
Abstract
Grazing behaviour influences animal productivity and the conservation of grassland ecosystem services. We used GPS tracking and remote sensing (NDVI index) to monitor the grazing patterns of lactating cows on the ‘Malga Ombretta’ summer farm (1,957 m asl) in the Dolomites, eastern Italian Alps, from 5th July to 5th August 2018. The pasture area (35 ha) was grazed by a mixed herd of Simmental and Alpine grey cows (stocking density = 0.6 LU/ha) under traditional management: each morning the farmer led the cows to graze in a selected sub-area of pasture, and during the afternoon he left them free to graze unrestricted until they returned to the barn for the night. GPS positions were collected every minute from 9 Simmental and 4 Alpine Grey cows with low milk production during the time they were outdoors. The farmer’s choice of where to drive the herd to graze in the morning determined the distances the cows walked/day, which varied from 2.0 to 8.9 km, and favoured the use of higher and steeper areas that the cows tended otherwise to avoid. When free in the afternoon, the cows selected areas with higher NDVI values than those selected by the farmer in the morning, and Alpine Grey cows used slightly higher slopes and altitudes than Simmental cows, suggesting better adaptation to mountain pastures. The study revealed highly heterogenous grazing patterns dependent on multiple factors that can be assessed at fine temporal and spatial scales using GPS and remote sensing technologies to improve grazing management.
Daily distances walked and grazing patterns were influenced differently by the farmer’s decisions and the animals’ choices in response to environmental features.
The NDVI index of vegetation productivity suggested that cows grazed more productive areas when free than when driven by the farmer.
GPS tracking and remote sensing shed light on how human and animal choices regarding grazing are influenced by environmental features.
HIGHLIGHTS
Introduction
Alpine pastures are semi-natural grasslands managed by humans for grazing herbivores, such as cattle (Bunce et al. Citation2004). These high-elevation agroecosystems are exploited during the summer seasonal transhumance and are still important in local livestock farming systems and for product quality, as well as for their biodiversity and delivery of non-provisioning ecosystem services (Sturaro et al. Citation2013; Schils et al. Citation2022). Livestock grazing patterns and their effects on animal welfare and productivity, and on the conservation of pasture ecosystem services depend on a complex set of interactions and trade-offs between animal characteristics (Isselstein et al. Citation2007; Spiegal et al. Citation2019; Pauler et al. Citation2020; Rivero et al. Citation2021), environmental features, such as local morphology (Kaufmann et al. Citation2013; Pittarello et al. Citation2021; Rivero et al. Citation2021), water location (Probo et al. Citation2014), the presence of shelter and shade, and climate conditions (Caton and Olson Citation2016; Liao et al. Citation2017; Rivero et al. Citation2021). These interactions and trade-offs are controlled by the grazing management system adopted. The most extensive continuous grazing systems minimise the control of grazing patterns by leaving livestock free to graze the whole pasture area, and have a low, long-term stocking rate. This requires only low labour and infrastructure investment, but increases livestock movement costs and, due to selective grazing patterns, results in a heterogeneous use of pasture with a mosaic of undergrazed and overgrazed patches, which gradually decreases the forage value of the pasture and allows shrub and tree encroachment (Kothmann Citation2009). At the opposite extreme, grazing patterns in the most intensive rotational systems are controlled by sequential grazing of cows at very high stocking rates in small paddocks for short periods, which restricts the animals’ selectivity but enhances the forage value of the pasture and livestock productivity. However, these systems are labour intensive, require considerable infrastructure investment, and may impact on pasture biodiversity and nutrient cycling (Kothmann Citation2009; Probo et al. Citation2013; Citation2014; Perotti et al. Citation2018).
In order to minimise the trade-off between these two extremes, traditional grazing management in the Alps has developed around extensive rotational systems, where the whole pasture area is divided into relatively large sub-areas where livestock are brought to graze at variable stocking rates and for variable periods based on the shepherd’s and/or expert’s knowledge of the vegetation conditions and evolution (Probo et al. Citation2013; Perotti et al. Citation2018; Pittarello et al. Citation2019). In this case, a thorough understanding of grazing patterns and how they are influenced by both the farmers’ and animals’ choices are fundamental to tailoring grazing management to local conditions to ensure animal welfare and productivity, and conservation of soil and vegetation ecosystem functions (Ravetto Enri et al. Citation2017).
The study of animal movement and behaviour has improved dramatically during recent decades with the introduction of GPS tracking (Cagnacci et al. Citation2010; Nathan et al. Citation2022). This technology allows the position of individual animals to be monitored with high time-frequency resolution on a scale as small as seconds or minutes, and high spatial accuracy with location errors of <5–10 m (D’Eon et al. Citation2002; Tomkiewicz et al. Citation2010; Muminov et al. Citation2019), and without interference with the animals’ behavioural patterns by an observer (Homburger et al. Citation2014). GPS tracking has therefore been used extensively to study the movement ecology of wildlife (Nathan et al. Citation2022) and more recently, but to a lesser extent, to study livestock grazing patterns (see the reviews by Bailey et al. Citation2018 and Rivero et al. Citation2021). GPS location technology can be combined with remote sensing technologies, which are able to characterise environmental conditions at multiple spatial and temporal scales from satellite images and provide a more detailed understanding of animal-habitat interactions (Pettorelli et al. Citation2005, Citation2014). A widely-used remote sensing index is the absolute Normalised Difference Vegetation Index (NDVI) (Myneni et al. Citation1995; Shariatinajafabadi et al. Citation2014). The NDVI ranges from −1 to +1, where values below 0 are typical of habitats without vegetation, while values close to 1 are indicative of areas with very abundant vegetation (Pettorelli et al. Citation2005). It is therefore used to estimate vegetation biomass, but, in addition, temporal variations in the index can also be used to estimate the phenological stage and stress conditions of plants (Myneni et al. Citation1995; Pettorelli et al. Citation2005; Shariatinajafabadi et al. Citation2014).
In this study, we used GPS tracking on a small temporal scale to investigate the movement patterns of lactating cows in an alpine pasture managed according to a combination of rotational and continuous grazing. Specifically, we aimed to describe the animals’ movement patterns and use of pasture at two spatio-temporal scales, i.e. the single movement step, defined as the segment connecting two consecutive locations, and the total daily trajectory, and assess how they were influenced by the farmer’s decisions and the animals’ selectivity in relation to land morphology, weather conditions, vegetation type and vegetation productivity (according to the NDVI). In addition, we took advantage of the presence of two breeds of cows, Alpine Grey and Simmental, to obtain a preliminary indication of possible differences between them.
