

ABSTRACT
Since the report of the Edmonton protocol in 2000, islet transplantation has been implemented worldwide, and xenotransplantation using porcine islets has also been reported. In addition, many basic experiments using pancreatic islets and exocrine tissue after isolation have been reported. Recently, exocrine cells have been found to be essential for inducing the differentiation of pancreatic islets. Therefore, the importance of the culture conditions for pancreatic tissue when conducting experiments using pancreatic tissue is also increasing. In this study, we focused on the coat material and examined the adhesive properties of porcine pancreatic islets and exocrine tissue after isolation. Porcine islet isolation was performed, and isolated islets (purity ≥95%) and exocrine tissue (purity ≥99%) were used to achieve adhesion to several extracellular matrixes, fibronectin, collagen type I, collagen type IV, laminin I, fibrinogen, and bovine serum albumin (BSA). DMEM with 0.5% FBS was used as the assay medium. For exocrine tissue, the adhesion was promoted in fibronectin, collagen type I, laminin I, and fibrinogen. The adhesive ability to fibronectin was more than twice that to BSA, while the adhesive ability to collagen type I, laminin I, and fibrinogen was less than twice that to BSA. For islets, the adhesive ability to fibronectin was weaker than that of exocrine tissue. Furthermore, the adhesion effect in fibronectin was obtained within 30 minutes and in medium containing little serum for both islets and exocrine tissues. These data suggest that fibronectin may be useful for the adhesion of pancreatic tissue.
Introduction
Pancreatic islet transplantation provides an alternative to pancreatic transplantation for tight glucose control in type 1 diabetic patients who experience hypoglycemic unawareness despite the best care.Citation1-4 However, islet transplantation from two or more donors is usually required to achieve euglycemia because current isolation techniques usually recover fewer than half of the islets from a given pancreas. The processes of isolation and culture disturb islet extracellular tissue and disrupt internal vascularization and innervation. The loss of extracellular matrix (ECM) during the isolation process is one of the many reasons for problems occurring after transplantation. The ability to reestablish critical cell-ECM signaling may improve the current islet culture techniques used between islet isolation and transplantation. It has been reported that collagen type IVCitation5 and lamininCitation6 contributed to greater insulin release than BSA and that islets with collagen type I or with fibronectin exhibited less apoptosis and greater insulin secretion than BSA.Citation7 Laminin is the major component of the basement membrane, whereas fibronectin is found more abundantly beneath endothelial cells and epithelial ducts in the interstitial matrix.Citation8
Recently, molecular biological methods, such as genome editing (e.g. CRISPR/Cas9 system), gene therapy, and cell reprogramming, have provided more opportunities for clinical applications beyond the scope of traditional research. With the development and improvements of these technologies, many basic experiments using not only pancreatic islets but also exocrine tissue after isolation have been performed. In addition, few studies have so far compared the cell adhesion properties of islets and exocrine tissues immediately after pancreatic islet isolation. We place such importance on cell adhesion because many molecular biological techniques for cells are performed under cell adhesion conditions. Regenerative medicine associated with the establishment of pluripotent stem cellsCitation9-12 and gene therapyCitation13-15 may offer new treatments for severe pancreatic diseases, such as diabetes mellitus.
However, gene transfer into pancreas cells in vitro is difficult because pancreatic cells cannot be cultured for a long period after islet isolation.Citation16 It is also difficult to establish pancreatic human induced pluripotent stem cells (iPSCs) and iPSC-like cells.Citation9,Citation10
Developmentally, both exocrine tissue and pancreatic islets are differentiated from pancreatic buds. Recently, exocrine tissue has been reported to be essential for inducing the differentiation of pancreatic islets.Citation17 Therefore, increased attention is being focused on the culture conditions for pancreatic islets as well as exocrine tissue.
In this study, we focused on the coat material and examined the adhesive properties of pancreatic islets and exocrine tissue after isolation. Since the use of human pancreatic tissue from cadaveric donors for research is unlawful in Japan, we used porcine pancreatic tissue in this study.
Results
Adhesive properties of islets and exocrine tissue to ECM proteins
To evaluate the adhesive properties of islets and exocrine tissue to ECM, the islets and exocrine tissue (total cell number: 1.0 × 105) were seeded into 48-well plates for 1 to 3 h with fibronectin, collagen type I, collagen type IV, laminin I, fibrinogen, and bovine serum albumin (BSA; control). Adherent cells were displaced into 96-well plates and quantified by an MTT assay at OD 560 nm after extraction. Both the islets and exocrine tissue were attached to all of the ECM components within 3 h (). The exocrine tissue adhered particularly strongly to fibronectin.
Figure 1. Adhesive property of islets and exocrine tissue to extracellular matrix. (A) Cell adhesion assays of both islets and exocrine tissue. The cells were evaluated following attachment to 48-well culture dishes coated with fibronectin, collagen type I, collagen type IV, laminin I, fibrinogen, and BSA at 1.0 × 105 cells/well in DMEM containing 0.5% FBS for 1–3 h (n = 1). (B) Cell adhesion assays of both islets and exocrine tissue. Cells were seeded onto fibronectin-coated (6.00 μg/well) or non-coated 6-well plates at 5.0 × 105 cells/well in DMEM containing 10.0% FBS for 3 h (n = 3). (C) Cell adhesion assays of both islets and exocrine tissue. Cells were seeded onto fibronectin-coated (1.50 μg/well) 96-well plates at 1.0 × 104 cells/well in DMEM (25–100 μl/well) containing 0.5% FBS for 2 h. (n = 4) Adherent cells were displaced into 96-well plate, stained with MTT solution, and quantified at OD 560 nm after extraction. The data are presented as the mean ± standard error (SE). **P < 0.01.
