

ABSTRACT
Background: Ginsenoside Rg3 has been proposed to mediate anti-diabetic effects, but their direct effect on pancreatic β cell viability and mechanisms are not clearly understood. Recent studies suggest that intermittent high glucose (IHG) could be more harmful to pancreatic β cells than sustained high glucose. There are few reports about the effect of the ginsenosideRg3 to β cell apoptosis and proliferation against IHG.
Methods: INS-1 cells were treated with alternative glucose concentration with or without ginsenoside Rg3. Cell apoptosis and viability were detected by Annexin V staining and MTT assay. The activation of mitogen-activated protein kinases (MAPKs) was analyzed by Western blotting using specific antibodies. Quantification of secreted insulin protein was measured using rat/mouse Insulin ELISA kits. Bromodeoxyuridine (BrdU) staining and florescence in situ hybridization (FISH) analysis was performed to compare cell proliferation.
Result: INS-1 cell viability was decreased under IHG and increased with Rg3 treatment.Rg3 significantly reduced the apoptotic INS-1 cells against IHG. The quantification of secreted insulin concentration was increased with Rg3. Rg3 increased INS-1 cell proliferation. ERK and p38 MAPK pathways reduced by IHG were activated by the ginsenoside Rg3.
Conclusion: Ginsenoside Rg3 protected INS-1 cell death from IHG with reducing apoptosis and increasing proliferation.
Introduction
Ginseng has a long history as a traditional medicine for various health purposes. It is one of the most popular herbs in East Asia,Citation1 and is now used worldwide as a natural medicine. Ginseng is known to have many pharmacological effects, including immune, cardiovascular, central nervous systems and metabolism.Citation2,3 Ginseng is also proposed to mediate anti-diabetic effects by stimulating insulin secretion, stimulating glucose uptake, inhibiting intestinal absorption of glucose, and reducing glycogenolysis.Citation4-6 Its mechanisms are observed in various organs, including liver, muscle, adipose tissue, intestine and pancreas. There are few studies about the direct effects of ginseng on anti-apoptosis and proliferation of pancreatic β cells. The active components of ginseng are ginsenosides, a group of steroidal saponins. Ginsenoside Rg3 is one of the most active component of Panax ginseng. It is known that ginsenoside Rg3 has a protective effect against hyperglycemia, obesity and diabetes in vivo.Citation7,8 Rg3 is also considered as one of the effective components on insulin secretionCitation7 and glucose uptake with different molecular targets in vitro.Citation6 Compared with other ginsenoside, Rg3 has reported a little more with various cell lines.Citation6
Several recent studies suggest a relationship between glucose fluctuations and the development of diabetic vascular complications, especially in type 1 diabetes.Citation9 In type 2 diabetes, HbA1c variability was associated with all-cause mortality.Citation10 Glucose variability exhibited a more pronounced effect on oxidative stress than sustained hyperglycemia.Citation11 We previously reported that a more harmful effect exists with intermittent high glucose concentrations than with sustained high glucose concentrations on pancreatic β cells and that was related to forkhead transcription factor (FOXO).Citation12,19
Oxidative stress induces increased production of free radicals and it can activate members of mitogen-activated protein kinase (MAPK) cascades, including JNK, ERK, and p38 MAPK. These proteins mediate many important signals in mammal cells, including cell proliferation, differentiation, and apoptosis.Citation13,14 In terms of cell survival or apoptosis, activation of MAPK pathways may be different depending on the stimuli and cell type.Citation15 Studies report ginseng can regulate cell survival through MAPK signaling pathways.Citation16,17 The relationship between the direct effect of ginseng saponins (specifically Rg3) on pancreatic β cells and MAPK signaling under intermittent hyperglycemia has not yet been clarified. We therefore investigated whether Rg3 has beneficial effects on pancreatic β cell function and viability against intermittent hyperglycemic stress, and whether this effect is regulated via ERK and p38 MAPK signaling.