Material and methods
Study area and summer farm management
The study was conducted in Val Ombretta, located in the Marmolada massif in the eastern Italian Alps (Dolomites; 46°26′13′′ N, 11°51′54′′ E), during the summer of 2018 (Figure ). Val Ombretta is characterised by a typical alpine environment with subalpine grasslands and sparse forest composed mostly of Larix decidua with also Pinus mugo and Picea abies, surrounded by rocky cliffs that reach an altitude of over 3,000 m asl. It has approximately 35 ha of grazing land at an average altitude of 1,957 m asl (SD = 15) and with an average slope of 16.5° (SD = 6.1).
Figure 1. Val Ombretta showing the location of the summer farm, the total grazing area comprising areas used freely by the cows in the afternoon (in blue, ‘Farmer absence’), the sub-areas where the farmer conducted the cows in the morning (in red, ‘Farmer presence’), and the positions of drinking troughs.
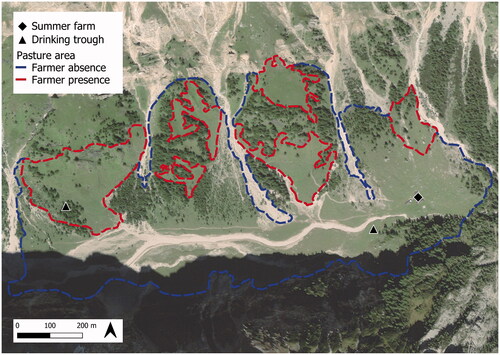
In summer − from mid-June until mid-September − the grasslands of the valley are grazed by a herd of dairy cattle managed by the local summer farm. During the study period, the summer farm hosted 14 Simmental and 7 Alpine Grey lactating cows at an average stocking density of 0.6 livestock units/ha. The herd was managed traditionally. After the evening milking (starting 5.30–6 pm), the cows spent the night inside the barn in fixed stalls. After the morning milking (ending around 8–8.30 am), they were conducted by the farmer to selected pasture sub-areas (Figure ), where they grazed until lunch time (around 12 am–1 pm), after which the farmer left them free for the afternoon until they returned spontaneously to the barn for the evening milking. The cows received a very limited amount (0.5–0.8 kg/d) of a compound feed supplement (19.0% crude protein, 6.4% crude fibre, 9.2% total ash; Palumbo et al. Citation2021), so they had to rely mostly on grazing to fulfil their energy requirements.
The study included 9 Simmental and 4 Alpine Grey cows, which were all multiparous. The summer farm was visited once during the study period and milk yield was estimated from the volume of milk collected in a graduated bucket (0.5 l resolution) at the morning and evening milking of each cow. On the same visit, body length (BL), girth circumference (GC) and body condition scores (BCS) were measured by a single trained operator. The values obtained were used to estimate live body weight according to the equation −1099.5 + 3.37*BL + 5.47*GC + 25.3*BCS (L. Gallo, unpublished data).
Spatial covariates and time-varying spatial covariates
Digital maps of the study area were generated in QGIS 3.14 (http://www.qgis.osgeo.org/) using the EPSG 4326 coordinate system. We created a raster map of slope (degrees) and altitude (m) using a Digital Elevation Model with a resolution of 10 m (https://www.regione.veneto.it/web/ambiente-e-territorio/ ctr). We created a categorical variable ‘slope class’ with four levels: <10°, ≥10° and <20°, ≥20°and <30° and ≥30°. We used the map produced by Scillitani et al. (Citation2013) with a spatial resolution of 50 m to categorise the land cover of the grazing area into three ‘habitat types’: ‘Larix’ (Larix decidua stands), ‘grassland’ (subalpine pastures), and ‘scree’ (areas covered by loose stone with little vegetation). We obtained NDVI values as an indicator of vegetation productivity of the study area from the Sentinel-2 constellation of satellites using the Google Earth Engine open-source platform (Gorelick et al. Citation2017). The NDVI raster maps were acquired at a 10 m resolution for the dates 5th, 8th and 13th July, according to the temporal resolution of Sentinel-2 spectral acquisition. We generated daily NDVI maps by interpolating the daily variation in NDVI across the three dates. We created a categorical variable ‘NDVI class’with four levels: <0.2, ≥0.2 and <0.4, ≥0.4 and <0.6, ≥0.6. We also obtained average hourly temperatures and precipitation from the nearest weather station (Malga Ciapela station, 1,475 m asl; Rocca Pietore BL ARPAV – Veneto Regional Agency for Environmental Protection and Prevention, https://www.arpa.veneto.it/) for every day of the study period. Because the temperatures were not recorded at the summer farm itself, but at a lower altitude, we created a categorical variable ‘temperature class’ with three levels: ‘low’ (<13.1 °C, first quartile); ‘average’ (≥13.1 and <17.4 °C, second and third quartiles); ‘high’ (≥17.4 °C, fourth quartile), which means the values of this variable are relative, rather than absolute. We also categorised precipitation as ‘rain occurrence’ (yes/no) as there were many days without rain.