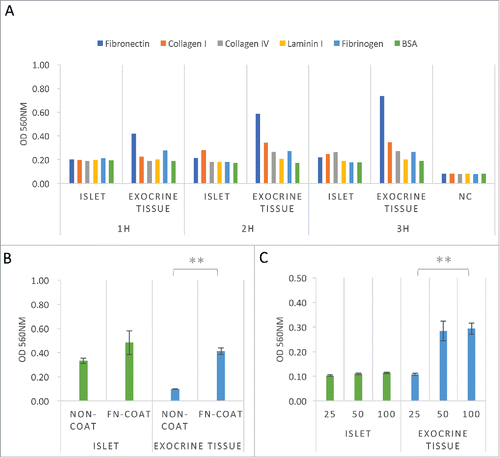
To confirm the adhesive ability of the pancreatic tissue to fibronectin, islets and exocrine tissue (total cell number: 5.0 × 105) were seeded into 6-well plates for 3 h. Adherent cells were displaced into 96-well plates and quantified at OD 560 nm after extraction. The presence of a fibronectin coat accelerated the capacity of the exocrine tissue to bind to the culture dish at 3 h (). To evaluate whether or not the volume of the medium affected the adhesive ability of the pancreatic tissue to fibronectin, islets and exocrine tissue (total cell number: 1.0 × 104) were seeded into 96-well plates in 25, 50, or 100 μl medium for 2 h. A low volume of medium (25 μl) affected the binding capacity of exocrine tissue to fibronectin (). These data suggest that fibronectin coating induced the cell adhesion of pancreatic tissue, especially exocrine tissue.
Time course of the adhesive ability of islets and exocrine tissue to fibronectin
To evaluate the time course of the adhesive ability of islets and exocrine tissue to fibronectin, isolated exocrine tissue (total cell number: 1.0 × 105) was seeded into 96-well plates with fibronectin for 10–30 min and quantified by an MTT assay at OD 560 nm after extraction. The adhesion of both islets and exocrine tissue was significantly increased at 30 min, and 55%–65% of islets and 65% of exocrine tissue were found to have adhered to fibronectin ().
Figure 2. Time course of the adhesive ability of islets and exocrine tissue to fibronectin. (A) Cell adhesion assays of both islets and exocrine tissue. Cells were seeded onto fibronectin-coated (1.50 μg/well) 96-well plates at 1.0 × 105 cells/well in DMEM containing 0.5% FBS after 10–30 min culture (n = 6). The data are presented as the mean ± standard error (SE). *P < 0.05 or **P < 0.01. (B) Cell adhesion assays of both islets and exocrine tissue. Cells were seeded onto fibronectin-coated (1.50 μg/well) 96-well plates at 1.0 × 104 cells/well in DMEM containing 0.5% FBS after 1–3 h culture (n = 6). (C) Cell viability assays of both islets and exocrine tissue. The cells were seeded onto fibronectin-coated (1.50 μg/well) 96-well plates at 1.0 × 104 cells/well in DMEM containing 0.5% FBS after 1–3 h culture (n = 6). (D) Cell adhesion assays of both islets and exocrine tissue. Cells were seeded onto fibronectin-coated (2.5, 5.0, 10.0, 20.0 mg/ml × 150 μl/well) 96-well plates for 1–3 h at 1.0 × 104 cells/well. Statistical analyses were performed using the central value (after 2 h culture) for each experimental condition (n = 6). Adherent cells were stained with MTT solution and quantified at OD 560 nm after extraction (A, B, D) or by WST-8 assay solution and quantified at OD 450 nm after extraction (C). The data are presented as the mean ± standard error (SE). *P < 0.05 or **P < 0.01.
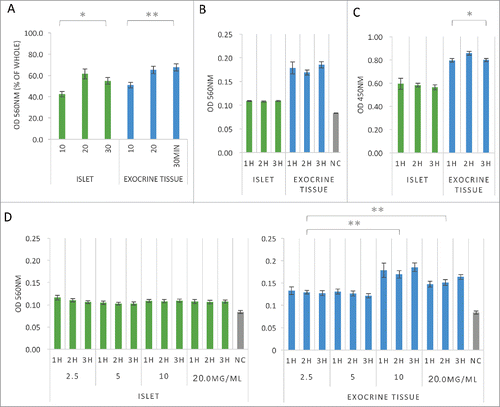
To characterize the adhesive activity of fibronectin, islets and exocrine tissue (total cell number: 1.0 × 104) were cultured on fibronectin-coated 96-well dishes for 1–3 h and quantified by an MTT assay. MTT [3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide] was used as a reagent to evaluate the metabolic activity of the viable cells. MTT is reduced by dehydrogenase in viable cells and is converted to water-insoluble formazan crystals. After dissolving this formazans in organic solvent, the absorbance at 570 nm is measured. An increase in the number of living cells results in an increase in the amount of formazan that is formed. After extraction, the absorbance (which is correlated with the number of living cells) is measured at OD 560 nm. In parallel, a WST-8 assay (measuring the absorbance at 450 nm) was performed using WST-8 [2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt] as a reagent. The measurements shown are proportional to the number of viable cells. Thus, it is possible to calculate the approximate value using the absorbance value of the calibration curve for the number of viable cells. For this assay, islets and exocrine tissue (total cell number: 1.0 × 104) were again cultured on fibronectin-coated 96-well dishes for 1–3 h before being quantified at OD 450 nm after extraction. The adhesion of both islets and exocrine tissue was steady for 1–3 h (). A significant decrease in the cellular metabolism activity in the exocrine tissue was observed from 2 to 3 h ().