Methods
Cell culture and intermittent high glucose treatment
INS-1 rat insulinoma cell lines were obtained from Yeung-Nam University (Daegu, Korea). INS-1 cells were maintained in RPMI 1640 medium containing 10% fetal bovine serum, 10 mM HEPES, 11 mM glucose, and 50μM2-mercaptoethanol. Rg3 was purchased from Sigma-Aldrich (MO, USA). All experiments were incubated at 37°C with 5% CO2 and studied between the 30th and 40th passage.INS-1 cells were plated in 6-well plates at 5 × 104 cells per well and incubated for 48 h.
Cells were exposed to constant 11 mM glucose (Control). We changed media every 12 h for 48 h. Intermittent high glucose condition was same as our previous studies.Citation11,18 Briefly, INS-1 cells were exposed to intermittent high glucose (alternating glucose concentration of 11 mM and 33 mM glucose every 12 h for 48 h) with/without Rg3 (1, 5, 10μM),.
Annexin V staining for apoptosis detection
INS-1 cells were plated in 6-well plates at 1 × 106 cells per well and incubated under appropriate conditions. To make single cell staining, cells were treated with trypsin-EDTA and centrifuged at 1200 rpm for 3 min at 4°C. After aspirating supernatants, cells were washed with 1 mL of Annexin V binding solution (140 mMNaCl, 10 mM HEPES pH 7.4, 2.5 mM CaCl2) and centrifuged at 1200 rpm for 3 min at 4°C. Supernatants were removed and 3 uL of Annexin V-FITC and 10 uLofPropidium Iodide were added. After incubation for 15 min in the dark, 300 uL of FACS buffer (1% FBS, 0.1% NaN3) was added and analyzed by FACSort (Becton Dickenson, BD Bioscience).
Cell viability
INS-1 cells were seeded in 48-well plates at 5 × 104 cells per well and incubated for 48 h. Cell viability was determined by MTT assay. The optical density was measured at 450 nm by an automatic plate reader (Seoul in Bioscience, Gyeonggi-do, Korea).
Western blotting
INS-1 cells were washed with PBS and lysed in mammalian tissue lysis/extraction reagent including protease inhibitor(Roche, CA, USA). A BCA protein assay kit was used to quantify proteins and samples were mixed with 1× SDS sample buffer (50 mM Tris pH6.8, 2% SDS, 10% glycerol, 50 mM DTT, and 0.01% bromophenol blue). Proteins were separated in 12% SDS-PAGE, transferred onto PVDF membrane, and immunoblotted with anti-p38 MAPK (1:1000,Cell Signaling, MA, USA), anti-phospho p38 MAPkinase (Thr180/Tyr182) (1:1000,Cell Signaling, MA, USA), anti-p44/42 MAPK (Erk1/2) (1:1000,Cell Signaling, MA, USA), anti-phospho p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (1:1000,Cell Signaling, MA, USA), anti-caspase 3 (1:1000,Cell Signaling, MA, USA) and anti-cleaved caspase 3 (1:1000,Cell Signaling, MA, USA) at 4°C overnight. Secondary antibodies (goat anti-rabbit conjugated alkaline phosphase, Cell Signaling, MA, USA) were applied for 1 h at room temperature and membranes were developed via AP-conjugated development kit (Bio-rad, CA, USA). Developed protein bands were quantified by Multi Gauge V2.2 program (Tokyo, Japan).
Quantification of secreted insulin protein by ELISA
INS-1 cells were plated in 6-well plates at 5 × 104 cells per well and incubated for 48 h. After incubation in normal or high glucose concentrations and/orRg3, cells were washed with PBS. To test insulin secretion on glucose-stimulated insulin secretion (GSIS), cells were starved for 5 h in RPMI medium containing 3 mM glucose and 2% FBS. Cell media was then changed to KRBB solution (4.74 mM KCl, 1.19 mM KH2PO4, 1.19 mM MgCl2·6 H2O, 35 mM NaHCO3, 10 mM HEPES) containing 3 mM glucose or 15 mM glucose and cells were incubated for 1 h. Secreted insulin protein in the media was measured using rat/mouse insulin ELISA kit (Linco Research, MO, USA).