Movement patterns of cows
The movement patterns of the cows were monitored with GPS collars (VERTEX Plus model; VECTRONIC Aerospace GmbH) scheduled to attempt a location every minute from 08:00 am to 6:00 pm from 5th July to 5th August. The median location error had been previously estimated at 4.5 m (Parraga Aguado et al. Citation2017). We obtained 198,186 GPS positions, which we pre-processed in PostgreSQL 14 (https://www.postgresql.org/docs/14/index.html) with the plugin PostGIS 3.1.5 (http://postgis.net/2022/02/01/postgis-3.1.5/). After excluding impossible locations (e.g. on steep rocky slopes or in other valleys), we associated to the second location of each ‘movement step’ (i.e. each consecutive pair of locations): (a) the ‘step distance’, calculated as the linear distance in m between the first and second locations corrected for the slope travelled as indicated by the elevation difference between them; (b) the ‘speed’, calculated as step distance/time interval in sec; (c) the ‘turning angle’, i.e. the cosine of the angle between the line intersecting the two locations of one step and the line intersecting the first location of that step with the second location of the preceding step (Urbano and Cagnacci Citation2014). We then identified the remaining outlier locations by first excluding those with an impossible speed (>15 km/h), as suggested by Liao et al. (Citation2017) and Spiegal et al. (Citation2019), and then those with an impossible combination of speed and turning angle (speed >2.4 km/h and cosine <–0.97), as suggested by Urbano and Cagnacci (Citation2014). Finally, since the daily schedule of data collection by the GPS collars (8:00 am–6:00 pm) could have included periods when the cows were in the barn, we identified the indoor period for each cow on each day using a combination of the frequency of missed locations (when the GPS is inside a building most positions are lost due to obstruction of the satellite signals) and movement trajectories. Specifically, we assumed that the indoor period started in the first 10-min time interval after 4:30 pm with >5 missed positions (out of 10 expected), and ended in the last interval after 8:00 am with >5 missed positions. We checked that all the 10-min intervals within the starting or ending intervals exhibited consistently high numbers of missed positions. To avoid removing any real outdoor positions from the starting and ending intervals, we deleted from these only the positions that were within a 25 m buffer surrounding the barn. Finally, we visually checked the continuity of the resulting individual outdoor daily movement trajectories. The final geodatabase contained 174,171 outdoor locations with each location associated with individual features (individual cow and breed), temporal features (Julian date and hour), environmental features (slope, elevation, land cover type and NDVI class), climate conditions (hourly and daily average temperature, temperature class, rain occurrence) and step movement features (step distance, speed). We also calculated for each day and each cow the ‘outdoor time’ (in h) as the difference between the time of the first and the last outdoor locations, and divided this into two ‘day periods’ using 12.00 am as the boundary: ‘morning’ (before 12.00 am), when the cows’ movement patterns are determined by the farmer, and ‘afternoon’ (after 12.00 am), when no restrictions are placed on the cow on where to graze and when to return to the barn. For each day and cow, we also calculated: (a) the ‘distance walked in the morning’ as the sum of the step distances of the morning trajectory, and the ‘distance walked daily’ as the sum of the step distances of the morning and afternoon trajectories; and (b) the ‘daily vertical movement’, as the difference (in m) between the lowest and highest elevation/cow/day.
Statistical analysis
The statistical analyses were conducted in R 4.0.2 (R Core Team Citation2016). As a preliminary, we assessed possible differences in milk yield and live body weight between the two breeds with a simple one-way ANOVA.
Daily and hourly movement variables
Daily movement-related variables (distance walked and vertical movement) were analysed with linear mixed effects models using the lmer function of the lme4 package (Bates et al. Citation2015). The models included the continuous linear effects of Julian date, distance walked in the morning, outdoor time, and (only for daily distance walked) daily vertical movement, the categorical effects of breed, rain occurrence and temperature class, and the random effect of individual cow. As a preliminary, we checked for absence of collinearity between the explanatory variables (all VIF values were below 2.6). We assessed the models’ marginal R2, due to fixed factors only, and conditional R2, due to fixed plus random factors (Nakagawa and Schielzeth Citation2013), using the Performance package (Lüdecke et al. Citation2021).
Hourly movement-related variables (slope, altitude and speed associated to each location/step) were analysed with generalised additive mixed models using the gam function of the mgcv package (Wood Citation2017), with a model that included the smoothed spline effect of hour, the categorical effects of breed, temperature class, rain occurrence and day period, the two-way interactions of day period with breed, temperature class or rain occurrence, and the random effects of Julian date and individual cow. Speed was expressed as m/h, and slope and speed were log-transformed before analysis.
Use of pasture and resource selection
We assessed the spatial evenness of the use of the pasture area during each day period by calculating Camargo’s index (Payne et al. Citation2005; Pauler et al. Citation2020) on the total GPS positions acquired during the mornings and afternoons of the entire study period within each cell of a 25 × 25 m grid overlaid onto the pasture area. The index ranges from 0, patchy or heterogeneous use of an area, to 1, homogeneous use.
To assess the selection – i.e. the preferential use of a resource limited by external conditions (Manly et al. Citation2002) – of spatial features (slope, NDVI, habitat type) we used a resource selection function approach (Boyce and McDonald Citation1999). We first extracted the areas ‘available’ to the cows in each day period of each date by calculating the minimum convex polygons that included the morning and afternoon locations of the monitored cows from 5th to 13th July. From within each polygon we extracted the ‘non-used area’ by excluding the area not forming part of the 25 m buffer around each acquired location. Within the non-used area, we generated a number of random ‘non-used positions’ equal to the number of acquired animal locations and assigned them randomly to each cow. Thus, we obtained a database of ‘used’ (with animal locations) and ‘non-used’ (without animal locations) positions, which we categorised by NDVI class, slope class and habitat type (simplified to ‘grassland’ and ‘forest’, excluding ‘scree’, which was very seldom used). Finally, we used a generalised linear mixed effects model with binomial distribution and logit link function (glmer function of the lmer4 package (Bates et al. Citation2015)) to assess the relative probability of a position being used in response to the fixed effects of habitat type, slope class, NDVI class and day period, the two-way interactions between period and the other effects, and the random effect of the individual cow. We assessed the model’s marginal R2, due to fixed factors only, and the conditional R2, due to fixed plus random factors (Nakagawa and Schielzeth Citation2013) with the Performance package (Lüdecke et al. Citation2021).
Results
Milk production and live body weight
Milk production was low and did not significantly differ between breeds (GLM least square means: Alpine Grey: 10.9 kg/day, SE = 2.1; Simmental: 11.2 kg/day, SE = 1.4; p = 0.63), nor did predicted live body weight (GLM least square means: Alpine Grey: 537.9 kg, SE = 39.3; Simmental: 553.0 kg, SE = 26.2; p = 0.74). Therefore, we assumed that any differences in movement patterns between breeds would not be related to milk yield and live body weight.
Daily movement-related variables
The average distance walked daily was 4,585 m (SD = 206, minimum = 2,019, maximum = 8,912), while the average daily vertical movement was 101 m (SD = 51 m, minimum = 18 m, maximum = 225 m).
