
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
This study was aimed to estimate the relative bioavailability (RBA) of l-methionine (l-Met) to dl-methionine (dl-Met) in broiler chickens from 1 to 21 d of age. A total of 1664 one-day-old broiler chicks were randomly divided into 13 groups with 8 replicates per group. The basal diet contained 60% of the NRC requirement for Met. Birds in treatment groups were fed the basal diet supplemented with graded levels of dl- or l-Met, resulting in the dietary content of Met at 75%, 80%, 85%, 90%, 95% and 100% of the NRC requirement, respectively. Results showed that there were linear increases (p < .05) in average daily gain (ADG), average daily feed intake, feed efficiency (FE), eviscerated weight, breast muscle weight and leg muscle weight in response to the increasing supplementation of either dl- or l-Met, which also linearly reduced (p < .05) plasma uric acid and protein carbonyl contents. Compared with dl-Met, l-Met addition resulted in elevations (p < .05) in ADG and FE, along with leg muscle yield of birds. A lower (p < .05) content of urea nitrogen with a higher (p < .05) content of uric acid in plasma were observed in the birds supplemented with l-Met compared with dl-Met. Slope-ratio assay revealed that the RBA of l-Met to dl-Met for ADG and FE were 141.5% and 189.1%, respectively. And the respective values were calculated as 122.9% and 116.8% when based on eviscerated weight and breast muscle weight as the response criteria. However, the above RBA estimates except for FE did not differ from 100%. Collectively, supplemental l-Met had a higher bioavailability for FE of broilers relative to dl-Met during the first 3 weeks of life.
l-methionine addition has an advantage over dl-methionine for supporting the growth performance of broiler chickens in early age.
The relative bioavailability of l-Met to dl-Met for feed efficiency of broiler chickens during the first 21 days post-hatch was calculated as 189.1% that differs from 100%.
Highlights
Introduction
Methionine (Met) is the first limiting amino acid in the corn-soybean meal based diets of poultry, being critical for protein synthesis and growth of poultry (Baker Citation2006). Beyond the role in protein accretion, Met has numerous biological functions, including methyl donation, antioxidative effects and as precursor of bioactive compounds such as glutathione and taurine (Riedijk et al. Citation2007; Tesseraud et al. Citation2009; Zhang et al. Citation2018). These functional roles of Met are important for the development and health status of animals (Riedijk et al. Citation2007).
dl-Met (an equal racemic mixture of the d- and l-isomers of Met) is commonly added to poultry feeds that are low in natural Met to satisfy the requirement of chickens for this amino acid. However, only l-isomer that had been commercially available by a fermentation approach recently (Huang et al. Citation2017), is the biologically functional form of Met, as it can be used readily in intestinal cells and directly incorporated into protein synthesis (Fang et al. Citation2010). It is indispensable that dietary d-Met is converted to l-Met in a process catalysed by d-Met oxidase that resides in liver, kidney and intestines prior to its utilisation (Fang et al. Citation2010). However, the oxidative activity towards d-Met exists in the intestine only represents about one-half and one-tenth of the hepatic and renal activity, respectively (Brachet and Puigserver Citation1992). Furthermore, there appears to be a specific need for the functional role of Met in maintenance and development of gastrointestinal tract (Stoll et al. Citation1998; Shen et al. Citation2014). It could be thus conjectured that dietary l-Met may deliver benefits more efficiently as compared with d-Met, which was supported by a previous study (Baker and Boebel Citation1980).
There had been several controversies surrounded the relative bioavailability (RBA) of l-Met to dl-Met in pigs. For instance, Htoo and Morales (Citation2016) observed no difference in the bioavailability between dl-Met and l-Met for weaned pigs when average daily gain (ADG) was used as the response criterion. Contradictorily, it was indicated that the RBA of dl-Met to l-Met in nursery pigs were 69.4 and 81.3% for ADG and feed efficiency (FE), respectively (Shen et al. Citation2014). So far, little information was available regarding the RBA of l-Met to dl-Met in chickens. Previous studies revealed that dl-Met was as efficacious as l-Met in improving the growth performance and carcase characteristic in broilers (Dilger and Baker Citation2007; Zhang et al. Citation2018). However, there were differences in their influences on several biochemical pathways inside the body of broilers (Dilger and Baker Citation2007; Zhang et al. Citation2018), moreover, the single dose of Met source employed in the previous studies could not be suitable for evaluating the RBA of l-Met to dl-Met in chickens. Thereby, this study was conducted to determine the RBA of l-Met to dl-Met in broilers using the slope-ratio assay.