To evaluate the relationship between the adhesive ability and the concentration of fibronectin, islets and exocrine tissue (total cell number: 1.0 × 104) were seeded into 96-well plates with several concentrations of fibronectin (2.5 to 20.0 mg/ml) for 1–3 h and quantified by an MTT assay at OD 560 nm after extraction. The adhesive ability of exocrine tissue was significantly higher at 10.0 to 20.0 mg/ml of fibronectin than at 2.5 mg/ml of fibronectin, while the adhesive ability of the islets was roughly the same, regardless of the concentration of fibronectin ().
Adhesive properties of islets and exocrine tissue to gelatin
Gelatin is a form of hydrolyzed collagen and widely used as a cell culture coat material. iPS cells in feeder culture are often cultured on gelatin-coated dishes. Thus, it is important to evaluate gelatin-coated dishes and fibronectin-coated dishes when culturing cells for regenerative medicine. In addition, pancreatic islets are reported to be activated by collagen type I- and type IV-coated dishes, which are the main components of gelatin and there are reports that promote the use of gelatin.Citation18 Thus, in the present study, experiments were carried out using gelatin-coated dishes. To investigate the adhesive activity to gelatin, islets and exocrine tissue (total cell number: 1.0 × 104) were cultured on gelatin (0.1%)-coated 96-well dishes for 1–3 h and quantified by a WST-8 assay at OD 450 nm after extraction. A significant decrease in the cellular metabolism activity was observed at 3 h ().
Figure 3. Adhesive properties of islets and exocrine tissue to gelatin. (A) Cell viability assays of both islets and exocrine tissue. Cells were seeded onto gelatin-coated (0.1%) 96-well plates at 1.0 × 104 cells/well in DMEM containing 0.5% FBS after 1–3 h culture. Adherent cells were stained by WST-8 assay solution and quantified at OD 450 nm after extraction (n = 6). (B) Cell adhesion assays of both islets and exocrine tissue. Cells were seeded onto gelatin-coated (0.05–0.10%) 96-well plates at 1.0 × 104 cells/well in DMEM containing 0.5% FBS. Adherent cells were stained with MTT solution and quantified at OD 560 nm after extraction (n = 6). The data are presented as the mean ± standard error (SE). *P < 0.05 or **P < 0.01.
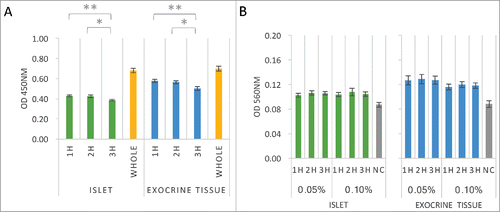
To evaluate the relationship between the adhesive ability and the concentration of gelatin, islets and exocrine tissue (total cell number: 1.0 × 104) were seeded into 96-well plates with several concentrations of gelatin (0.05–0.1%) for 1–3 h and quantified by an MTT assay at OD 560 nm after extraction. The adhesive ability of both islets and exocrine tissue was roughly the same, regardless of the concentration of gelatin (). Bovine gelatin is often used as a coating material in cell culture experiments. However, in cultures of islets and exocrine tissues, bovine gelatin-coated dishes are thought to have a weak promoting effect on cell adhesion and less effect with regard to the promotion of cell survival.
The insulin secretory response of islets to culture on fibronectin and gelatin
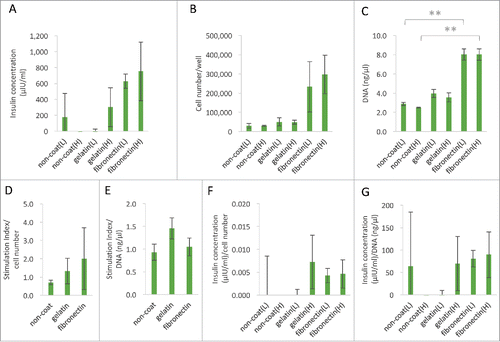
It is well known that islets lose their functionality in some ECM components, and that only specific ECM components support insulin secretion.Citation18 A glucose tolerance test was performed to investigate the function of pancreatic islets seeded in fibronectin- or gelatin-coated wells.Citation19 To assess the islet quality in each group (cultured on non-coated, gelatin [0.1%]-coated, fibronectin [10.0 mg/ml]-coated 24-well plates) for 2 h in vitro, the insulin concentration in medium containing 2.8 or 25 mM glucose was measured (), and then the attached cell number () and amount of DNA of the attached cell () after 2 h were determined. In the “gelatin coat” and “fibronectin coat” groups, the medium concentration of insulin increased under high-glucose conditions. However, no significant differences were noted among the three groups with regard to the insulin concentration or the stimulation index. These data suggest that the islet quality was similar among the three groups in vitro.