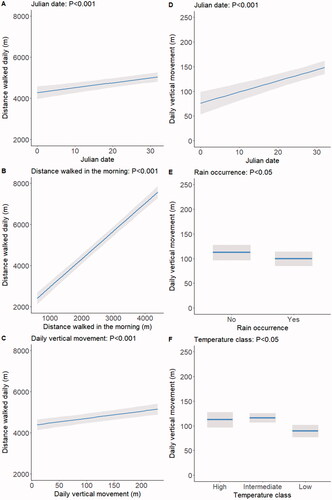
For brevity, the parametric coefficients of the models analysing the two variables are given in Supplementary Table S1. The distance walked daily was unaffected by breed, rain occurrence and temperature class (P values ranging between 0.26 and 0.73) and increased only tendentially with outdoor time (p = 0.06). However, it increased significantly (p < 0.001) with Julian date and mainly, although unsurprisingly, with distance walked in the morning (Figure , panels A and B). Longer distances walked daily were also associated (p < 0.001) with higher daily vertical movement (Figure , panel C). Daily vertical movement did not vary significantly in response to distance walked in the morning, breed and outdoor time (p values ranging between 0.25 and 0.79) but increased markedly with Julian date (p < 0.001; Figure , panel D). It was also negatively affected (p < 0.05) by rain occurrence and temperature class (Figure , panels E and F).
Figure 2. Effects on distance walked daily of Julian date, distance walked in the morning and daily vertical movement (panels A, B, and C, respectively), and effects on daily vertical movement of Julian date, rain occurrence, and temperature class (panels D, E, and F, respectively). Shaded areas indicate 95% confidence intervals. Only significant effects are shown; for details of the parametric coefficients of the statistical models see Supplementary Table S1.
Hourly movement-related variables
The parameters of the models analysing slope used, altitude used and speed are reported in Supplementary Table S2. Slope used followed a spiked pattern with peaks at around 11 am and 4 pm and troughs at around 9 am and 2 pm (Figure , panel A, smoothed spline effect of hour; p < 0.001). Alpine Grey cows used steeper slopes than Simmental cows, especially in the afternoon (Figure , panel B, breed by day period interaction; p < 0.05). The cows used a markedly gentler slope in the morning than in the afternoon in the presence of rain, and a slightly steeper slope in the absence of rain (Figure , panel C, rain class by day period interaction; p < 0.001). At high temperatures the cows grazed on a much gentler slope in the afternoon than in the morning, but the difference was smaller at low temperatures and non-existent at intermediate temperatures (Figure , panel D, temperature class by day period interaction; p < 0.001).
Figure 3. Effects of the smoothed spline of hour, and of the interactions breed by day period, rain occurrence by day period, and temperature class by day period on the slope (panels A, B, C and D, respectively) and on the altitude used (panels E, F, G and H, respectively), and effects of the smoothed spline of hour, breed, rain occurrence and the temperature class by period interaction on speed (panels I, J, H and K, respectively). Shaded areas indicate 95% confidence intervals. The vertical dotted line in panels A, E and I indicate the separation between the morning and afternoon day periods at 12 am. For details of the parametric coefficients of the statistical models see Supplementary Table S2.
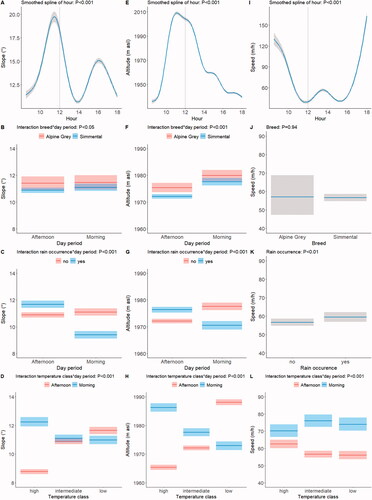
The altitude used increased rapidly during the morning, almost paralleling the increasing pattern of slope, but in the afternoon showed a constant decline (Figure , panel E, smoothed spline effect of hour; p < 0.001) The Alpine Grey cows used a higher altitude than the Simmental cows, especially in the afternoon (Figure , panel F, breed by day period interaction; p < 0.001). The occurrence of rain was accompanied by lower altitudes used in the morning, and by higher altitudes in the afternoon (Figure , panel G, rain occurrence by day period interaction; p < 0.001). Higher temperatures were accompanied by higher altitudes in the morning, but by lower altitudes in the afternoon, while the opposite pattern was observed at lower temperatures (Figure , panel H, temperature class by day period interaction; p < 0.001).
The cows left the barn at high speed, then gradually slowed until 12 am when they reached the maximum slopes and altitudes; there was then a period of slow movement until 4 pm, followed by increasing speed until they returned to the barn (Figure , panel I, smoothed spline effect of hour; p < 0.001). There were no differences between breeds (Figure , panel J; p = 0.94), and no breed by day period interaction (p = 0.33). The occurrence of rain reduced speed (Figure , panel K; p < 0.01). Finally, speed was faster in the morning than in the afternoon, but this difference was less marked at high temperatures as morning speed was slower and afternoon speed was faster than at intermediate or low temperatures (Figure , panel L, temperature class by day period interaction; p < 0.001).
Use of pasture and resource selection
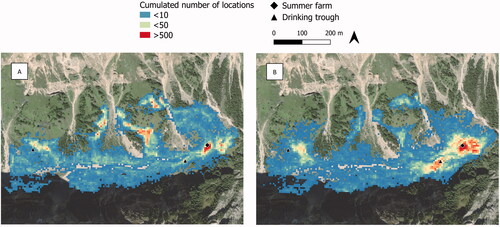
Camargo’s index (0.11 ± 0.06 in the morning, and 0.13 ± 0.07 in the afternoon) indicated a highly uneven use of the pasture, with greater use in the steeper, peripheral areas in the morning, and in the flatter areas close to the barn in the afternoon (Figure ).
Figure 4. Intensity of use of pasture (no. of locations cumulated over the study period within a 10 × 10 m grid) in the morning (A) and afternoon (B).
The probability of a position being used increased markedly with increasing NDVI values (see Supplementary Table S3 for the parametric coefficients of the model), but in the afternoon the cows grazed patches with high NDVI more frequently and patches with low NDVI less frequently than in the morning (Figure , panel A, NDVI class by day period interaction; p < 0.01). Slope had little influence on the probability of a position being used in the morning, but in the afternoon this probability was clearly lower for slopes steeper than 20 degrees (Figure , panel B, slope class by day period interaction; p < 0.001). Finally, there was a strong preference for areas of grasslands over areas of Larix decidua stands, but less so in the afternoon than in the morning (Figure , panel C, habitat type by day period interaction; p < 0.001).
Figure 5. Predicted probability of use of pasture area according to the two-way interactions between day period and NDVI class, slope class and habitat type. Shaded areas indicate 95% confidence intervals.