BrdU staining
Bromodeoxyuridine (BrdU) was added in cell cultures to a final concentration of 10 μM BrdU. The cells were incubated for an additional 16 h at 37°C with 5% CO2. After supernatants were aspirated, cells were fixed and permeabilized with cytoperm/cytofix (BD Biosciences, CA, USA) for 25 min and washed with perm/wash buffer (0.1% saponin, 1 mM CaCl2•2 H2O, 1 mM MgSO4, 0.05% NaN3, 10 mM HEPES, and 1% BSA). For DNA denaturation, the cells were incubated with 2 M HCl and 0.5% Tween20 in H2O. The cells were incubated at room temperature for 20 min, then washed with 100μL of 10% 2 M NaOH / 90% Hank's Balanced Salt Solution (HBSS), twice with 1 mL of HBSS, and once with 0.1% Tween 20, 0.5% BSA. Then 100 mL of 0.5 μg/mL mouse anti-BrdU FITC-conjugated antibody was added and cells were incubated for 1 h at room temperature, and finally washed twice with 1 mL PBS. Cells were then stained with diluted DAPI (10 μg/mL) for 20 min and washed 3 times in 1X PBS for 5 min each. The cells were dropped on slide glass and cover slipped with Prolong® Gold Antifade Reagent (In vitro gen Life Technologies, NY, USA). FISH analysis was performed using a green fluorescence microscope, Olympus BX-51 (Olympus, Tokyo, Japan).
Statistical analysis
All the experiments were repeated 3 times and values are presented as Means ± SD. Data were analyzed using a one way ANOVA, followed by Turkey's post hoc test with SPSS 19.0 statistical software. A value of p<0.05 was considered significant and p < 0.01 highly significant.
Results
Ginsenoside Rg3 increased INS cell viability and reduced apoptosis against intermittent high glucose
After exposure to intermittent high glucose for 48 h, INS-1 cell viability decreased by 38% compared to controls. Rg3 treatment significantly increased INS-1 cell viability under IHG (p < 0.05). Rg3 at 5μM showed significantly higher cell viability than Rg3 at 1μM in IHG stress (p < 0.05). Cell viability under IHG with treatment of Rg3 10μM was better than Rg3 1μM and 5μM, yet there was no significant difference (Rg3 5μM (76%) vs Rg3 10μM (81%), p = 0.16, )
Figure 1. Effect of ginsenoside Rg3 on INS-1 cell viability and apoptosis in intermittent high glucose. The cells were treated with normal and high (33 mM) glucose alternating 12 h with Rg3 (1, 5, 10μM) for 2 d. (A) Cell viability was determined by MTT assay. (B) Cell was stained by Annexin V for apoptosis detection and analyzed by FACSort. Values are means ± SD (n = 3). Means with different marks differ significantly among groups (p < 0.05). C means control group. *p < 0.01 relative to control; **p < 0.05 relative to IHG treated without Rg3 group.
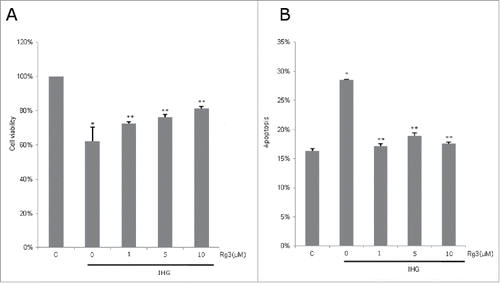
Ginsenoside Rg3 attenuated intermittent high glucose-induced apoptosis
Intermittent high glucose significantly increased INS-1 cell apoptosis compared to control (p < 0.05). The proportion of apoptotic INS-1 cells under IHG was significantly reduced with Rg3 application. Anti-apoptotic effects of Rg3 were not dose-dependent, showing similar results among Rg3 1μM, 5μM and 10μM treatments (). To determine IHG induced apoptotic cell death, cleavedcaspase-3 was also measured by Western blotting. The cleavage fragment of caspase-3 was higher in cells exposed to IHG and decreased with ginsenoside Rg3 (). The proportion of apoptotic cells in the Rg-treated group in immunoblotting (relative cleaved caspase 3 / α-tubulin) showed similar results among Rg3 1μM, 5μM and 10μM concentrations ().