Figure 4. The function of islets for fibronectin. (A) The insulin concentrations contained in low glucose medium and high glucose medium after 2 hours of islet separation using non-coated, gelatin (0.1%)-coated and fibronectin (10.0 mg/ml)-coated 24-well plates (n = 6). The data are presented as the mean ± standard error (SE). (B and C) Measurements of the islet cell number and DNA amount. The cells were seeded onto non-coated, gelatin-coated (0.1%) and fibronectin-coated (10.0 mg/ml) 24-well plates at 500 IEQ. Number of cells: 5.0 × 103/well in low glucose medium and high glucose medium after 2 h of culture. The total number of cells and the amount of DNA were measured (n = 6). The data are presented as the mean ± standard error (SE). *P < 0.05. (D and E) The stimulation index corrected by cell number and DNA quantity of the isolated islets. Using non-coated, gelatin (0.1%)-coated and fibronectin (10.0 mg/ml)-coated 24-well plates (n = 6). The data are presented as the mean ± standard error (SE). (F and G) The insulin concentrations corrected by cell number and DNA quantity of the isolated islets. Using non-coated, gelatin (0.1%)-coated and fibronectin (10.0 mg/ml)-coated 24-well plates (n = 6). The data are presented as the mean ± standard error (SE).
Measurement of cell number and DNA quantity of islets in cultures on fibronectin and gelatin
To assess the cell number and DNA quantity in cultures made with medium containing 2.8 or 25 mM glucose on fibronectin-coated and gelatin-coated dishes, islets (500 IEQ; total cell number: 5.0 × 103) were cultured in each group (cultured on non-coated, gelatin [0.1%]-coated and fibronectin [10.0 mg/ml]-coated 24-well plates) for 2 h in vitro, and the number of adhesion cells was quantified using a Luna Automated Cell Counter. Total pancreatic islet significantly promoted adhesion to the fibronectin-coated dishes, both in low glucose medium and high glucose medium.
Expression of integrin in islets and exocrine tissue
Since various integrins are known to bind to fibronectin, we evaluated the expression of integrin chains in islets and exocrine tissue using reverse transcription polymerase chain reaction (RT-PCR) and quantitative RT-PCR (qRT-PCR). The commercially available primers of porcine integrins for real-time PCR analyses are integrins αV, β1, and β3. On qRT-PCR, the integrins αV, β1 and β3 were expressed in islets, and the integrins αV and β1 were expressed in exocrine tissue (). Other types of integrins were evaluated by RT-PCR. The islets expressed integrins α1, α2, α3, α4, α5, α6, α10, α11, αL, αM, αV, β1, β2, β3, β4, β5, β6, β7, and β8. The exocrine tissue expressed integrins α1, α2, α5, α6, α8, α9, α10, α11, αL, αV, β1, β3, β4, β5, β7, and β8. Two subtype αA/Bs have been reported for humanCitation20,Citation21 and mouseCitation22 integrin α6, and 2 subtype βB/βCs have been reported for humanCitation23 integrin β4. Therefore, double bands of integrin α6 and β4 might be detected ().
Figure 5. Expression of integrin in islets and exocrine tissue. (A) The expression of porcine integrins αV, β1, and β3 in both islets and exocrine tissue was evaluated by a quantitative RT-PCR analysis using commercially available primers (left graph; n = 1). The expression of integrin αV, β1, and β2 was evaluated by RT-PCR assay. GAPDH housekeeping genes were used as denominators for the comparison of samples (right figure; n = 1). (B) The expression of integrins α1, α2, α3, α4, α5, α6, α8, α9, α10, αE, αL, αM, αV, 2αb, αX, β1, β2, β3, β4, β5, β6, β7, and β8 in both islets (upper figure) and exocrine tissue (lower figure) (n = 1). GAPDH housekeeping genes were used as denominators for the comparison of samples.
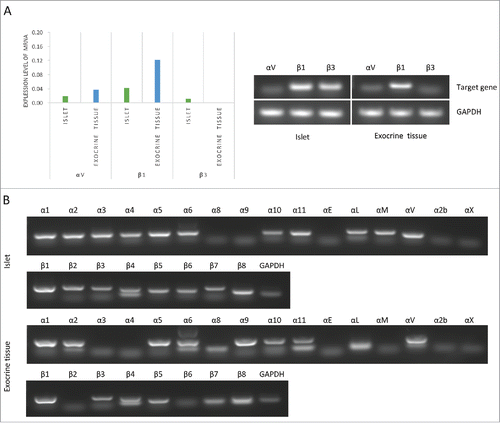
The islets expressed α4β1, α5β1, αVβ1, αVβ3, αVβ6, and α4β7, which were integrin chains of fibronectin receptor, and the exocrine tissue expressed α5β1, α8β1, αVβ1, and αVβ3, which were integrin chains of fibronectin receptor. Therefore, both islets and exocrine tissue had a high adhesive ability to fibronectin.