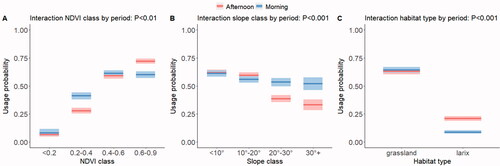
Figure 5. Predicted probability of use of pasture area according to the two-way interactions between day period and NDVI class, slope class and habitat type. Shaded areas indicate 95% confidence intervals
Discussion
In this study, we identified the role played by the farmer’s daily decisions in determining grazing patterns in a mixed rotational/continuous management system, and at the same time showed how cows seek an energy landscape characterised by low cost (gentler slope, lower elevations) and high forage biomass (according to NDVI values). However, the resulting use of the pasture areas remained highly heterogeneous. In the following sections, we will discuss these findings.
The importance of the farmer’s decisions
The farmer’s daily decision regarding which pasture sub-area to conduct the herd to in the morning had a major influence on the distance the cows walked daily, as indicated by the distance walked in the morning, but also on the daily vertical movement and slopes used, as evidenced by the fact that the cows reached the highest altitudes (>1,950 m asl) and steepest slopes (>20 degrees) around 11 am. There was substantial variability in the distances covered daily by the cows, and in their daily vertical movement, and consequently slopes used. Since these variables are positively related, there was also considerable daily variation in movement costs, and hence maintenance requirements. There is a surprisingly small literature on the energy costs of the movement of grazing livestock. According to the Agricultural Research Council (Citation1980), the energy costs associated with cattle walking 1 km horizontally and 0.1 km vertically are 2.0 and 2.8 kJ/kg body weight, respectively. In our study, the minimum distance walked daily was 2.0 km with a vertical movement of 0.02 km, while the maximum was 8.9 km with a vertical movement of 0.16 km. For a cow with a body weight of 550 kg this would, according to ARC (Citation1980), correspond to energy costs of 2.5 and 12.3 MJ, which would increase maintenance requirements (Nozière et al. Citation2017) by approximately 7 and 34%, respectively. These additional costs are within the range suggested by the Standing Committee on Agriculture (SCA) (Citation1990) for grazing cows, but the considerable variability between days cannot be compensated for on summer farms where supplementary feeding is very limited and cows have to rely on herbage intake to fulfil their needs, as in our case study.
Herd management also influenced the patterns of speed. The cows moved faster soon after leaving the barn in the morning when the farmer conducted them to the selected grazing areas. They then gradually slowed down as they increased their grazing activity until noon, when the animals, now left free by the farmer, increased their speed to reach the drinking troughs. Afterwards, they reduced speed again for 2–3 hours, before starting to move faster to reach the barn for the evening milking. In this symmetrical diurnal pattern, the cows moved faster in the morning, when the farmer was driving them, than in the afternoon.
Our results also suggest that the selection of pasture sub-areas by the farmer was influenced by weather conditions. Daily vertical movements were smaller on days with rain and low temperatures than on days without rain and with intermediate or high temperatures, which appears to be a consequence of lower altitudes (and consequently gentler slopes) being used in the morning than in the afternoon. It seems, therefore, that the farmer decided to stay closer to the bottom of the valley when the weather was less favourable. Furthermore, the positive relationship between daily horizontal and vertical movement and Julian date probably reflects an attempt by the farmer to exploit the more peripheral areas with the advancing season, possibly because the more accessible areas had already been exploited. This is consistent with Palumbo et al. (Citation2021) findings that in the same pasture the average forage value of the plant species ingested by grazing cows gradually declined over the grazing period because of an increase in the proportion of species with low palatability and low forage value.
Finally, analyses of movement variables on an hourly scale and of resource selection revealed that in various instances the farmer’s decisions were in contrast to the cows’ choices. This is important to consider in addressing grazing management and will be discussed in the next section.
The animal component remains important
It is well known that in terms of body size/morphology, behaviour and performance levels, traditional local breeds are more suited to grazing in harsher areas than conventional and especially highly productive breeds (Hessle et al. Citation2014; Zendri et al. Citation2016). However, comparisons of the movement patterns of different breeds or their crosses in European mountain pastures are rare and have involved genotypes that are more divergent in body size and productivity than those compared here (Hessle et al. Citation2008; Pauler et al. Citation2020). Although our results are to be treated with caution and considered preliminary, requiring verification with larger sample sizes with a wider variation in body mass and milk yield and including factors, as parity, which we could not test here, they suggest that the local Alpine Grey has a better ability to move quickly and on steeper slopes than the Simmental. Alpine Grey cows also used higher altitudes, and although the difference was very modest in absolute values (around 10 m) it indicates spatial separation between individuals of the two breeds. Overall, these findings suggest that Alpine Grey are more suited than Simmental to grazing alpine pastures. In a study involving multi-breed herds on 15 alpine summer farms, Zendri et al. (Citation2016) found that lactating cows of local dual-purpose breeds (Alpine Grey, Rendena and crossbreds) maintained higher body condition scores during the season than Simmental cows, whose milk yields were higher at the beginning of the summer grazing periods, but similar to the other breeds at the end.
In general, land morphology and the spatial distribution of vegetation types and forage values influence animals’ movement patterns in interaction with climate conditions (Rivero et al. Citation2021). We also found an effect of land morphology in our study. In the afternoon, when left free by the farmer, the cows showed a clear preference for gentler slopes. This was expected, since steep slopes and rocky or soft soils are avoided by cattle due to the increased energy cost of movement and risk of injuries (Kaufmann et al. Citation2013; Probo et al. Citation2014; Pittarello et al. Citation2021; Rivero et al. Citation2021). In our study, the gentler slopes and lower altitudes were also used more in the afternoon on hotter and sunny days, which might be to reduce energy expenditure and avoid the risk of heat stress. However, we could not test this hypothesis directly because we did not have the actual temperatures at the summer farm, which would probably have been lower than those we used. Furthermore, we do not have an explanation for why the cows moved faster in the afternoon and more slowly in the morning on hotter days.
The analysis of habitat selection showed a strong preference for grassland patches over Larix decidua stands, consistent with the general observation that cattle prefer open grassland to shrub or forest habitat for grazing (Meisser et al. Citation2014; Spedener et al. Citation2019; Tofastrud et al. Citation2019). Interestingly, however, the cows had a greater tendency to avoid the Larix decidua stands during the morning, when their grazing area was chosen by the farmer, than in the afternoon, when it was chosen by the cows. This indicates that humans and animals have different perceptions of this habitat. Larix decidua stands have a loose canopy cover that allows ground vegetation to grow, and because they offer both forage and shade or shelter they are often grazed (Da Ronch et al. Citation2016). Possible explanations for the higher use of Larix decidua in the afternoon could be that the cows simply passed through the forest on their way from the high areas to the valley bottom, or alternatively that they deliberately made greater use of this habitat during the afternoon, for instance in search of shade while continuing feeding. This last hypothesis could be tested with accurate and more variable temperature values, and by investigating the types of activities the animals engage in (Homburger et al. Citation2014, Citation2015).
