

Abstract
Diabetes is a consequence of a decrease on functional β-cell mass. We have recently demonstrated that epoxypukalide (Epoxy) is a natural compound with beneficial effects on primary cultures of rat islets. In this study, we extend our previous investigations to test the hypothesis that Epoxy protects β-cells and improves glucose metabolism in STZ-induced diabetic mice.
We used 3-months old male mice that were treated with Epoxy at 200 μg/kg body weight. Glucose intolerance was induced by multiple intraperitoneal low-doses of streptozotocin (STZ) on 5 consecutive days. Glucose homeostasis was evaluated measuring plasma insulin levels and glucose tolerance. Histomorphometry was used to quantify the number of pancreatic β-cells per islet. β-cell proliferation was assessed by BrdU incorporation, and apoptosis by TUNEL staining. Epoxy treatment significantly improved glucose tolerance and plasma insulin levels. These metabolic changes were associated with increased β-cell numbers, as a result of a two-fold increase in β-cell proliferation and a 50% decrease in β-cell death.
Our results demonstrate that Epoxy improves whole-body glucose homeostasis by preventing pancreatic β-cell death due to STZ-induced toxicity in STZ-treated mice.
Abbreviations
| DMSO | = | dimethyl sulfoxide |
| Epoxy | = | Epoxypukalide |
| IPGTT | = | intraperitoneal glucose tolerance test |
| STZ | = | Streptozotocin. |
Introduction
Diabetes is one of the most devastating chronic diseases in the world.Citation1,2 A major hallmark of diabetes is hyperglycemia, a condition related to abnormal insulin secretion by pancreatic β-cells.Citation3 In type 1 diabetes (T1D), β-cell mass decreased by ˜70-100%, whereas in type 2 diabetes (T2D) up to 65%.Citation4-6 In both forms of the disease, pancreatic β-cell apoptosis is augmented by incompletely understood mechanisms that, at least in part, involve β-cell damage and inflammatory infiltrates within pancreas.Citation3-5
In view of the relevance of diabetes disease worldwide, increasing interest has been raised to search for new compounds with therapeutic effects on β-cell abnormal insulin secretion and normalize glycemia in T1D and T2D patients. Thus, intensive research has been focusing on biochemical substances from plant and marine sources with therapeutic potential. Following this line of work, we recently reported that epoxypukalide (Epoxy), a member of the furanocembranolide family isolated from the gorgonian Leptogorgia setacea, exhibited protective effects on pancreatic β-cells ex vivo without altering glucose-stimulated insulin secretion.Citation7 We demonstrated that Epoxy induced β-cell replication in pancreatic β-cell lines (INS1 832/13) and primary rat β-cell cultures through a mechanism involving activation of ERK1/2 signaling pathway and upregulation of cell cycle activators such as cyclin D2 and cyclin E. In addition, Epoxy showed protection against basal and cytokine-mediated β-cell death in primary cultures of rat islets.Citation7 These results spurred us to spread the notion that Epoxy may be translated into a new treatment for diabetes prevention.
Experimental animal models of diabetes exhibit β-cell abnormal insulin secretion and elevated blood glucose levels, which are associated with oxidative stress and inflammation. Streptozotocin (STZ) is a natural compound with toxic effects on β-cells, currently used in single large doses or multiple low doses to induce diabetes in mice. Thus, the STZ-induced diabetic mouse model is widely used for the screening of new compounds with protective properties against β-cell death and hypoglycemic effects.Citation8,9
This work was designed to test the hypothesis that in vivo administration of Epoxy exerts protective effects on pancreatic β-cells and glucose homeostasis in a STZ-induced diabetic mouse model.
Methods
Animals
Male Swiss OF1 (Clr:OF1), were used for the study. Mice were 3 months-old of age at the beginning of the study. They were bred and housed in the Animal Facilities of the School of Medicine, University of Valladolid (UVa), Spain. Mice were fed standard rodent chow and water ad libitum in ventilation-controlled cages in a 12-hour light/dark cycle. Experimental procedures were approved by the Animal Care and Use Committee of the UVa in accordance with the European and Spanish Guidelines for the Care and Use of Mammals in Research.
Study design
Mice were randomly assigned into 2 groups: “Control” (vehicle = 0.001% DMSO) (N = 8) and “Epoxy” (200 µg epoxypukalide/kg body weight) (N = 10). Both groups received daily IP (intraperitoneal) injections of vehicle or Epoxy for 30 days before they were treated with multiple low dose-STZ to study if Epoxy was having a protective effect on pancreatic β-cells. IP injections of freshly prepared STZ at dose of 50 mg/kg body weight (in 0.01 M citrate buffer, pH 4.5) for 5 consecutive days.