Discussion
It has been reported that collagen I-αVβ3, collagen IV-αVβ1, fibrinogen-αVβ3, fibronectin-αVβ1, αVβ3, α5β1, laminin-α1β1, α3β1, α5β1, α6β1, and vitronectin-αVβ1, αVβ5 promote cell adhesion.Citation5,Citation24-27 We evaluated the expression of integrins by both quantitative RT-PCR and RT-PCR, since the only commercially available porcine integrin primers were integrin αV, β1, and β3. For RT-PCR, we designed the mRNA sequence from the database of National Center for Biotechnology Information (NCBI) using the Primer 3 Plus software program (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi/). After electrophoresis of the gel, we confirmed a double band, which is a feature of a subtype of integrin β4 in islets. In addition, we confirmed subtypes for integrin α2, α6, α11, and β4 in exocrine tissue. This report is the first to confirm that porcine pancreatic cells have integrin subtypes. It has been reported that human and mouse pancreatic cells have subtypes of integrin α6 and β4.Citation20-22
Integrin-ECM interactions are well-known regulators of the islet cell survival and function. The most extensively studied integrin in islet biology is β1. Integrin β1 promotes the expression of Pdx1 and Nkx6.1, which are the master genes of the islet.Citation28 In pancreatic cells, the twelve types of α subunits of integrin are involved in the regulation of morphogenesis, differentiation, proliferation, and survival.
It has been reported that integrin α3β1 and α6β1, both major laminin receptors, mediate the attachment and spreading of rat islet cells, improving the cell survival and insulin secretion.Citation24,Citation29 Other reports have shown that binding of collagen type IV-integrin α1β1,Citation5,Citation30 the major collagen receptor, on human or mouse pancreatic β cells promotes their insulin secretions. Our study showed that both islets and exocrine tissue were easily adhesive to fibronectin, and both cells expressed integrin α5β1, αVβ1, and αVβ3. These are the major fibronectin receptors. Our data and other previous reports suggest that pancreatic islets may be effectively adhesive to fibronectin as well as laminin and collagen and that integrin β1 may be a major factor that plays an important role in the process of cell adhesion in both types of cells. This result suggests that fibronectin has favorable effects on producing iPS cells and iPS-like cells using pancreatic islets and exocrine tissues. However, the fibronectin coat had no marked effect of promoting insulin secretion according to the glucose tolerance test (, and ).
Few studies have evaluated the adhesive properties on exocrine tissue. Our study showed that exocrine tissue after isolation strongly expressed integrin αV and β1 mRNA, while integrin β3 mRNA expression was lower than that of integrin αV and β1 (). These data suggest that the strong cell adhesion of exocrine tissue to fibronectin may originate not from integrin αVβ3, but rather from αVβ1. However, both integrins αVβ1 and αVβ3 were expressed at lower levels in islets than in exocrine tissue (). It has been reported that integrin αVβ3 of human β cells plays no major role in cell adhesion or migration.Citation26
In conclusion, fibronectin as a coating material promotes adhesion and cell survival activity of islets and exocrine tissue. Both islet and exocrine tissue express integrin αVβ1, αVβ3, and α5β1, which are receptors of fibronectin. The usage of fibronectin-based coating materials may be useful for culturing pancreatic cells for genetic manipulation, and iPS cell and iPS-like cell production.
Materials and methods
Reagents
University of Wisconsin solution (Viaspan) was obtained from Astellas Pharma Inc. (Tokyo, Japan). Fetal bovine serum (FBS) was obtained from BioWest (Nuaille, France). Iodixanol (OptiPrep) was obtained from AXIS-SHIELD PoC AS (Oslo, Norway). The Gradient Mixer and Ricordi Isolator Tubing Set were obtained from Biorep Technologies (Miami, FL, USA). Dulbecco's modified Eagle medium (DMEM), RPMI-1640 medium and 100 bp DNA Ladder One were obtained from Nacalai Tesque (Kyoto, Japan). Dithizone and gelatin from bovine skin were obtained from Sigma-Aldrich Co. LLC (St. Louis, MO, USA). Fibronectin Solution from Human Plasma was purchased from WAKO (Osaka, Japan). The plastic dishes were obtained from TPP (Trasadingen, Switzerland). All other materials used were of the highest commercial grade.
Pancreas procurement and islet isolation
Three-year-old porcine pancreata (female, n = 5) were obtained from a local slaughterhouse. After removing the pancreas, we immediately inserted a cannula into the main pancreatic duct. The pancreas was weighed, and 1 ml/g pancreas weight of modified Kyoto (MK) solution (Otsuka Pharmaceutical Factory Inc., Naruto, Japan) was infused through the intraductal cannula.Citation31 The pancreata were placed into the MK solution container at 4°C for about 18 h until the islet isolation procedure.Citation32
Islet isolation was conducted as previously described.Citation3 Briefly, the ducts were perfused in a controlled fashion with a cold enzyme blend of Liberase MTF (1.0 mg/ml) with thermolysin (0.075 mg/ml) (Roche Diagnostics Corporation, Indianapolis, IN, USA). The distended pancreas was then placed in a Ricordi chamber and gently shaken. Once digestion was confirmed to be complete, the dilution solution (Center for Promotion of Education and Science, Hiroshima, Japan) was introduced into the system. The system was then cooled to stop further digestive activity. The digested tissue was collected in flasks containing 5% fetal bovine serum (FBS; GIBCO-Invitrogen, Carlsbad, CA, USA). Islets were purified with a continuous density gradient of iodixanol (Optiprep®; Sigma-Aldrich)-UW solution as previously reported.Citation19,Citation33-35 The islet yield was determined with dithizone staining (2 mg/ml).