Figure 2. Effect of ginsenoside Rg3 on expression of caspase 3, p38 MAPK, ERK activation in intermittent high glucose-treated INS-1 cells. The cells were treated with IHG in the presence or absence of Rg3 for 48 h and harvested for whole-cell lysates to be used in Western blots. Protein bands were quantified and the ratio of phosphorylated forms of (A) caspase 3 (B) p38 MAPK (C) ERK were presented by graph. All data are representative of 3 independent experiments. Means with different marks differ significantly among groups (p < 0.05). C means control group. #p < 0.05 relative to control; *p < 0.05 relative to IHG treated without Rg3 group.
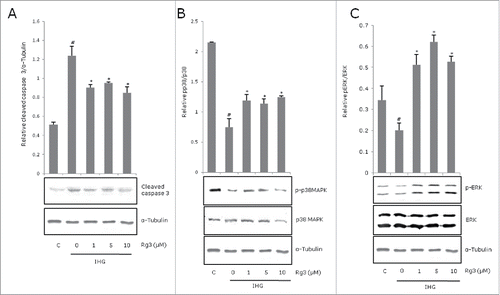
Ginsenoside Rg3 suppressed intermittent high glucose-induced apoptosis by activation of ERK and p38 with phosphorylation
To examine the involvement of MAPKs in INS-1 cell apoptosis, the phosphorylated forms of ERK and P38 were measured by Western blotting with specific antibodies to determine the activation status of these MAPKs. IHG suppressed phosphorylated forms ofp38 MAPK and ERK. (). Rg3 treatment significantly increased phosphorylation of p38 MAPK and ERK, but did not show a dose-dependent effect.
Ginsenoside Rg3 enhanced insulin secretion and β cell proliferation
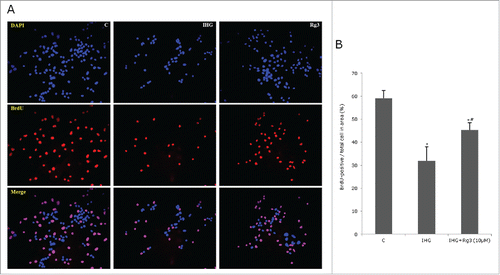
We examined the effect of Rg3 on insulin secretion and INS-1 cell proliferation. Glucose stimulated insulin secretion (GSIS) was decreased in IHG compared to control. After co-culturing with Rg3(10μM), GSIS was increased 47% in 15 mM glucose solution ().We evaluated cell proliferation with bromodeoxyuridine (BrdU), and used DAPI as a counter stain to visualize cell nuclei. The percentage of BrdU-positive cells / total cells in area were 35% in IHG condition and 45.3% in IHG + Rg3 condition (p < 0.05) (). When the images were merged, the majority of the cells labeled with DAPI (Blue) presented BrdU-positive (Red, ) in cells treated with Rg3 compared to IHG.
Figure 3. Effect of ginsenoside Rg3 on Glucose stimulated insulin secretion. The cells were starved and incubated with 3 mM glucose and 5 mM glucose containing medium for 1 h. Secreted insulin protein in the media was measured using Rat/mouse Insulin ELISA kit. All data are representative of 3 independent experiments. Means with different marks differ significantly among groups (p < 0.05). #P < 0.05 relative to control; *p < 0.05 relative to control; **p < 0.05 relative to IHG treated without Rg3 group.
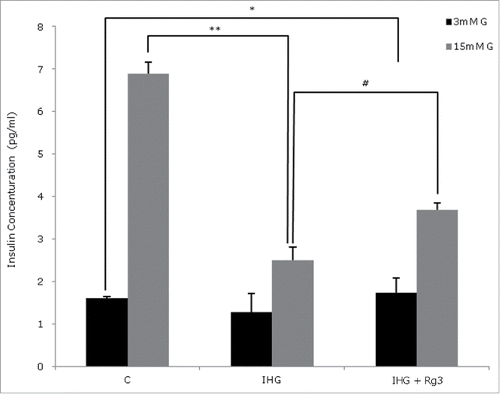
Figure 4. Effect of ginsenoside Rg3 on INS-1 cell proliferation. The effects on INS-cell proliferations were assessed by BrdU and DAPI staining. Control, IHG, Rg3 (10μM) was compared. (A) Quantification of INS-cell proliferations were assessed by BrdU-positive / total cell in area (%). (B) All data are representative of 3 independent experiments. C means control group. *P < 0.05 relative to control; #p < 0.05 relative to IHG treated without Rg3 group.