Materials and methods
Animals and experimental design
The experimental protocols and procedures were approved by the Animal Care and Use Committee of the Feed Research Institute of the Chinese Academy of Agricultural Sciences. A total of 1664 one-day-old Cobb 500 broiler chicks (832 male and 832 female chicks) were divided into 13 groups with 8 replicates per group in a randomised complete block design based on initial body weight and sex. Each replicate cage encompassed 16 chicks. The basal diets (Table ) were formulated to be deficient in Met but all other essential nutrients were adequate according to the NRC requirements (NRC 1994), which contained Met at 0.3% (60% of the NRC requirement, 1994) from 1 to 21 d of age. Birds in the control group were fed a basal diet without any supplementation of Met sources. While birds in the treatment groups were received a basal diet supplemented with graded levels (0.075%, 0.10%, 0.125%, 0.15%, 0.175% and 0.2%) of dl-Met (Evonik Industries AG, Hanau, Germany) or l-Met (CJ CheilJedang Co., Seoul, Korea). Both crystal dl-Met and l-Met had 99% purity and they were supplemented to the basal diets at the expense of corn. These test diets eventually contained Met at 75%, 80%, 85%, 90%, 95% and 100% of the NRC requirement (NRC 1994), respectively. Dietary Met in various groups were determined as described previously (Li et al. Citation2018). Briefly, Met was transformed into Met sulphone after oxidation overnight by cold performic acid and then hydrolysed with 7.5 N HCl at 110 °C for 24 h using an Amino Acid Analyser (Hitachi L-8800, Tokyo, Japan). The analysed Met levels in various groups are shown in Table . All birds were raised in battery cages in an environmentally controlled room with continuous incandescent white light throughout the experiment. The room temperature was maintained at 34 °C for the first week, and then reduced by 3 °C per week until it reached 24 °C. The birds had free access to the mashed feed and water throughout the trial period.
Table 1. Nutrient composition of the basal diet (as-fed basis).
Table 2. The analysed values for supplemented methionine (Met) in diets.
Performance measurement
Body weight and feed intake were recorded for each replicate on d 21. Average body weight (ABW) on d 21, along with ADG, average daily feed intake (ADFI), and feed efficiency (FE) from 1 to 21 d of age were calculated.
Measurement of carcass traits
On d 21, two birds (one male and one female) from each replicate cage were randomly selected and euthanized by cervical dislocation after starvation for 12 h. The carcases were plucked and weighed, and nonedible viscera (intestines, proventriculus, gall bladder, spleen, oesophagus, and full crop) were removed. Afterwards, the weight of eviscerated carcase was recorded and various measurements were obtained, including eviscerated weight (EW), breast muscle weight (BMW), leg muscle weight (LMW), eviscerated yield (EY, the percentage of eviscerated carcase weight to slaughter weight), breast muscle yield (BMY, the percentage of breast muscle weight to eviscerated carcase weight), leg muscle yield (LMY, the percentage of leg muscle weight to eviscerated carcase weight), and abdominal fat percentage (AFP, the percentage of abdominal fat weight to eviscerated carcase weight).
Determination of the relative weight of immune organs
On d 21, two birds (one male and one female) from each replicate cage were randomly selected and euthanized. Spleen, bursa of Fabricius and thymus of these birds were removed and weighed for the determination of the relative weight of immune organs, which were expressed as the ratio of organ weight (g) to live body weight (kg).