Cell adhesion assays
Cell adhesion was measured using a CytoSelect 48-Well Cell Adhesion Assay (Cell Biolabs, Inc., San Diego, CA, USA) in accordance with the manufacturer's instructions. Briefly, the cells were seeded into 48-well plates coated with human fibronectin, human collagen IV, human fibrinogen, mouse laminin I, bovine collagen I, and bovine serum albumin (BSA) and then incubated for 1–3 h. Each well received 1 × 105 cells/500 μL in DMEM containing 0.5% FBS. At the end of the incubation period, the medium was aspirated from each well. Each well was gently washed 3 times with PBS. Immediately, Cell Stain Solution (MTT assay) was added to each well, and the cells were incubated for 10 minutes at room temperature. At the end of the incubation period, the Cell Stain Solution was removed, and the wells were washed with warm PBS (37 deg;C). Subsequently, Extraction Solution (200 μl) was added to each 96-well, and the absorbance was measured with a micro plate reader (Infinite M200 PRO; Tecan Group Ltd., Männedorf, Switzerland) at a wavelength of OD 560 nm.
Cell viability assays
Cell viability was measured using Cell Count Reagent SF (WST-8 assay) and an MTT Cell Count Kit (Nacalai Tesque, Kyoto, Japan) in accordance with the manufacturer's instructions. Briefly, the cells were seeded into 96-well plates coated with human fibronectin or bovine gelatin and incubated for 10–30 minutes or 1–3 h. Each well received 1 × 104–5 cells/100 μL in DMEM containing 0.5% or 10.0% FBS. At the end of the incubation period, each well was gently washed 3 times with PBS, Cell Count Reagent or MTT solution was added to each well, and the cells were incubated for 2–4 h in the incubator (37 deg;C). The absorbance was measured with a micro plate reader at a wavelength of 450 or 560 nm. The cell count was determined using a Luna Automated Cell Counter (Logos Biosystems, Gyeonggi-do, South Korea).
The assessment of the islet function
The crude number of islets in each diameter class was determined by counting the islets after dithizone staining using an optical graticule (Olympus, Tokyo, Japan). The crude number of islets was then converted to the standard number of islet equivalents (IEQ) (diameter standardizing to 150 μm).Citation36 The islet function was assessed by monitoring the insulin secretory response of the purified islets during glucose stimulation using the procedure described by Shapiro et al.Citation1,Citation37 Briefly, 500 IEQ was seeded into each well. After 2 h, each well was washed twice with PBS, and any non-adherent cells were removed. The attached cells incubated with either 2.8 or 25 mM glucose in RPMI-1640 (Sigma-Aldrich) for 2 h at 37°C and 5% CO2. The supernatant was collected, and the insulin levels were determined using a commercially available chemiluminescence enzyme immunoassay (CLEIA) system (LUMIPULSE PRESTO II; FUJIREBIO INC, Tokyo, Japan). The insulin concentration in each group was calculated by subtracting the average value of the non-coat group. The stimulation index was calculated by determining the ratio of the insulin released from the islets under high-glucose conditions to the insulin released under low-glucose conditions. The stimulation index was calculated from the original data. DNA was extracted from the cells that adhered to the 24-well plate. The extracted DNA was eluted in a volume of 100 μl, and then the concentration was measured. DNA content was quantified with a BioSpec-nano (SHIMADZU, Kyoto, Japan).
Real time PCR and RT-PCR analyses
Both isolated islets and exocrine tissue populations (1 × 106 cells) were prepared. RNA was prepared using the RNeasy Mini Kit (Qiagen, Hilden, Germany). From the isolated RNA, cDNA was synthesized using a Quantitech Reverse Transcription kit (Qiagen) in accordance with the manufacturer's instructions. PCR amplification was performed with Taqman Universal Master Mix II without UNG (Thermo Fisher Scientific Inc., Waltham, MA, USA) or Quick Taq HS DyeMix (Toyobo Co., Ltd., Osaka, Japan). Real time PCR analyses were performed using a LightCycler 96 Real-Time PCR system (Roche, Basel, Switzerland). Porcine GAPDH primers (Ss03375435) and integrin αV primers (Ss03388773), integrin β1 primers (Ss03391119), and integrin β3 primers (Ss03391243) were obtained from Applied Biosystems (Thermo Fisher Scientific Inc.). RT-PCR analyses were performed using GeneAtlas (ASTEC, Fukuoka, Japan). Porcine GAPDH primers (Ss03375435) and the primers listed in were used.
Table 1. Sequences of primers used for the RT-PCR.
