000–1100 g/day; Piedrafita et al. Citation2003; Albertí et al. Citation2008; Martínez et al. Citation2010). The lower BW gains obtained in the current study, apart from the known reduced growth rates in steers compared to intact bulls (Field Citation1971; Bretschneider Citation2005; Álvarez-Rodríguez et al. Citation2017), would be partly related to lower concentrate feeding rates, in addition to the BW being closer to the maximum adult BW attainable. In support, even lower gains (500 g/day) were obtained in AV oxen fed 10 kg/day of concentrate combined with forage (Vieira et al. Citation2007). In the case of AV breed, it should also be noted that both double-muscled and normal animals were studied in some works, with the former ones showing higher gains (Martínez et al. Citation2010).
Yields at slaughter
Final BW at slaughter was greater in AV than in AM steers according to the targeted objectives. This was achieved thanks to the greater growth potential of AV breed, as both breeds were managed under the same pasture and feeding conditions throughout the life cycle. Notably, there were not differences between managements, so the observed compensatory growth was enough to compensate for the differences established during the second summer, although at the expense of a somewhat longer finishing period and lower carcase yield. As well, carcases from AV steers were heavier than those from AM, regardless of previous management. Albertí et al. (Citation2008) noted that, in general, carcase weight and dressing percentage are more influenced by breed type than by environment. In other studies comparing these breeds, heavier carcases and higher dressing percentages were obtained for the specialised beef AV compared with the more rustic AM (Vallejo et al. Citation1992a, Citation1992b; Piedrafita et al. Citation2003; Martínez et al. Citation2010). Compared with these studies with intact animals, in general lower dressing percentages were observed in the current work, excepting that of Vallejo et al. (Citation1992a) with AM. Although differences between steers and bulls in dressing percentage are not consistent across different cattle breeds (Field Citation1971), greater values have been found in bulls than in steers from another Spanish local breed (Serrana de Teruel; Ripoll et al. Citation2016). Nevertheless, the dressing percentages in AV steers from our study continue to be lower than those obtained with AV oxen slaughtered at 42 months old (Vieira et al. Citation2007), which is probably related to the feeds offered during the finishing phase.
This study did not include quality parameters in carcase and meat. Although productive traits were superior in AV than in AM steers, the quality of saleable product should also be taken into account when evaluating market prices and final profit. In that sense, young AM bulls have previously been found to show higher fatness and better sensory attributes than AV ones (Aldai et al. Citation2007; Martínez et al. Citation2010; Sierra et al. Citation2010). Therefore, a greater added value could compensate to a certain extent the lower carcase yields obtained with AM steers but further studies are required to investigate this.
Conclusions
Overall, AV steers showed their higher growth and productive potential compared to AM, achieving greater BW gains throughout most of their life cycle and reaching significantly higher yields at slaughter. Nevertheless, in situations of limited forage availability, AM steers attained performance levels similar to or higher than AV in summer pastures as yearlings. Their hardiness and smaller body size allow them to thrive better under harsh environmental and nutritional conditions like those in high mountains. Although managing yearling steers in summer mountain pastures considerably lowered their BW gains compared to those achieved in lowland improved grasslands, the compensatory growth shown in the next seasons was enough to equalise final BW and carcase weight. Valley-mountain system could constitute a cost-effective alternative for steer production when properly managed, not prolonging summer grazing season too much and taking advantage of compensatory growth. Apart from the final yield and product quality, the environment and available pasture resources should also be considered to choose the appropriate breed for steer production. In conclusion, the findings show that summer pastures can be used to grow yearling steers of these well-adapted local breeds to achieve saleable product yields similar to those obtained at lowland improved pastures.
Ethical approval
The study was conducted upon the approval of the Bioethics Committee of SERIDA.