Metabolic characterization was performed when glucose intolerance was identified. Mice were IP injected with BrdU (50 mg/kg) 16 hours before been euthanized.
Plasma biochemistry
Blood samples were obtained from mice under non-fasting conditions. Blood was collected from the tail vein into capillary tubes precoated with potassium-EDTA (Sarstedt, Germany) for the preparation of plasma. Non-fasting plasma insulin levels were measured using ultrasensitive mouse ELISA assay according to manufacturer’s instructions (Mercodia, Sweeden).
Glucose tolerance test
IP-GTT (intraperitoneal glucose tolerance test) was performed as previously described.Citation10 Briefly, mice were fasted for 16 hours and injected intraperitoneally with glucose at 2 g/kg of body weight. Blood glucose levels were determined using a Glucometer Xceed (Abbott Diabetes Care, UK). Blood glucose levels were determined at indicated time points and plotted as a function of time.
Histological examinations determination of islet histomorphometry
Mice were euthanized and histopathological pancreata samples were excised and fixed in Bouin’s solution for 4 h, washed in water and then fixed in 10% neutral buffered formalin. Fixed tissue samples were processed and paraffin embedded, sectioned and stained.
For islet histomorphometry sections were stained with anti-insulin antibody (Abcam, UK). The number of islets and the number of pancreatic β-cells of each islet were quantified, and quantitative islet histomorphometry was performed as previously described using ImageJ software (NIH, USA).Citation11
β-cell proliferation was detected by BrdU incorporation and double detection of BrdU/insulin (Anti-BrdU, Abcam, UK), a minimum of 1,000 β-cells were counted per pancreas. β-cell death was detected by double staining of TUNEL (Promega, USA) and insulin, a minimum of 1,000 β-cells were counted per pancreas, as previously described (3, 17).
Statistical analysis
Statistical analysis of data was performed using the Graph Pad Prism4 (GraphPad Software, Inc., La Jolla, CA, USA). Distribution of variables was checked using the Kolmogorov-Smirnov test. When a variable was distributed normally, data were presented as mean ± SD. Comparisons between 2 groups were done by unpaired Student´s t-test, and comparisons between more than 2 groups were performed by ANOVA followed by Tukey's Multiple Comparison Test. Differences were considered significant at P < 0.05.
Results
Effects of epoxypukalide treatment on pancreatic β-cells
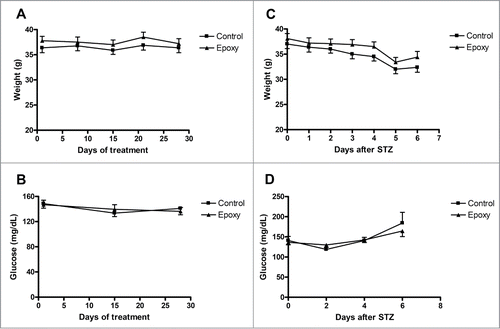
In this study, we have used a STZ-diabetic mouse model to investigate the effects of Epoxy on glucose metabolism. STZ is an alkylating agent that causes DNA damage in pancreatic β-cells leading to β-cell damage and lower insulin production.Citation8,12,Citation13 First, we determined if Epoxypukalide had an impact on body weight or glucose before STZ treatment. Therefore, we monitored body weight once a week and non-fasting blood glucose every other week. Body weight and glucose levels were similar in both Control and Epoxy groups during the duration of the treatment and before STZ treatment (). Epoxy injections did not have a harmful effect on mice, they looked perfectly healthy, and they ate, drank and behaved normally. Furthermore, we tracked body weight daily and non-fasting glucose every other day for a week after STZ treatment, observing no-differences between both groups ().
Figure 1. Body weight and non-fasting blood glucose tracking before and after STZ treatment. (A) Body weight was measured every week and blood glucose was measured every other week before STZ treatment in both groups, vehicle-treated (Control) and Epoxy-treated (Epoxy). (B) Body weight was measured every day and blood glucose was measured every other day after STZ treatment in both groups, Control and Epoxy. No significant differences were detected.