Discussion
Our present study questioned whether the traditional herb ginseng has direct beneficial effects on pancreatic β cell viability. We found that ginsenoside Rg3 protected against INS-1 cell apoptosis induced by intermittent high glucose (IHG) stress. Rg3 increased cell viability, proliferation and glucose induced insulin secretion. One possible mechanism is Rg3 may activate p38 MAPK and ERK phosphorylation against IHG.
Recent epidemiologic studies reported that glucose fluctuation or HbA1c variability are related to diabetic complications including cardiovascular events and diabetic nephropathy.Citation9,11 Impairment of pancreatic β cell functions are induced by chronic sustained high glucose (SHG), but a recent study showed that cellular ER and oxidative stress were significantly increased under conditions of IHG.Citation18 Our previous studies also showed that IHG induced more INS-1 cell apoptosis than SHG and it was related to higher oxidative stress.Citation12,19 The protective effects of ginsenosides on pancreatic β cells against intermittent hyperglycemia have not been studied. Our present study showed that the ginsenoside Rg3 attenuated pancreatic β cell apoptosis and improved insulin secretion under IHG stress.
Several mechanisms of anti-diabetic effects of ginseng have been suggested. They are categorized depending on organs. In liver, ginsenosides can inhibit glucose production. Rg1, another kind of ginsenoside, suppresses hepatic glucose production in HepG2 human hepatoma cells, and the effect was associated with the LKB1-AMPK-FoxO1 pathway.Citation20 Ginsenosides increase insulin-mediated glucose uptake in myotubes and adipocytes.Citation4,5 Most studies about the effects of ginsenosides on the pancreas were related to insulin secretion. Rg3 could enhance insulin secretion through the activation of ATP sensitive K+ channels and AMPK signaling pathways and stimulated elevation of intracellular calcium concentrations.Citation8,21 There are some studies about the anti-apoptotic effects of ginseng on pancreatic β cells. They attenuated pancreatic β cell death against free fatty acids, cytokines, palmitate and sustained high glucose.Citation5,22,23 To date, studies about protective effects of ginseng against intermittent high glucose are rare. In this study, we showed that intermittent high glucose increased apoptosis and decreased insulin secretion in INS-1 cells. The ginsenoside Rg3, especially in a high dose (10μM), attenuated cell apoptosis and improved cell viability in INS-1 cells under IHG.
The mechanism for the direct effects of ginsenosides on pancreatic β cell viability has not been fully evaluated. Luo JZ,Citation23 wrote that American ginseng may regulate mitochondrial UCP-2 and ATP levels, and thereby increase insulin synthesis and anti-apoptotic cascades in pancreatic β cells. High glucose/cytokine induced β cell loss by increasing NO production and apoptosis-related gene expressions were also reported. Ginsenosides reduced iNOS gene expression, and NO production may lead to pancreatic β cell protection from high glucose/cytokine interactions.Citation24 Cleaved caspase 3 as an apoptotic signal was increased in IHG and decreased with Rg3 treatments in our study, which could signify thatRg3 decreased β cell apoptosis against IHG.