Statistical analyses
Statistical analyses were performed using Student's t-test to compare two sample means. The analyses of multiple groups (i.e. > two groups) were performed using a one-way analysis of variance with the XLSTAT software program (Addinsoft, Paris, France). Statistical significance was set at *P < 0.05 or **P < 0.01 for all tests. The data showed are representative examples of two independent experiments.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Ms. Naomi Kakazu (University of the Ryukyus) for the office processing and Ms. Saki Uema, Chie Horiguchi, Yuka Watanabe, Maki Higa (University of the Ryukyus) for technical support. This work was supported in part by the Japan Society for the Promotion of Science, Japan Agency for Medical Research and Development, The Uehara Memorial Foundation, TERUMO Foundation for Life Science and Arts, the Waksman Foundation of Japan Inc., The Naito Foundation, and Okinawa Science and Technology Promotion Center (OSTC). All authors state that they have no conflicts of interest. Author roles: study design, YN, HN; study conduct, YN, HN; data collection, YN, SC, HN; data analyses, YN, HN; data interpretation, YN, SC, HN; providing materials, NK, IS, MW; drafting the manuscript, YN, HN; revising the manuscript content, YN, HN; approving the final version of the manuscript, YN, SC, NK, IS, MW, HN. YN takes responsibility for the integrity of all data analyses.
Additional information
Funding
References
- Shapiro AM, Lakey JR, Ryan EA, Korbutt GS, Toth E, Warnock GL, Kneteman NM, Rajotte RV. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl J Med. 2000;343:230–8. doi:10.1056/NEJM200007273430401.
- Street CN, Lakey JR, Shapiro AM, Imes S, Rajotte RV, Ryan EA, Lyon JG, Kin T, Avila J, Tsujimura T, et al. Islet graft assessment in the Edmonton Protocol: implications for predicting long-term clinical outcome. Diabetes. 2004;53:3107–14. doi:10.2337/diabetes.53.12.3107.
- Noguchi H, Iwanaga Y, Okitsu T, Nagata H, Yonekawa Y, Matsumoto S. Evaluation of islet transplantation from non-heart beating donors. Am J Transplant. 2006;6:2476–82. doi:10.1111/j.1600-6143.2006.01499.x.
- Robertson RP. Islet transplantation as a treatment for diabetes - a work in progress. N Engl J Med. 2004;350:694–705. doi:10.1056/NEJMra032425.
- Kaido T, Yebra M, Cirulli V, Montgomery AM. Regulation of human beta-cell adhesion, motility, and insulin secretion by collagen IV and its receptor alpha1beta1. J Biol Chem. 2004;279:53762–9. doi:10.1074/jbc.M411202200.
- Edamura K, Nasu K, Iwami Y, Ogawa H, Sasaki N, Ohgawara H. Effect of adhesion or collagen molecules on cell attachment, insulin secretion, and glucose responsiveness in the cultured adult porcine endocrine pancreas: a preliminary study. Cell Transplant. 2003;12:439–46. doi:10.3727/000000003108746867.
- Wang RN, Rosenberg L. Maintenance of beta-cell function and survival following islet isolation requires re-establishment of the islet-matrix relationship. J Endocrinol. 1999;163:181–90. doi:10.1677/joe.0.1630181.
- Peiris H, Bonder CS, Coates PT, Keating DJ, Jessup CF. The beta-cell/EC axis: how do islet cells talk to each other? Diabetes. 2014;63:3–11. doi:10.2337/db13-0617.
- Noguchi H, Saitoh I, Tsugata T, Kataoka H, Watanabe M, Noguchi Y. Induction of tissue-specific stem cells by reprogramming factors, and tissue-specific selection. Cell Death Differ. 2015;22:145–55. doi:10.1038/cdd.2014.132.
- Saitoh I, Sato M, Soda M, Inada E, Iwase Y, Murakami T, Ohshima H, Hayasaki H, Noguchi H. Tissue-Specific Stem Cells Obtained by Reprogramming of Non-Obese Diabetic (NOD) Mouse-Derived Pancreatic Cells Confer Insulin Production in Response to Glucose. PLoS One. 2016;11:e0163580. doi:10.1371/journal.pone.0163580.
- Pagliuca FW, Millman JR, Gurtler M, Segel M, Van Dervort A, Ryu JH, Peterson QP, Greiner D, Melton DA. Generation of functional human pancreatic beta cells in vitro. Cell. 2014;159:428–39. doi:10.1016/j.cell.2014.09.040.
- D'Amour KA, Bang AG, Eliazer S, Kelly OG, Agulnick AD, Smart NG, Moorman MA, Kroon E, Carpenter MK, Baetge EE. Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat Biotechnol. 2006;24:1392–401. doi:10.1038/nbt1259.
- Bone RN, Icyuz M, Zhang Y, Zhang Y, Cui W, Wang H, Peng JB, Matthews QL, Siegal GP, Wu H. Gene transfer of active Akt1 by an infectivity-enhanced adenovirus impacts beta-cell survival and proliferation differentially in vitro and in vivo. Islets. 2012;4:366–78. doi:10.4161/isl.22721.
- Chen S, Shimoda M, Wang MY, Ding J, Noguchi H, Matsumoto S, Grayburn PA. Regeneration of pancreatic islets in vivo by ultrasound-targeted gene therapy. Gene Ther. 2010;17:1411–20. doi:10.1038/gt.2010.85.