Biochemical assay of plasma
On d 21, two birds (one male and one female) from each replicate cage were randomly selected for blood collection using heparin tubes. Plasma samples were obtained by centrifugation of blood at 3000 rpm for 10 min at 4 °C, followed by storage at −30 °C until analysis. The plasma uric acid (UA), urea nitrogen (UN) and protein carbonyl (PC) were quantified using commercial assay kits from Institute of Jiancheng Biological Engineering (Nanjing, China). PC content was normalised by total protein level, which was determined by using bicinchoninic acid protein assay kits (Jiancheng Bioengineering Institute, Nanjing, China).
Statistical analysis
Data were presented as mean ± standard error of mean (SEM) and were analysed by ANOVA using the GLM procedure of SAS software. Differences among treatment groups were detected by Duncan’s multiple comparisons. Differences were considered to be statistically significant if p < .05 and were described as tendencies if .05 < p < .10. Polynomial contrasts were also used to determine the linear effects of dl-Met and l-Met levels on response criteria along with the effects of Met sources. Because it is more appropriate to compare the criteria responses of broilers based on the same levels of nutrient intake, the RBA of l-Met to dl-Met was evaluated by slope-ratio assay. The statistical model (exponential regression model) employed in the polynomial regression analysis was the following equation: where y is the response criterion, a is the common intercept, b is the asymptotic response, c1 and c2 are the steepness coefficient for dl-Met and l-Met, respectively, x1 and x2 are the levels of dl-Met and l-Met, respectively. The RBA of l-Met to dl-Met was calculated as the ratio of their steepness coefficients (c2/c1×100) as described previously (Littell et al. Citation1997).
Results and discussion
Growth performance
As shown in Table , increasing supplementation of both l-Met and dl-Met linearly increased (p < .05) ABW, ADG and ADFI of broilers, but linearly reduced (p < .05) FE of birds. Supplemented l-Met induced a higher (p < .05) ADG concomitant with a lower (p < .05) FE of birds as compared with dl-Met. The exponential regression model was found to be propitious to estimate the RBA of l-Met to dl-Met for ADG and FE, whose values were calculated as 141.5% and 189.1%, respectively (Figure ).
Figure 1. Slope-ratio assay of the relative bioavailability of l-methionine (Met) to dl-Met based on average daily gain (ADG) and feed efficiency as the response criteria of broiler chickens from 1 to 21 d of age. Values in brackets indicate 95% confidence intervals. ADG: average daily gain.
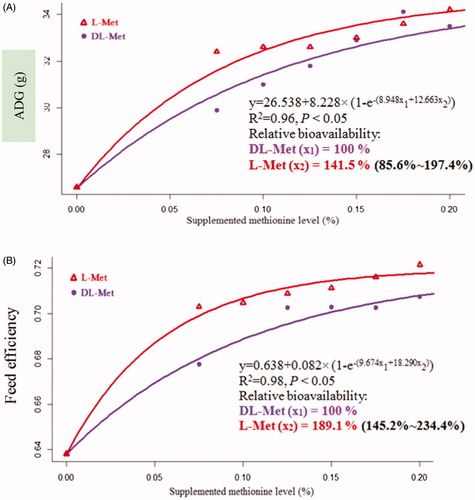
Table 3. Effects of dietary l-methionine (l-Met) and dl-Met supplementation on growth performanceTable Footnoteg in broiler chickens.