We further investigated whether the effects of ginsenosides on pancreatic β cell viability under IHG was related to the MAPK/ERK pathway. Mitogen activated protein kinase (MAPK) is known to be involved in cell survival, apoptosis, proliferation, and differentiation in mammalian cells, including pancreatic β cells. The targeted selection of downstream substrates depends upon the cell type and nature of the stimuli that specify differentiation.Citation15 MAPK signaling pathways are reported to be involved in pancreatic β cell impairment. P38 MAPK and ERK are activated by interleukin-1 β and an inhibition of these signals decreased cytokine-induced rat islet cell apoptosis.Citation17 When pancreatic β cells we reexposed to both hyperglycemia and palmitic acid, p38may be involved in increasing the phosphorylated forms of JNK and ERK1/2.Citation16 There are several studies showing the role of ERK1/2 as survival signals, but the role of p38 during pancreatic β cell apoptosis has remained controversial. Lingli Zhou et al,Citation25 suggested the inhibition of p38 prevents glucolipotoxicity in β cells. The relationships between ginsenosides and p38 and ERK signaling are also not consistent. Rb1 increased phosphorylation levels of ERK1/2 in MPP(+)-treated PC12 cells, but it reduced phosphorylated p38.Citation26 Another ginsenoside, AD-1, up-regulates the expression of p38 and ERK phosphorylation in cancer cells, which induced cancer cell death.Citation27 Other studies reported a cell proliferating effect of ginsenoisdes, which was related to p38 and ERK signal activation.Citation28,29 Our study showed that Rg3 increased proliferation marker BrdU staining in INS-1 cells under IHG. ERK and p38 MAPK phosphorylations were decreased significantly in IHG but increased with Rg3 treatment. Because previous studies showed inconsistent effect of ginsenosides on p38 and ERK phosphorylation depending on cell type and stimuli, our results may be interpreted asRg3 proliferates INS-1 cells against IHG through ERK and p38 MPPK activation.
There were some limitations in this study. First, we used INS-1 cell for evaluating pancreas β-cell function. Islet cells might be more physiologic and attractive approach for treatment of diabetes mellitus. Second, we use ginsenoside Rg3 and it is not certain whether other ginsenosides also have the same effects. Further studies are needed about islet cells and effect of other ginsenosides.
In conclusion, our present study showed that the ginsenoside Rg3 protects against INS-1 cell death from IHG and increased INS-1 cell insulin secretion. The effects of Rg3 on pancreatic β cell viability were related to decreasing apoptosis and increasing proliferation through ERK and p38 MAPKs signaling.
Abbreviations
| BrdU | = | Bromodeoxyuridine |
| FISH | = | florescence in situ hybridization |
| FOXO | = | forkhead transcription factor |
| GSIS | = | glucose-stimulated insulin secretion |
| IHG | = | intermittent high glucose |
| MAPKs | = | mitogen-activated protein kinases |
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank to Mr. John Hotra for assistance with the English language.
References
- Park S, Ahn IS, Kwon DY, Ko BS, Jun WK. Ginsenoside Rb1 and Rg1 suppress triglyceride accumulation in 3T3-L1 adipocytes and enhance β-cell insulin secretion and viability in Min6 cells via PKA-dependent pathways. Biosci Biotechnol Biochem 2008; 72:2815-23; PMID:18997435; http://dx.doi.org/10.1271/bbb.80205