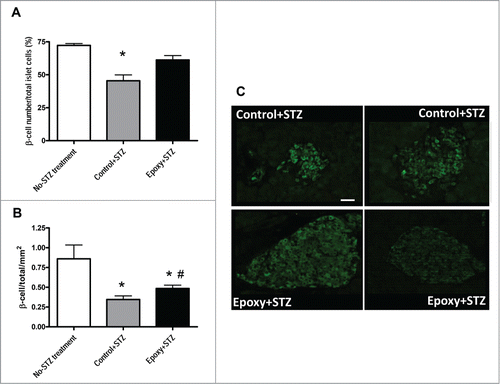
To evaluate the impact of Epoxy on pancreatic β-cells treated with STZ, we determined the number of islets and the number of pancreatic β-cells of each islet in Control and Epoxy treated mice. We used a group of no-STZ treated mice as a control of STZ effect on β-cell area. Islet numbers were similar in Epoxy+STZ versus Control+STZ pancreata (0.19 ± 0.02 islets/ mm2 vs. 0.18 ± 0.02 islets/ mm2) but they were significantly different to no-STZ treated pancreata (0.46 ± 0.04 islets/mm2). Interestingly, STZ treatment reduced β-cell number by half; this reduction was partially prevented by Epoxy since there was a significant 30% increase in β-cell number in Epoxy+STZ versus Control+STZ pancreata ().
Figure 2. Quantitative islet histomorphometry in mice treated with epoxypukalide. Histomorphometry on pancreata sections was performed in mice without STZ or Epoxy treatment (no-STZ treated mice), in mice treated with STZ plus vehicle (Control+STZ)and mice treated with STZ plus Epoxy (Epoxy+STZ). (A) Average β-cell number per islet. (B) Average β-cell number per islet per pancreas area (mm2). (C) Representative insulin-stained islets from Control+STZ and Epoxy+STZ treated mice (scale bar = 50 µm). N = 8-10 mice per group. *P < 0.05 relative to no-STZ treatment group; #P < 0.05 relative to Control+STZ group.
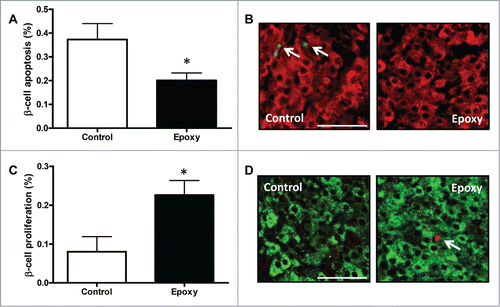
β-cell mass can be increased by augmented proliferation and/or decreased β-cell death. To understand how β-cell number was increased in Epoxy treated mice we quantified both. β-cell death was detected by TUNEL staining, and proliferation by measuring BrdU incorporation in insulin-positive cells. Our results showed a 50% decrease in β-cell death in Epoxy treated β-cells (), and a 2-fold increase in β-cell proliferation (). Interestingly, this proliferative effect is specific on pancreatic β-cells, since other endocrine cells as α-cells showed similar proliferation in both conditions (0.04 ± 0.03 % vs. 0.09 ± 0.05 %; p = 0.49). These results support the view that Epoxy is an inhibitor of β-cell apoptosis under STZ toxicity, and it enhances β-cell proliferation in vivo.
Figure 3. β-cell death and proliferation in mice treated with epoxypukalide. (A) Quantification of β-cell apoptosis (measured by TUNEL) in Control and Epoxy treated STZ-diabetic mice (N = 7-10). (B) Representative pictures of insulin and TUNEL staining. (C) Quantification of β-cell proliferation (measured by BrdU incorporation) in Control and Epoxy treated STZ-diabetic mice (N = 6-10). (D) Representative pictures of insulin and BrdU staining (scale bar = 50 µm). *P < 0.05 relative to control group.
Effects of epoxypukalide treatment on glucose homeostasis
To investigate whether in vivo β-cell expansion was having an impact on glucose homeostasis, we measured non-fasting plasma insulin levels and glucose tolerance in STZ-mice treated with vehicle or Epoxy. As shown in , non-fasting plasma insulin levels exhibited a 2-fold augment in Epoxy treated mice compared to Control mice. These results nicely correlated to β-cell expansion data shown in .
Figure 4. Effect of epoxypukalide treatment on glucose metabolism. (A) Non-fasting plasma insulin was measured in Control and Epoxy treated STZ-diabetic mice. N = 8-10 mice per group. (B) Glucose tolerance test was measured in Control and Epoxy treated STZ-diabetic mice, non-diabetic mice were used to establish the levels of “normal glucose tolerance.” N = 6-7 mice per group. (C) Area under the curve. *P < 0.05 measured by ANOVA.