Acknowledgements
The authors thank the staff of Animal Production Systems Area of SERIDA for their work in animal management.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Albertí P, Panea B, Sañudo C, Olleta JL, Ripoll G, Ertbjerg P, Christensen M, Gigli S, Failla S, Concetti S, et al. 2008. Live weight, body size and carcass characteristics of young bulls of fifteen European breeds. Livest Sci. 114:19–30.
- Aldai N, Nájera AI, Martínez A, Celaya R, Osoro K. 2007. Correlation between carcass conformation and fat cover degree, and muscle fatty acid profile of yearling bulls depending on breed and mh-genotype. Livest Sci. 107:199–212.
- Álvarez-Rodríguez J, Albertí P, Ripoll G, Blasco I, Sanz A. 2017. Effect of castration at 10 months of age on growth physiology and behavior of male feral beef cattle. Anim Sci J. 88:991–998.
- AOAC. 2006. Official methods of analysis. 18th ed. Gaithersburg (MD): AOAC.
- Baker RD, Alvarez F, Le Du Y. 1981. The effect of herbage allowance upon the herbage intake and performance of suckler cows and calves. Grass Forage Sci. 36:189–200.
- Barthram GT. 1986. Experimental techniques: the HFRO swardstick. The Hill Farming Research Organisation biennial report 1984–85. Midlothian (UK): HFRO; p. 29–30.
- Bedia J, Busqué J. 2013. Productivity, grazing utilization, forage quality and primary production controls of species-rich alpine grasslands with Nardus stricta in northern Spain. Grass Forage Sci. 68:297–312.
- Brea T, Monserrat L, Zea J. 1998. Calidad de la hierba aprovechada en pastoreo: efecto del sistema de manejo y de la estación en las condiciones de Galicia [Herbage quality under grazing: management and season effects under Galician conditions]. Pastos. 28:57–70. Spanish.
- Bretschneider G. 2005. Effects of age and method of castration on performance and stress response of beef male cattle: a review. Livest Prod Sci. 97:89–100.
- Cañón J, Gutiérrez JP, Dunner S, Goyache F, Vallejo M. 1994. Herdbook analyses of the Asturiana beef cattle breeds. Genet Sel Evol. 26:65–75.
- Casasús I, Sanz A, Villalba D, Ferrer R, Revilla R. 2002. Factors affecting animal performance during the grazing season in a mountain cattle production system. J Anim Sci. 80:1638–1651.
- Celaya R, Ferreira LMM, García U, Rosa García R, Osoro K. 2011. Diet selection and performance of cattle and horses grazing in heathlands. Animal. 5:1467–1473.
- Celaya R, Oliván M, Ferreira LMM, Martínez A, García U, Osoro K. 2007. Comparison of grazing behaviour, dietary overlap and performance in non-lactating domestic ruminants grazing on marginal heathland areas. Livest Sci. 106:271–281.
- Connolly J, 2018. Perspectives on the use of animal as replicate in grazing experiments. In: Horan B. Hennessy D, O’Donovan M, Kennedy E, McCarthy B, Finn JA, O’Brien B, editors. Sustainable meat and milk production from grasslands. Proceedings of the 27th General Meeting of the European Grassland Federation; Jun 17–21; Cork, Ireland. Wageningen (The Netherlands): Wageningen Academic Publishers. (Grassland Science in Europe, vol. 23). pp. 404–406.
- Ferreira LMM, Celaya R, Benavides R, Jáuregui BM, García U, Santos AS, Rosa García R, Rodrigues MAM, Osoro K. 2013. Foraging behaviour of domestic herbivore species grazing on heathlands associated with improved pasture areas. Livest Sci. 155:373–383.
- Ferrell CL, Kohlmeier RH, Crouse JD, Glimp H. 1978. Influence of dietary energy, protein and biological type of steer upon rate of gain. J Anim Sci. 46:255–270.
- Field RA. 1971. Effect of castration on meat quality and quantity. J Anim Sci. 32:849–858.