Considerable controversies had been surrounding the bioefficacy between dl-Met and l-Met in pigs. Kong et al. (Citation2016) reported that the estimates of RBA of dl-Met to l-Met were 87.9 and 89.3% in nursery pigs, respectively, using retained nitrogen and retention of nitrogen as dependent variables and supplemental Met isomers as independent variable. When ADG was used as the response criterion, the RBA was estimated at 99.6% for l-Met to dl-Met in weaned pigs (Htoo and Morales Citation2016). Besides, Zeitz et al. (Citation2019) indicated that the RBA of l-Met to dl-Met were 99.7% and 99.8% in weaned piglets, respectively, based on the body weight gain and feed conversion ratio as the response criteria, suggesting an identical bioefficacy between dl-Met and l-Met in supporting the growth performance of piglets. Differently, Shen et al. (Citation2014) indicated that the respective RBA of l-Met to dl-Met were 159.1% and 138.5%, based on ADG and FE as the response criteria, respectively, in nursery pigs from d 1 to 10 of the experiment. In chickens, l-Met was found to be superior in growth-promoting efficacy to d-isomer (Baker and Boebel Citation1980). Nevertheless, the RBA of l-Met to dl-Met in broilers remain unclear. In the present study, l-Met addition resulted in higher ADG and FE of broilers compared with dl-Met, and the respective RBA of l-Met to dl-Met for ADG and FE of birds were 141.5% and 189.1%, the latter of which was different from 100%. These demonstrated that supplemental l-Met had an advantage over dl-Met for supporting the growth performance of broilers at an early age (1 to 21 d of age), which might be associated with the higher efficacy of l-Met than dl-Met utilised for protein synthesis in certain tissues such as skin and muscle in chickens (Saunderson Citation1987). In addition to the species of animals, the difference in the age of animals might also be responsible for the discrepancies in the bioefficacy of Met isomers (Cho et al. Citation1980; Shen et al. Citation2014), since research has indicated that that utilisation of Met isomers in animals was likely a function of age (Shen et al. Citation2014). It was found that the expression of d-amino acid oxidase that can convert d-Met into l-Met is very low in young animals (D'Aniello et al. Citation1993). Thereby, there might be an inefficiency of d-Met utilisation by broiler chickens at an early age instead of at the late phase of growth, which could account for the higher bioavailability of l-Met relative to dl-Met for FE of broiler chickens during starter period (1 to 21 d of age) rather than the finisher period (22 to 42 of age) (data not shown).
Carcase traits and relative weight of immune organs
Linear increases (p < .05) in EW, BMW, LMW and BMY, along with a linear reduction (p < .05) in AFP of broilers were observed with the increase of supplementation of both l-Met and dl-Met (Table ). There was a linear increase (p < .05) in LMY of birds responded to the increasing supplementation of l-Met. As compared with dl-Met, l-Met addition led to increases (p < .05) in LMW and LMY coupled with increasing trends (p < .10) of EW and BMW of birds. Based on carcase traits as the response to the addition of Met, the regression model estimated the RBA of l-Met to dl-Met for EW and BMW were 122.9% and 116.8%, respectively (Figure ), however, 95% of the fiducial limits for these RBA estimates included 100%. There were no changes (p > .05) in the relative weight of thymus, spleen and bursa of Fabricius of broilers due to Met addition regardless of the source (Table ).
Figure 2. Slope-ratio assay of the relative bioavailability (RBA) of l-methionine (Met) to dl-Met based on eviscerated weight and breast muscle weight as the response criteria of broiler chickens at 21 d of age. Values in brackets indicate 95% confidence intervals.
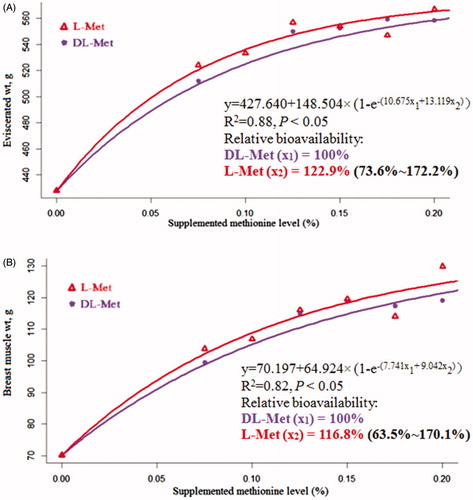
Table 4. Effects of dietary l-methionine (l-Met) and dl-Met supplementation on carcase traitsg in broiler chickens at 21 d of age.
Table 5. Effects of dietary l-methionine (l-Met) and dl-Met supplementation on the relative weightTable Footnotea of immune organs in broiler chickens at 21 d of age.
Carcase traits are particularly important for broiler production. In the present study, supplementing l-Met to the diets elevated or tended to increase EW and BMW, LMW and LMY of broilers compared with dl-Met. One possible explanation for this would be that l-Met rather than d-Met was directly incorporated into protein biosynthesis, resulting in a greater efficiency of l-Met involved in body weight gain of broilers (Saunderson Citation1987). The respective RBA of l-Met to dl-Met for EW and BMW of birds were 122.9% and 116.8%, which, however, were not significantly different from 100%. Measurement of internal immune organ weight is a common method of estimating immune status in animals (Wu et al. Citation2018). In this study, Met addition was likely to elicit little benefit for the immune function of broilers, based on the absence of enlarged relative weight of thymus, spleen or bursa of Fabricius of birds as a result of Met addition.