The NDVI has been used as an indicator of vegetation productivity, especially on rangelands, often at large spatial and temporal scales (Browning et al. Citation2018; Spiegal et al. Citation2019). It has been shown that cattle have a clear preference for higher NDVI values (>0.5) (Manning et al. Citation2017). In this study we found a similar trend, suggesting that the index can be used to shed light on grazing patterns, even at small spatial and temporal scales. Interestingly, the areas selected by the cows in the afternoon had higher NDVI values than the areas selected by the farmer in the morning. This was not related to the greater use of Larix decidua stands, which had on average a lower NDVI value than the grasslands (least squared means of a one-way ANOVA: grassland = 0.540, SE = 0.001; Larix decidua = 0.507, SE = 0.002, p < 0.001), and therefore seems to reflect the cows’ ability to graze in (grassland) areas with richer vegetation than those selected by the farmer. While absolute NDVI values reflect vegetation biomass, relative variations in them over time index phenology, and hence nutritional quality, increasing rapidly during the early growth phases, more slowly towards maturation, and decreasing during senescence (Beck et al. Citation2008; Shariatinajafabadi et al. Citation2014). For this reason, various studies on wild species have found that movement and foraging patterns have a positive relationship with vegetation quality as indexed by NDVI variation (Geremia et al. Citation2019; Semenzato et al. Citation2021). To our knowledge, this has not been replicated with domestic species and we were unable to do so in this study because we had too few NDVI time intervals, although it is a promising topic for future research.
The use of pasture is highly heterogeneous
Our assessment of pasture use is partial because we did not monitor the whole grazing season, but it clearly suggests that the combination of the farmer’s daily decisions and the cows’ preferences when left free resulted in highly variable stock densities over the grazed area. The morning hotspots reflected the farmer’s attempts to use areas at higher elevations and greater distances from the barn that the cows would otherwise probably not have used, while the afternoon hotspots were influenced in part by those of the morning, but mainly reflected the cows seeking the valley bottom, the water troughs and proximity to the barn. This pattern illustrates the difficulty faced by non-intensive grazing management systems in reducing heterogeneity in the use of pasture (Probo et al. Citation2013; Pittarello et al. Citation2019), which has various potential consequences. In pasture ecosystems, livestock presence is a local disturbance because the structure and composition of the vegetation can be shaped by selective herbivory (Isselstein et al. Citation2007; Rivero et al. Citation2021), and soil abiotic conditions and biotic functions can be modified by trampling and nutrient translocation through excreta (Teague et al. Citation2011; Peco et al. Citation2017; Koch et al. Citation2018). Undergrazed areas may suffer from depletion of nutrients, progressive shrub or tree encroachment, and even abandonment (Koch et al. Citation2018) and, in fact, visual observations of this pasture clearly reveal an ongoing expansion of Larix decidua towards the valley bottom. Overgrazed areas, on the other hand, may be impacted by erosion and depletion of grass vegetation – indeed vegetation sampling (Stefano Macolino, unpublished results) has indicated that standing grass biomass is very low in proximity to the barn during the grazing season −, and an excess of nutrients (Teague et al. Citation2011; Sartorello et al. Citation2020). In order to address the complexity of these interactions, the possibility offered by GPS tracking to accurately map the variation in animal stock density at a fine spatial scale is clearly of considerable interest. The data it yields may be used to improve grazing management, address the identification and rotation of grazing sub-areas or paddocks, and also the placement of points of attraction for the animals, such as water troughs and mineral supplements (Probo et al. Citation2013).
Conclusion
This study provides an example of the variability in grazing patterns that can be found in alpine summer pastures managed with a combination of extensive rotational and continuous grazing, an increasingly common practice in recent decades in the Alps. The farmer’s driving of the herd in the morning was fundamental to using pasture areas that under continuous grazing management would have been avoided by the cows. The daily selection of these areas, which was influenced by weather conditions, was a determining factor in the variability in energy costs associated with animal movement and resulted in the cows using less productive grassland patches than those used when grazing freely in the afternoon. In addition, the intensity of use of the pasture area was highly heterogeneous. Given the wide diversity in the environmental and management conditions of alpine pastures, and pastures managed extensively in general, in other contexts an even greater variability in grazing patterns than found in this study may be expected. Describing this variability is important for addressing the multifunctional management of pastures, which needs to combine animal welfare and productivity with the conservation of ecosystem services, both provisioning (i.e. forage production) and non-provisioning (e.g. biodiversity, carbon and nutrient balance, aesthetic and cultural values). In this regard, our study shows that the rapidly expanding GPS and remote sensing technologies may be used not only to understand spontaneous animal grazing patterns and to index land morphology and grassland productivity, but also to verify farmers’ choices and their consequences. In geographic contexts other than Europe, this knowledge and technology is being increasingly expanded and used to develop precision grazing management tools. We suggest that further studies in different alpine contexts, with larger sample sizes and a wider range of environmental conditions (e.g. the actual temperature animals are exposed to) would provide useful information for improving grazing management of alpine pastures.
Ethical approval
The study was approved by the ethical committee of the University of Padova with prot. number 389576 09/11/2017.
Supplemental Material
Download MS Word (29.9 KB)Acknowledgments
We thank the farmers of Malga Ombretta for their support and Dr. Luca Carraro for taking measurements and calculating body condition scores.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data of this study are freely available from the corresponding author upon request. The data are not publicly available due to the involvement of private partners (farmers).
Additional information
Funding
References
- Agricultural Research Council. 1980. The nutrient requirements of ruminant livestock. Slough, UK: CAB.
- Bailey DW, Trotter MG, Knight CW, Thomas MG. 2018. Use of GPS tracking collars and accelerometers for rangeland livestock production research. Transl Anim Sci. 2(1):81–88.
- Bates D, Mächler M, Bolker B, Walker S. 2015. Fitting linear mixed-effects models using lme4. J Stat Softw. 67(1):1–48.