- Fitzhugh HA. 1978. Animal size and efficiency, with special reference to the breeding female. Anim Prod. 27:393–401.
- Fraser MD, Theobald VJ, Davies DR, Moorby JM. 2009. Impact of diet selected by cattle and sheep grazing heathland communities on nutrient supply and faecal micro-flora activity. Agric Ecosyst Environ. 129:367–377.
- Gordon IJ. 1989. Vegetation community selection by ungulates on the isle of Rhum. III. Determinants of vegetation community selection. J Appl Ecol. 26:65–79.
- Grant SA. 1981. Sward components. In: Hodgson J, Baker RD, Davies A, Laidlaw AS, Leaver JD, editors. Sward measurement handbook. Hurley (UK): British Grassland Society; p. 71–92.
- Grobet L, Poncelet D, Royo JL, Brouwers B, Pirottin D, Michaux C, Ménissier F, Zanotti M, Dunner S, Georges M. 1998. Molecular definition of an allelic series of mutations disrupting the myostatin function and causing double-muscling in cattle. Mamm Genome. 9:210–213.
- Hadjigeorgiou I, Osoro K, Fragoso de Almeida JP, Molle G. 2005. Southern European grazing lands: Production, environmental and landscape management aspects. Livest Prod Sci. 96:51–59.
- Hodgson J, Forbes TDA, Armstrong RH, Beattie MM, Hunter EA. 1991. Comparative studies of the ingestive behaviour and herbage intake of sheep and cattle grazing indigenous hill plant communities. J Appl Ecol. 28:205–227.
- Hornick JL, Van Eenaeme C, Gérard O, Dufrasne I, Istasse L. 2000. Mechanisms of reduced and compensatory growth. Domest Anim Endocrinol. 19:121–132.
- Iason GR, Elston DA. 2002. Groups, individuals, efficiency and validity of statistical analyses. Appl Anim Behav Sci. 75:261–265.
- Mandaluniz N, Aldezabal A, Oregui LM. 2009. Atlantic mountain grassland-heathlands: structure and feeding value. Span J Agric Res. 7:129–136.
- Ministerio de Medio Ambiente y Medio Rural y Marino [MARM]. 2010. Razas de ganado del Catálogo Oficial de España [Livestock breeds from the Official Catalogue of Spain]. Madrid: MARM [Spanish].
- Martínez A, Aldai N, Celaya R, Osoro K. 2010. Effect of breed body size and the muscular hypertrophy gene in the production and carcass traits of concentrate-finished yearling bulls. J Anim Sci. 88:1229–1239.
- Morris CA, Baker RL, Hickey SM, Johnson DL, Cullen NG, Wilson JA. 1993. Evidence of genotype by environment interaction for reproductive and maternal traits in beef cattle. Anim Prod. 56:69–83.
- Morris ST, Hirschberg SW, Michel A, Parker WJ, McCutcheon SN. 1993. Herbage intake and liveweight gain of bulls and steers continuously stocked at fixed sward heights during autumn and spring. Grass Forage Sci. 48:109–117.
- Osoro K, Fernández Prieto E, Celaya R, Noval G, Alonso L, Castro P. 1999. Respuesta productiva de dos razas de ganado vacuno manejadas en dos cubiertas vegetales de montaña [Productive responses of two cattle breeds managed at two mountain vegetation types]. ITEA. 95A:188–203. Spanish.
- Osoro K, Oliván M, Celaya R. 1998. Comportamiento productivo de las vacas de cría en comunidades vegetales de montaña constituidas por Agrostis-Festuca-Nardus-Calluna y dominadas por herbáceas [Productive performance of suckler cows in mountain plant communities composed of Agrostis-Festuca-Nardus-Calluna and dominated by herbaceous species]. ITEA. 94A:109–124. Spanish.