Plasma biochemical indices
There were linear reductions (p < .05) of UC and PC contents in plasma of broilers due to the increasing supplementation of either l-Met or dl-Met (Table ). Plasma UN content was decreased (p < .05) by l-Met addition without a linear response. Comparatively, increasing dl-Met addition linearly reduced (p < .05) plasma UN content. Birds fed with l-Met had a lower (p < .05) UN content with a higher (p < .05) UA content in plasma than those fed with dl-Met.
Table 6. Effects of dietary l-methionine (l-Met) and dl-Met supplementation on plasma biochemical parameters in broiler chickens at 21 d of age.
Circulating UN and UA contents could be served as influential criteria to reflect protein utilisation in chickens (Khosravinia et al. Citation2015). In the present study, plasma UN and UA contents were all decreased with the increasing supplementation of either l-Met or dl-Met, highlighting an enhanced utilisation efficiency of dietary nitrogen in broilers due to Met addition. It was evidenced that l-Met addition had a higher efficacy than dl-Met to decrease plasma UN content in pigs (Shen et al. Citation2014), possibly due to that l-Met was better utilised by gastrointestinal tract and served as a more efficient substrate for protein metabolism than dl-Met (Shen et al. Citation2014). In the present study, l-Met addition had an advantage over dl-Met in lowering plasma UN content, but was inferior to dl-Met in reducing plasma UA content, implying a complexity of the linkage between dietary Met source and nitrogen metabolism of broilers. PC that produced from the oxidation of amino acid side chains is commonly considered as an indicator for protein oxidation (Dalle-Donne et al. Citation2003). The current reduction of plasma PC content thus suggested alleviated oxidation stress of broilers following Met addition. Nevertheless, we observed no change in plasma PC content of birds as a result of l-Met addition relative to dl-Met, indicating that l-Met and dl-Met were equally bioavailable for attenuating protein oxidation in broilers. This was similar to some previous studies in piglets (Zeitz et al. Citation2019) and broilers (Zhang et al. Citation2018).
Conclusions
The RBA of l-Met to dl-Met for ADG, eviscerated weight and breast muscle weight of broilers from 1 to 21 d of age were 141.5%, 122.9% and 116.8%, respectively. However, these RBA values were not different from100%. When based on FE as the response criterium, the RBA value was calculated as 189.1%, whose fiducial limits excluded 100%. Thereby, l-Met appears to be a more effective source of Met compared with dl-Met for improving FE of broiler chickens in early age.
Disclosure statement
The authors (Xiaoli Dong and Chunyan Guo) may have interests in the submitted work.
Additional information
Funding
References
- Baker DH. 2006. Comparative species utilization and toxicity of sulfur amino acids. J Nutr. 136:1670S–1675S.
- Baker DH, Boebel KP. 1980. Utilization of the D- and L-isomers of methionine and methionine hydroxy analogue as determined by chick bioassay. J Nutr. 110:959–964.
- Brachet P, Puigserver A. 1992. Regional differences for the D-amino acid oxidase-catalysed oxidation of D-methionine in chicken small intestine. Comp Biochem Phys B. 101:509–511.
- Cho ES, Andersen DW, Filer LJ, Jr, Stegink LD. 1980. D-Methionine utilization in young miniature pigs, adult rabbits, and adult dogs. J Parenter Enteral Nutr. 4:544–547.
- D'Aniello A, D'Onofrio G, Pischetola M, D'Aniello G, Vetere A, Petrucelli L, Fisher GH. 1993. Biological role of D-amino acid oxidase and D-aspartate oxidase. Effects of D-amino acids. J Biol Chem. 268:26941–26949.
- Dalle-Donne I, Rossi R, Giustarini D, Milzani A, Colombo R. 2003. Protein carbonyl groups as biomarkers of oxidative stress. Clin Chim Acta. 329:23–38.