- Nah SY, Park HJ, McCleskey EW. A trace component of ginseng that inhibits Ca2+ channels through a pertussis toxin-sensitive G protein. Proc Natl Acad Sci USA 1995; 92:8739-43; PMID:7568008; http://dx.doi.org/10.1073/pnas.92.19.8739
- Attele AS, Wu JA, Yuan CS. Ginseng pharmacology: multiple constituents and multiple actions. Biochem Pharmacol 1999; 58(11):1685-93; PMID:10571242; http://dx.doi.org/10.1016/S0006-2952(99)00212-9
- Cha JY, Park EY, Kim HJ, Park SU, Nam KY, Choi JE, Jun HS. Effect of white, Taegeuk, and red ginseng root extracts on insulin-stimulated glucose uptake in muscle cells and proliferation of β-cells. J. Ginseng Res 2010; 34:192-7; PMID:NOT_FOUND; http://dx.doi.org/10.5142/jgr.2010.34.3.192
- Chen F, Chen Y, Kang X, Zhou Z, Zhang Z, Liu D. Anti-apoptotic function and mechanism of ginseng saponins in rattus pancreatic β-cells. Biol Pharm Bull 2012; 35:1568-73; PMID:22975510; http://dx.doi.org/10.1248/bpb.b12-00461
- Yuan HD, Kim JT, Kim SH, Chung SH. Ginseng and diabetes: the evidences from in vitro, animal and human studies. J Ginseng Res 2012; 36:27-39; PMID:23717101; http://dx.doi.org/10.5142/jgr.2012.36.1.27
- Kim M, Ahn BY, Lee JS, Chung SS, Lim S, Park SG, Jung HS, Lee HK, Park KS. The ginsenoside Rg3 has a stimulatory effect on insulin signaling in L6 myotubes. Biochem Bio phys Res Commun 2009 Nov 6; 389(1):70-3; PMID:19699714; http://dx.doi.org/10.1016/j.bbrc.2009.08.088
- Park MW, Ha J, Chung SH. 20(S)-ginsenoside Rg3 enhances glucose-stimulated insulin secretion and activates AMPK. Biol Pharm Bull 2008; 31(4):748-51; PMID:18379076; http://dx.doi.org/10.1248/bpb.31.748
- Wadén J, Forsblom C, Thorn LM, Gordin D, Saraheimo M, Groop PH, Finnish Diabetic Nephropathy StudyGroup. A1C variability predicts incident cardiovascular events, microalbuminuria, and overt diabetic nephropathy in patients with type 1 diabetes. Diabetes 2009 Nov; 58(11):2649-55; PMID:19651819; http://dx.doi.org/10.2337/db09-0693
- Ma WY, Li HY, Pei D, Hsia TL, Lu kc, Tsai LY, Wei JN, SU CC. Variability in hemoglobin A1c predicts all-cause mortality in patients with type 2 diabetes. J Diabetes Complications 2012; 26:296-300; PMID:22626873; http://dx.doi.org/10.1016/j.jdiacomp.2012.03.028
- Monnier L, Mas E, Ginet C, Michel F, Villon L, Cristol JP, Colette C. Activation of oxidative stress by acute glucose fluctuations compared with sustained chronic hyperglycemia in patients with type 2 diabetes. JAMA 2006 Apr 12; 295(14):1681-7; PMID:16609090; http://dx.doi.org/10.1001/jama.295.14.1681
- Kim M, Chung H, Yoon C, Lee E, Kim T, Kim T, Kwon M, Lee S, Rhee B, Park J. Increase of INS-1 cell apoptosis under glucose fluctuation and the involvement of FOXO–SIRT pathway. Diabetes Res Clin Pract 2012; 98(1):132-9; PMID:22579214
- Chang L, Karin M. Mammalian MAP kinase signaling cascades. Nature 2001 Mar 1; 410(6824):37-40; PMID:11242034; http://dx.doi.org/10.1038/35065000
- Pearson G, Robinson F, Beers Gibson T, Xu BE, Karandikar M, Berman K, Cobb MH. Mitogen-activated protein (MAP) kinase pathways: regulation and physiological functions. Endocr Rev 2001 Apr; 22(2):153-83; PMID:11294822
- Kim EK, Choi EJ. Pathological roles of MAPK signaling pathways in human diseases. Biochim Biophys Acta 2010 Apr; 1802(4):396-405; PMID:NOT_FOUND; http://dx.doi.org/10.1016/j.bbadis.2009.12.009
- Zhou L, Cai X, Han X, Ji L. P38 Plays an Important Role in Glucolipotoxicity-Induced Apoptosis in INS-1 Cells. J Diabetes Res 2014; 2014:834528; PMID:24734256