- Beck PSA, Wang TJ, Skidmore AK, Liu XH. 2008. Displaying remotely sensed vegetation dynamics along natural gradients for ecological studies. Int J Remote Sens. 29(14):4277–4283.
- Boyce MS, McDonald LL. 1999. Relating populations to habitats using resource selection functions. Trends Ecol Evol. 14(7):268–272.
- Browning DM, Spiegal S, Estell RE, Cibils AF, Peinetti RH. 2018. Integrating space and time: a case for phenological context in grazing studies and management. Front Agr Sci Eng. 5(1):44–56.
- Bunce RGH, Pérez-Soba M, Jongman RH, Gómez Sal A, Herzog F, Austad I. 2004. Transhumance and biodiversity in european mountains. IALE publication series nr 1, Wageningen UR.
- Cagnacci F, Boitani L, Powell RA, Boyce MS. 2010. Animal ecology meets GPS-based radiotelemetry: a perfect storm of opportunities and challenges. Philos Trans R Soc Lond B Biol Sci. 365(1550):2157–2162.
- Caton JS, Olson BE. 2016. Energetics of grazing cattle: impacts of activity and climate. J Anim Sci. 94(suppl_6):74–83.
- Da Ronch F, Caudullo G, Tinner W, de Rigo D. 2016. Larix decidua and other larches in Europe: distribution, habitat, usage and threats. Eur Atlas For Tree Species. 2016:108–110.
- D’Eon RG, Serrouya R, Smith G, Kochanny CO. 2002. GPS radiotelemetry error and bias in mountainous terrain. Wildl Soc Bull. 30(2):430–439.
- Geremia C, Merkle JA, Eacker DR, Wallen RL, White PJ, Hebblewhite M, Kauffman MJ. 2019. Migrating bison engineer the green wave. Proc Natl Acad Sci USA. 116(51):25707–25713.
- Gorelick N, Hancher M, Dixon M, Ilyushchenko S, Thau D, Moore R. 2017. Google earth engine: planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 202:18–27.
- Hessle A, Dahlström F, Bele B, Norderhaug A, Söderström M. 2014. Effects of breed on foraging sites and diets in dairy cows on mountain pasture. Int J Biodivers Sci Ecosyst Serv Manag. 10(4):334–342.
- Hessle A, Rutter M, Wallin K. 2008. Effect of breed, season and pasture moisture gradient on foraging behaviour in cattle on semi-natural grasslands. Appl Anim Behav Sci. 111(1–2):108–119.
- Homburger H, Lüscher A, Scherer-Lorenzen M, Schneider MK. 2015. Patterns of livestock activity on heterogeneous subalpine pastures reveal distinct responses to spatial autocorrelation, environment and management. Mov Ecol. 3(1):1–15.
- Homburger H, Schneider MK, Hilfiker S, Lüscher A. 2014. Inferring behavioral states of grazing livestock from high-frequency position data alone. PLoS One. 9(12):e114522–22.
- Isselstein J, Griffith BA, Pradel P, Venerus S. 2007. Effects of livestock breed and grazing intensity on biodiversity and production in grazing systems. 1. Nutritive value of herbage and livestock performance. Grass Forage Sci. 62(2):145–158.
- Kaufmann J, Bork EW, Blenis PV, Alexander MJ. 2013. Cattle habitat selection and associated habitat characteristics under free-range grazing within heterogeneous Montane rangelands of Alberta. Appl Anim Behav Sci. 146(1–4):1–10.
- Koch B, Homburger H, Edwards PJ, Schneider MK. 2018. Phosphorus redistribution by dairy cattle on a heterogeneous subalpine pasture, quantified using GPS tracking. Agric. Ecosyst. Environ. 257:183–192.
- Kothmann M. 2009. Grazing methods: a viewpoint. Rangel. 31(5):5–10.
- Liao C, Clark PE, Degloria SD, Barrett CB. 2017. Complexity in the spatial utilization of rangelands: pastoral mobility in the Horn of. Africa Appl Geogr. 86:208–219.
- Lüdecke D, Ben-Shachar MS, Patil I, Waggoner P, Makowski D. 2021. Performance: an R package for assessment, comparison and testing of statistical models. J Open Source Softw. 6(60):139.
- Manly BFL, McDonald L, Thomas DL, McDonald TL, Erickson WP. 2002. Resource selection by animals: statistical design and analysis for field studies. 2nd ed. Dordrecht: Springer.
- Manning J, Cronin G, González L, Hall E, Merchant A, Ingram L. 2017. The behavioural responses of beef cattle (Bos taurus) to declining pasture availability and the use of gnss technology to determine grazing preference. Agric. 7(5):1–12.
- Meisser M, Deléglise C, Freléchoux F, Chassot A, Jeangros B, Mosimann E. 2014. Foraging behaviour and occupation pattern of beef cows on a heterogeneous pasture in the Swiss alps. Czech J Anim Sci. 59(2):84–95.
- Muminov A, Sattarov O, Lee CW, Kang HK, Ko MC, Oh R, Ahn J, Oh HJ, Jeon HS. 2019. Reducing GPS error for smart collars based on animal’s behavior. Appl Sci. 9(16):408.
- Myneni RB, Hall FG, Sellers PJ, Marshak AL. 1995. Interpretation of spectral vegetation indexes. IEEE Trans Geosci Remote Sens. 33(2):481–486.
- Nakagawa S, Schielzeth H. 2013. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol. 4(2):133–142.
- Nathan R, Monk CT, Arlinghaus R, Adam T, Alós J, Assaf M, Baktoft H, Beardsworth CE, Bertram MG, Bijleveld AI, et al. 2022. Big-data approaches lead to an increased understanding of the ecology of animal movement. Science. 375(6582):eabg1780.
- Nozière P, Sauvant D, Delaby L. 2017. INRA feeding system for ruminants. Wageningen: Wageningen Academic Publishers.
- Palumbo F, Squartini A, Barcaccia G, Macolino S, Pornaro C, Pindo M, Sturaro E, Ramanzin M. 2021. A multi-kingdom metabarcoding study on cattle grazing Alpine pastures discloses intra-seasonal shifts in plant selection and faecal microbiota. Sci Rep. 11(1):1–14.
- Parraga Aguado MÁ, Sturaro E, Ramanzin M. 2017. Individual activity interacts with climate and habitat features in influencing GPS telemetry performance in an alpine herbivore. Hystrix. 28(1):1–7.
- Pauler CM, Isselstein J, Berard J, Braunbeck T, Schneider MK. 2020. Grazing allometry: anatomy, movement, and foraging behavior of three cattle breeds of different productivity. Front Vet Sci. 7:494–417.