- Piedrafita J, Quintanilla R, Sañudo C, Olleta J-L, Campo M-M, Panea B, Renand G, Turin F, Jabet S, Osoro K, et al. 2003. Carcass quality of 10 beef cattle breeds of the Southwest of Europe in their typical production systems. Livest Prod Sci. 82:1–13.
- Realini CE, Hodgson J, Morris ST, Purchas RW. 1999. Effect of sward surface height on herbage intake and performance of finishing beef cattle. N Z J Agric Res. 42:155–164.
- Ripoll G, Albertí P, Álvarez-Rodríguez J, Blasco I, Sanz A. 2016. Body size, carcass and meat quality of three commercial beef categories of ‘Serrana de Teruel’ breed. Span J Agric Res. 14:e0604.
- Sainz RD, De la Torre F, Oltjen JW. 1995. Compensatory growth and carcass quality in growth-restricted and refed beef steers. J Anim Sci. 73:2971–2979.
- Salcedo G, Sarmiento M. 1994. Composición nutritiva de las praderas aprovechadas bajo pastoreo rotacional, en la zona costera de Cantabria [Nutritive composition of rotationally managed grasslands in the coastal area of Cantabria]. In: Actas de la XXXIV Reunión Científica de la Sociedad Española para el Estudio de los Pastos (SEEP); May 30–Jun 3. Santander: SEEP. p. 313–317 [Spanish].
- Sierra V, Guerrero L, Fernández-Suárez V, Martínez A, Castro P, Osoro K, Rodríguez-Colunga MJ, Coto-Montes A, Oliván M. 2010. Eating quality of beef from biotypes included in the PGI Ternera Asturiana showing distinct physicochemical characteristics and tenderization pattern. Meat Sci. 86:343–351.
- Vallejo M, Gutiérrez JP, Alonso L, Cañón J, Revuelta JR, Goyache F, Cima M. 1992a. Características de las canales de las razas bovinas asturianas. II Valoración cuantitativa y predicción de la composición tisular de canales en la raza Asturiana de la Montaña [Carcass characteristics of Asturian bovine breeds. II Quantitative evaluation and prediction of tissue composition in carcasses from Asturian Mountain cattle breed]. Arch Zootec. 41:645–656 [Spanish].
- Vallejo M, Gutiérrez JP, Cima M, Cañón J, Alonso L, Revuelta JR, Goyache F. 1992b. Características de las canales de las razas bovinas asturianas. III.- Valoración cuantitativa y predicción de la composición tisular de canales en la raza Asturiana de los Valles [Carcass characteristics of Asturian bovine breeds. III. Quantitative evaluation and prediction of tissue composition in carcasses from Asturian Valley cattle breed]. Arch Zootec. 42:29–40 [Spanish].
- Van Soest PJ, Robertson JB, Lewis BA. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. 74:3583–3597.
- Vercoe JE, Frisch JE. 1992. Genotype (breed) and environment interaction with particular reference to cattle in the tropics. Asian-Australas J Anim Sci. 5:401–409.
- Vieira C, Cerdeño A, Serrano E, Lavín P, Mantecón AR. 2007. Breed and ageing extent on carcass and meat quality of beef from adult steers (oxen). Livest Sci. 107:62–69.
- Wright IA, Jones JR, Maxwell TJ, Russel AJF, Hunter EA. 1994. The effect of genotype × environment interactions on biological efficiency in beef cows. Anim Prod. 58:197–207.
- Wright IA, Russel JF, Hunter EA. 1989. Compensatory growth in cattle grazing different vegetation types. Anim Prod. 48:43–50.
- Wright IA, Whyte TK, Osoro K. 1990. The herbage intake and performance of autumn calving beef cows and their calves when grazing continuously at two sward heights. Anim Prod. 51:85–92.
- Yarrow NH, Penning PD, Johnson RH. 1996. The effect of plane of winter nutrition and sward height on the performance of steers grazing grass/white clover swards. Grass Forage Sci. 51:424–433.