- Dilger RN, Baker DH. 2007. DL-Methionine is as efficacious as L-Methionine, but modest L-cystine excesses are anorexigenic in sulfur amino acid-deficient purified and practical-type diets fed to chicks. Poult Sci. 86:2367–2374.
- Fang ZF, Luo HF, Wei HK, Huang FR, Qi ZL, Jiang SW, Peng J. 2010. Methionine metabolism in piglets fed DL-methionine or its hydroxy analogue was affected by distribution of enzymes oxidizing these sources to Keto-methionine. J Agr Food Chem. 58:2008–2014.
- Htoo JK, Morales J. 2016. Bioavailability of L-Methionine relative to DL-Methionine as a methionine source for weaned pig. J Anim Sci. 94:249–252.
- Huang JF, Liu ZQ, Jin LQ, Tang XL, Shen ZY, Yin HH, Zheng YG. 2017. Metabolic engineering of Escherichia coli for microbial production of L-methionine. Biotechnol Bioeng. 114:843–851.
- Khosravinia H, Azarfar A, Sokhtehzary A. 2015. Effects of substituting fish meal with poultry by-product meal in broiler diets on blood urea and uric acid concentrations and nitrogen content of litter. J Appl Anim Res. 43:191–195.
- Kong C, Park CS, Ahn JY, Kim BG. 2016. Relative bioavailability of DL-methionine compared with L-methionine fed to nursery pigs. Anim Feed Sci Tech. 215:181–185.
- Li Y, Lyu Z, Li Z, Liu L, Wang F, Li D, Lai C. 2018. Effects of feeding level and dietary supplementation with crystalline amino acids on digestible, metabolizable and net energy values of corn in growing pigs. Anim Feed Sci Tech. 240:197–205.
- Littell RC, Henry PR, Lewis AJ, Ammerman CB. 1997. Estimation of relative bioavailability of nutrients using SAS procedures. J Anim Sci. 75:2672–2683.
- NRC. 1994. National Research Council. Nutrient Requirements of Poultry: Ninth Revised Edition, Natl. Acad. Press, Washington, DC.
- Riedijk MA, Stoll B, Chacko S, Schierbeek H, Sunehag AL, van Goudoever JB, Burrin DG. 2007. Methionine transmethylation and transsulfuration in the piglet gastrointestinal tract. PNAS. 104:3408–3413.
- Saunderson CL. 1987. Effect of fasting and of methionine deficiency on L-methionine, DL-methionine and DL-2-hydroxy-4-methylthiobutanoic acid metabolism in broiler chicks. Br J Nutr. 57:429–437.
- Shen YB, Weaver AC, Kim SW. 2014. Effect of feed grade L-methionine on growth performance and gut health in nursery pigs compared with conventional DL-methionine. J Anim Sci. 92:5530–5539.
- Stoll B, Henry J, Reeds PJ, Yu H, Jahoor F, Burrin DG. 1998. Catabolism dominates the first-pass intestinal metabolism of dietary essential amino acids in milk protein-fed piglets. J Nutr. 128:606–614.
- Tesseraud S, Metayer-Coustard S, Collin A, Seiliez I. 2009. Role of sulfur amino acids in controlling nutrient metabolism and cell functions: implications for nutrition. Br J Nutr. 101:1132–1139.
- Wu QJ, Zheng XC, Wang T, Zhang TY. 2018. Effects of dietary supplementation with oridonin on the growth performance, relative organ weight, lymphocyte proliferation, and cytokine concentration in broiler chickens. BMC Vet Res. 14:34.
- Zeitz JO, Kaltenböck S, Most E, Eder K. 2019. Effects of L-methionine on performance, gut morphology and antioxidant status in gut and liver of piglets in relation to DL-methionine. J Anim Physiol Anim Nutr. 103:242–250.
- Zhang S, Gilbert ER, Saremi B, Wong EA. 2018. Supplemental methionine sources have a neutral impact on oxidative status in broiler chickens. J Anim Physiol Anim Nutr. 102:1274–1283.