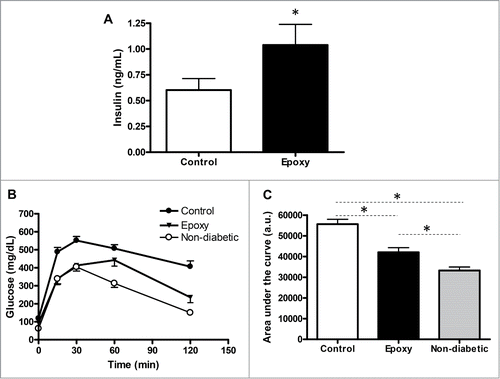
On the other hand, glucose tolerance measured by IP-GTT was significantly improved in Epoxy-treated mice compared to control mice (). Non-diabetic mice were used to establish the levels of normal glucose tolerance. These data are in good agreement with those observed for plasma insulin levels in Epoxy-treated mice. Taken together, these results suggest that Epoxy is a potential treatment for β-cell preservation and diabetes prevention.
Discussion
Epoxypukalide (Epoxy) is a natural compound firstly isolated by Ksebati et al from the gorgonian Leptogorgia setacea.Citation14 Since then, different groups have identified and purified Epoxy and other members of the furanocembranolides family. In 2007, Dorta et al. reported their isolation from specimens from the genus Leptogorgia collected around the Isthmus of Panama;Citation15 and Grote et al. also isolated Epoxy from the soft coral Litophyton arboretum.Citation16 Although Epoxy has not yet been obtained by synthesis, furanocembranolides are also a popular target for total synthesis.Citation17,18
In this manuscript, we have shown for the first time that Epoxy treatment prevents β-cell death and induces β-cell proliferation in a preclinical diabetes model. Both events led to increased β-cell number and augmented insulinemia, which allowed a better glycemic control. These results are nicely supported by our recent manuscript reporting that Epoxy is an inductor of β-cell proliferation ex vivo, and that it protects β-cells from basal and cytokine-induced apoptosis.Citation7
Of note, Epoxy does not induce proliferation in non-β-cell proliferation in vivo, similar results have been observed in our laboratory in primary cultures of rat pancreatic cells (unpublished data). These data point out to a specific effect of Epoxy on the pancreatic β-cell.
Most important, increased β-cell proliferation may involve loss of differentiation and cell function failure, but despite its positive effect on proliferation, Epoxy did not show a deleterious effect on insulin production and secretion in Epoxy-treated β-cells ex vivo.Citation7 In the same way, now we have showed that Epoxy treatment in STZ-diabetic mice did not exert negative effects on pancreatic β-cell function, in fact, increased β-cell mass produced higher plasma insulin levels improving glucose control.
Interestingly, pancreatic β-cells are highly susceptible to oxidative stress because of low antioxidant enzyme activity levels.Citation19 We previously showed that Epoxy protected primary rat β-cell cultures from a cocktail of pro-inflammatory cytokines including IL-1β, IFN-γ and TNF-α.Citation7 Herein, we have showed that Epoxy is protecting β-cell death from detrimental actions of local inflammation on STZ pancreata. The diabetogenic action of STZ is mediated mostly by reactive oxygen species (ROS).Citation9 The presence of the furan ring of Epoxy could decrease oxidative stress by quenching damaging ROS.Citation15 Taken together, these results indicate that Epoxy protects β-cells from cytokine-mediated oxidative stress. We are currently working on obtaining Epoxy related compounds to examine further their therapeutic potential for diabetes prevention and/or treatment.
In conclusion, our current data demonstrate that Epoxy can significantly prevent experimental T1D in STZ-diabetic mice. In addition, we provide evidences that its positive effects on glucose metabolism are mediated by improved pancreatic β-cell mass due to decreased cell death and increased proliferation.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
FUNDING
This work was supported by grants from: Programa Ramón y Cajal (RYC-2011-08101) Ministerio de Economía y Competitividad (Spain); Sociedad Española de Diabetes Basic Research Grant (2014) to IC.