- Larsen CM, Wadt KA, Juhl LF, Andersen HU, Karlsen AE, Su MS, Seedorf K, Shapiro L, Dinarello CA, Mandrup-Poulsen T. Interleukin-1beta-inducedratpancreatic islet nitricoxide synthesis requires both the p38 and extra cellular signal-regulated kinase1/2 mitogen-activated protein kinases. J Biol Chem 1998 Jun 12; 273(24):15294-300; http://dx.doi.org/10.1074/jbc.273.24.15294
- Hou ZQ, Li HL, Gao L, Pan L, Zhao JJ, Li GW. Involvement of chronic stresses in rat islet and INS-1 cell glucotoxicity induced by intermittent high glucose. Mol Cell Endocrinol 2008 Sep 10; 291(1-2):71-8; PMID:18485584; http://dx.doi.org/10.1016/j.mce.2008.03.004
- Kim MK, Jung HS, Yoon CS, Ko JH, Jun HJ, Kim TK, Kwon MJ, Lee SH, Ko KS, Rhee BD, et al. The effect of glucose fluctuation on apoptosis and function of INS-1 pancreatic beta cells. Korean Diabetes J 2010; 34:47-54; PMID:20532020; http://dx.doi.org/10.4093/kdj.2010.34.1.47
- Kim SJ, Yuan HD, Chung SH. Ginsenoside Rg1 suppresses hepatic glucose production via AMP-activated protein kinase in HepG2 cells. Biol Pharm Bull 2010; 33(2):325-8; PMID:20118562; http://dx.doi.org/10.1248/bpb.33.325
- Lee HR, Kim JE, Lee YJ, Kwak MH, Im DS, Hwang DY. Red Liriopeplatyphylla stimulated the insulin secretion through the regulation of calcium concentration in rat insulinoma cells and animal models. Lab Anim Res 2013 Jun; 29(2):84-95; PMID:23825481; http://dx.doi.org/10.5625/lar.2013.29.2.84
- Kim K, Park M, Kim HY. Ginsenoside Rg3 suppresses palmitate-induced apoptosis in MIN6N8 pancreatic beta-cells. J Clin Bio chem Nutr 2010; 46(1):30-5; PMID:20104262; http://dx.doi.org/10.3164/jcbn.09-49
- Luo JZ, Luo L. American ginseng stimulates insulin production and prevents apoptosis through regulation of uncoupling protein-2 in cultured beta cells. Evid Based Complement Alternat Med 2006 Sep; 3(3):365-72; PMID:16951721; http://dx.doi.org/10.1093/ecam/nel026
- Chen F, Chen Y, Kang X, Zhou Z, Zhang Z, Liu D. Anti-apoptotic function and mechanism of ginseng saponins in Rattus pancreatic β-cells. Biol Pharm Bull 2012; 35(9):1568-73; PMID:22975510; http://dx.doi.org/10.1248/bpb.b12-00461
- Cnop M, Welsh N, Jonas JC, Jörns A, Lenzen S, Eizirik DL. Mechanisms of pancreatic beta-cell death in type1 and type2 diabetes: many differences, few similarities. Diabetes 2005 Dec; 54(Suppl 2):S97-107; PMID:16306347
- Hashimoto R, Yu J, Koizumi H, Ouchi Y, Okabe T. Ginsenoside Rb1 Prevents MPP(+)-Induced Apoptosis in PC12 Cells by Stimulating Estrogen Receptors with Consequent Activation of ERK1/2, Akt and Inhibition of SAPK/JNK, p38 MAPK. Evid Based Complement Alternat Med 2012; 2012:693717; PMID:23024694; http://dx.doi.org/10.1155/2012/693717
- Zhang LH, Jia YL, Lin XX, Zhang HQ, Dong XW, Zhao JM, Shen J, Shen HJ, Li FF, Yan XF, et al. AD-1, a novel ginsenoside derivative, shows anti-lung cancer activity via activation of p38 MAPK pathway and generation of reactive oxygen species. Biochim Biophys Acta 2013 Aug; 1830(8):4148-59; PMID:23583729; http://dx.doi.org/10.1016/j.bbagen.2013.04.008
- Lv S, Yi PF, Shen HQ, Zhang LY, Dong HB, Wu SC, Xia F, Guo X, Wei XB, Fu BD. Ginsenoside Rh2-B1 stimulates cell proliferation and IFN-γ production by activating the p38 MAPK and ERK-dependent signaling pathways in CTLL-2 cells. Immuno Pharmacol Immuno Toxicol 2014 Feb; 36(1):43-51; PMID:24294901; http://dx.doi.org/10.3109/08923973.2013.864669
- Kim DY, Jung MS, Park YG, Yuan HD, Quan HY, Chung SH. Ginsenoside Rh2(S) induces the differentiation and mineralization of osteoblastic MC3T3-E1 cells through activation of PKD and p38 MAPK pathways. BMB Rep 2011 Oct; 44(10):659-64; PMID:22026999; http://dx.doi.org/10.5483/BMBRep.2011.44.10.659