- Payne LX, Schindler DE, Parrish JK, Temple SA. 2005. Quantifying spatial pattern with evenness indices. Ecol Appl. 15(2):507–520.
- Peco B, Navarro E, Carmona CP, Medina NG, Marques MJ. 2017. Effects of grazing abandonment on soil multifunctionality: the role of plant functional traits. Agric Ecosyst Environ. 249:215–225.
- Perotti E, Probo M, Pittarello M, Lonati M, Lombardi G. 2018. A 5-year rotational grazing changes the botanical composition of sub-alpine and alpine grasslands. Appl Veg Sci. 21(4):647–657.
- Pettorelli N, Laurance WF, O’Brien TG, Wegmann M, Nagendra H, Turner W. 2014. Satellite remote sensing for applied ecologists: opportunities and challenges. J Appl Ecol. 51(4):839–848.
- Pettorelli N, Vik JO, Mysterud A, Gaillard JM, Tucker CJ, Stenseth NC. 2005. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol Evol. 20(9):503–510.
- Pittarello M, Probo M, Perotti E, Lonati M, Lombardi G, Ravetto Enri S. 2019. Grazing management plans improve pasture selection by cattle and forage quality in sub-alpine and alpine grasslands. J Mt Sci. 16(9):2126–2135.
- Pittarello M, Ravetto Enri S, Lonati M, Lombardi G. 2021. Slope and distance from buildings are easy-to-retrieve proxies for estimating livestock site-use intensity in alpine summer pastures. PLoS One. 16(11):e0259120.
- Probo M, Lonati M, Pittarello M, Bailey DW, Garbarino M, Gorlier A, Lombardi G. 2014. Implementation of a rotational grazing system with large paddocks changes the distribution of grazing cattle in the south-western Italian Alps. Rangel J. 36(5):445–458.
- Probo M, Massolo A, Lonati M, Bailey DW, Gorlier A, Maurino L, Lombardi G. 2013. Use of mineral mix supplements to modify the grazing patterns by cattle for the restoration of sub-alpine and alpine shrub-encroached grasslands. Rangel J. 35(1):85–93.
- R Core Team. 2016. R: A language and environment for statistical computing. Vienna, Austria: The R Foundation for Statistical Computing.
- Ravetto Enri S, Probo M, Farruggia A, Lanore L, Blanchetete A, Dumont B. 2017. A biodiversity-friendly rotational grazing system enhancing flower-visiting insect assemblages while maintaining animal and grassland productivity. Agric Ecosyst Environ. 241:1–10.
- Rivero MJ, Grau-Campanario P, Mullan S, Held SD, Stokes JE, Lee MR, Cardenas LM. 2021. Factors affecting site use preference of grazing cattle studied from 2000 to 2020 through GPS tracking: A review. Sensors. 21(8):2696.
- Sartorello Y, Pastorino A, Bogliani G, Ghidotti S, Viterbi R, Cerrato C. 2020. The impact of pastoral activities on animal biodiversity in Europe: a systematic review and meta-analysis. J Nat Conserv. 56:125863.
- Schils RLM, Bufe C, Rhymer CM, Francksen RM, Klaus VH, Abdalla M, Milazzo F, Lellei-Kovács E, ten Berge H, Bertora C, et al. 2022. Permanent grasslands in Europe: land use change and intensification decrease their multifunctionality. Agric Ecosyst Environ. 330:107891.
- Scillitani L, Darmon G, Monaco A, Cocca G, Sturaro E, Rossi L, Ramanzin M. 2013. Habitat selection in translocated gregarious ungulate species: an interplay between sociality and ecological requirements. J Wildl Manag. 77(4):761–769.
- Semenzato P, Cagnacci F, Ossi F, Eccel E, Morellet N, Hewison AJM, Sturaro E, Ramanzin M. 2021. Behavioural heat-stress compensation in a cold-adapted ungulate: forage-mediated responses to warming Alpine summers. Ecol Lett. 24(8):1556–1568.
- Shariatinajafabadi M, Wang T, Skidmore AK, Toxopeus AG, Kölzsch A, Nolet BA, Exo KM, Griffin L, Stahl J, Cabot D. 2014. Migratory herbivorous waterfowl track satellite-derived green wave index. PLoS One. 9(9):e108331–11.
- Spedener M, Tofastrud M, Devineau O, Zimmermann B. 2019. Microhabitat selection of free-ranging beef cattle in south-boreal forest. Appl Anim Behav Sci. 213(February):33–39.
- Spiegal S, Estell RE, Cibils AF, James DK, Peinetti HR, Browning DM, Romig KB, Gonzalez AL, Lyons AJ, Bestelmeyer BT. 2019. Seasonal divergence of landscape use by heritage and conventional cattle on desert rangeland. Rangel Ecol Manag. 72(4):590–601.
- Standing Committee on Agriculture (SCA). 1990. Feeding standards for Australian livestock — Ruminants. CSIRO, Australia.
- Sturaro E, Marchiori E, Cocca G, Penasa M, Ramanzin M, Bittante G. 2013. Dairy systems in mountainous areas: farm animal biodiversity, milk production and destination, and land use. Livest. Sci. 158(1–3):157–168.
- Teague WR, Dowhower SL, Baker SA, Haile N, DeLaune PB, Conover DM. 2011. Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric Ecosyst Environ. 141(3–4):310–322.
- Tofastrud M, Devineau O, Zimmermann B. 2019. Habitat selection of free-ranging cattle in productive coniferous forests of south-eastern Norway. For. Ecol. Manage. 437:1–9.
- Tomkiewicz SM, Fuller MR, Kie JG, Bates KK. 2010. Global positioning system and associated technologies in animal behaviour and ecological research. Philos Trans R Soc Lond B Biol Sci. 365(1550):2163–2176.
- Urbano F, Cagnacci F. 2014. Spatial database for GPS Wildlife tracking data. A practical guide to creating a data management system with PostgreSQL/PostGIS and R. 1st ed. Cham: Springer.
- Wood S. 2017. Generalized additive models: an introduction with R. 2nd ed. Boca Raton: Chapman and Hall/CRC.
- Zendri F, Ramanzin M, Bittante G, Sturaro E. 2016. Transhumance of dairy cows to highland summer pastures interacts with breed to influence body condition, milk yield and quality. Ital J Anim Sci. 15(3):481–491.