References
- Horton ES. NIDDM–the devastating disease. Diabetes Res Clin Pract 1995; 28(Suppl):S3-11; PMID:8529517; http://dx.doi.org/10.1016/0168-8227(95)01087-T
- Wild S, Roglic G, Green A, Sicree R, King H. Global prevalence of diabetes: estimates for the year 2000 and projections for 2030. Diabetes Care 2004; 27(5):1047-53; PMID:15111519; http://dx.doi.org/10.2337/diacare.27.5.1047
- Lontchi-Yimagou E, Sobngwi E, Matsha TE, Kengne AP. Diabetes mellitus and inflammation. Curr Diab Rep 13(3):435-44; PMID:23494755; http://dx.doi.org/10.1007/s11892-013-0375-y
- Butler AE, Janson J, Bonner-Weir S, Ritzel R, Rizza RA, Butler PC. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 2003; 52(1):102-10; PMID:12502499; http://dx.doi.org/10.2337/diabetes.52.1.102
- Matveyenko AV, Butler PC. Relationship between beta-cell mass and diabetes onset. Diabetes Obes Metab 2008; 10(Suppl 4):23-31; PMID:18834430; http://dx.doi.org/10.1111/j.1463-1326.2008.00939.x
- Weir GC, Bonner-Weir S. Islet beta cell mass in diabetes and how it relates to function, birth, and death. Ann N Y Acad Sci 2013; 1281:92-105; PMID:23363033; http://dx.doi.org/10.1111/nyas.12031
- Lopez-Acosta JF, Moreno-Amador JL, Jimenez-Palomares M, Diaz-Marrero AR, Cueto M, Perdomo G, Cozar-Castellano I. Epoxypukalide induces proliferation and protects against cytokine-mediated apoptosis in primary cultures of pancreatic beta-cells. PLoS One 2013; 8(1):e52862; PMID:23300997; http://dx.doi.org/10.1371/journal.pone.0052862
- Like AA, Rossini AA. Streptozotocin-induced pancreatic insulitis: new model of diabetes mellitus. Science 1976; 193(4251):415-7; PMID:180605; http://dx.doi.org/10.1126/science.180605
- Lei YC, Hwang JS, Chan CC, Lee CT, Cheng TJ. Enhanced oxidative stress and endothelial dysfunction in streptozotocin-diabetic rats exposed to fine particles. Environ Res 2005; 99(3):335-43; PMID:16307975; http://dx.doi.org/10.1016/j.envres.2005.03.011
- Jimenez-Palomares M, Ramos-Rodriguez JJ, Lopez-Acosta JF, Pacheco-Herrero M, Lechuga-Sancho AM, Perdomo G, Garcia-Alloza M, Cozar-Castellano I. Increased Abeta production prompts the onset of glucose intolerance and insulin resistance. Am J Physiol Endocrinol Metab 2012; 302(11):E1373-80; PMID:22414803; http://dx.doi.org/10.1152/ajpendo.00500.2011
- Cozar-Castellano I, Haught M, Stewart AF. The cell cycle inhibitory protein p21cip is not essential for maintaining beta-cell cycle arrest or beta-cell function in vivo. Diabetes 2006; 55(12):3271-8; PMID:17130470; http://dx.doi.org/10.2337/db06-0627
- Nukatsuka M, Yoshimura Y, Nishida M, Kawada J. Importance of the concentration of ATP in rat pancreatic beta cells in the mechanism of streptozotocin-induced cytotoxicity. J Endocrinol 1990; 127(1):161-5; PMID:2151740; http://dx.doi.org/10.1677/joe.0.1270161
- Reddy S, Wu D, Elliott RB. Low dose streptozotocin causes diabetes in severe combined immunodeficient (SCID) mice without immune cell infiltration of the pancreatic islets. Autoimmunity 1995; 20(2): 83-92; PMID:7578872; http://dx.doi.org/10.3109/08916939509001931
- Ksebati MB, Ciereszko LS, Schmitz FJ. 11 beta, 12 beta-Epoxypukalide, a furanocembranolide from the gorgonian Leptogorgia setacea. J Nat Prod 1984; 47(6):1009-12; PMID:6152470; http://dx.doi.org/10.1021/np50036a018
- Dorta E, Diaz-Marrero AR, Brito I, Cueto M, D'Croz L, Darias J. The oxidation profile at C-18 of furanocembranolides may provide a taxonomical marker for several genera of octocorals. Tetrahedron 2007; 63(37):9057-62; http://dx.doi.org/10.1016/j.tet.2007.06.080
- Grote D, Dahse HM, Seifert K. Furanocembranoids from the soft corals Sinularia asterolobata and Litophyton arboreum. Chem Biodivers 2008; 5(11):2449-56; PMID:19035574; http://dx.doi.org/10.1002/cbdv.200890210
- Roethle PA, Trauner D. The chemistry of marine furanocembranoids, pseudopteranes, gersolanes, and related natural products. Nat Prod Rep 2008; 25(2):298-317; PMID:18389139; http://dx.doi.org/10.1039/b705660p
- Toelle N, Weinstabl H, Gaich T, Gaich T, Mulzer J. Light-mediated total synthesis of 17-deoxyprovidencin. Angew Che Int Ed 2014; (53):3859-62; PMID: 24604885; http://dx.doi.org/10.1002/anie.201400617
- Tiedge M, Lortz S, Drinkgern J, Lenzen S. Relation between antioxidant enzyme gene expression and antioxidative defense status of insulin-producing cells. Diabetes 1997; 46(11):1733-42; PMID:9356019; http://dx.doi.org/10.2337/diab.46.11.1733