- Zhu S, Russ HA, Wang X, Zhang M, Ma T, Xu T, Tang S, Hebrok M, Ding S. Human pancreatic beta-like cells converted from fibroblasts. Nat Commun. 2016;7:10080. doi:10.1038/ncomms10080.
- Yang YH, Johnson JD. Multi-parameter single-cell kinetic analysis reveals multiple modes of cell death in primary pancreatic beta-cells. J Cell Sci. 2013;126:4286–95. doi:10.1242/jcs.133017.
- Kodama S, Nakano Y, Hirata K, Furuyama K, Horiguchi M, Kuhara T, Masui T, Kawaguchi M, Gannon M, Wright CV, et al. Diabetes Caused by Elastase-Cre-Mediated Pdx1 Inactivation in Mice. Sci Rep. 2016;6:21211. doi:10.1038/srep21211.
- Llacua A, de Haan BJ, Smink SA, de Vos P. Extracellular matrix components supporting human islet function in alginate-based immunoprotective microcapsules for treatment of diabetes. J Biomed Mater Res A. 2016;104:1788–96. doi:10.1002/jbm.a.35706.
- Miyagi-Shiohira C, Kobayashi N, Saitoh I, Watanabe M, Noguchi Y, Matsushita M, Noguchi H. Comparison of Purification Solutions With Different Osmolality for Porcine Islet Purification. Cell Med. 2017;9:53–9. doi:10.3727/215517916X693140.
- Dydensborg AB, Teller IC, Groulx JF, Basora N, Pare F, Herring E, Gauthier R, Jean D, Beaulieu JF. Integrin alpha6Bbeta4 inhibits colon cancer cell proliferation and c-Myc activity. BMC Cancer. 2009;9:223. doi:10.1186/1471-2407-9-223.
- Gu YC, Kortesmaa J, Tryggvason K, Persson J, Ekblom P, Jacobsen SE, Ekblom M. Laminin isoform-specific promotion of adhesion and migration of human bone marrow progenitor cells. Blood. 2003;101:877–85. doi:10.1182/blood-2002-03-0796.
- Qian H, Georges-Labouesse E, Nystrom A, Domogatskaya A, Tryggvason K, Jacobsen SE, Ekblom M. Distinct roles of integrins alpha6 and alpha4 in homing of fetal liver hematopoietic stem and progenitor cells. Blood. 2007;110:2399–407. doi:10.1182/blood-2006-10-051276.
- Micheloni A, Falcioni R, Zambruno G, D'Alessio M. The human integrin beta4B and beta4C variants are not expressed in a tissue-specific manner. FEBS Lett. 2002;519:238–9. doi:10.1016/S0014-5793(02)02750-3.
- Bosco D, Meda P, Halban PA, Rouiller DG. Importance of cell-matrix interactions in rat islet beta-cell secretion in vitro - Role of alpha 6 beta 1 integrin. Diabetes. 2000;49:233–43. doi:10.2337/diabetes.49.2.233.
- Ris F, Hammar E, Bosco D, Pilloud C, Maedler K, Donath MY, Oberholzer J, Zeender E, Morel P, Rouiller D, et al. Impact of integrin-matrix matching and inhibition of apoptosis on the survival of purified human beta-cells in vitro. Diabetologia. 2002;45:841–50. doi:10.1007/s00125-002-0840-7.
- Kaido T, Perez B, Yebra M, Hill J, Cirulli V, Hayek A, Montgomery AM. Alphav-integrin utilization in human beta-cell adhesion, spreading, and motility. J Biol Chem. 2004;279:17731–7. doi:10.1074/jbc.M308425200.
- Weber LM, Hayda KN, Anseth KS. Cell-matrix interactions improve beta-cell survival and insulin secretion in three-dimensional culture. Tissue Eng Part A. 2008;14:1959–68. doi:10.1089/ten.tea.2007.0238.
- Riopel M, Krishnamurthy M, Li J, Liu S, Leask A, Wang R. Conditional beta1-integrin-deficient mice display impaired pancreatic beta cell function. J Pathol. 2011;224:45–55. doi:10.1002/path.2849.
- Kantengwa S, Baetens D, Sadoul K, Buck CA, Halban PA, Rouiller DG. Identification and characterization of alpha 3 beta 1 integrin on primary and transformed rat islet cells. Exp Cell Res. 1997;237:394–402. doi:10.1006/excr.1997.3803.
- Yap WT, Salvay DM, Silliman MA, Zhang X, Bannon ZG, Kaufman DB, Lowe WL Jr, Shea LD. Collagen IV-modified scaffolds improve islet survival and function and reduce time to euglycemia. Tissue Eng Part A. 2013;19:2361–72. doi:10.1089/ten.tea.2013.0033.
- Noguchi H, Ueda M, Hayashi S, Kobayashi N, Okitsu T, Iwanaga Y, Nagata H, Nakai Y, Matsumoto S. Ductal injection of preservation solution increases islet yields in islet isolation and improves islet graft function. Cell Transplant. 2008;17:69–81. doi:10.3727/000000008783907062.
- Noguchi H, Ueda M, Nakai Y, Iwanaga Y, Okitsu T, Nagata H, Yonekawa Y, Kobayashi N, Nakamura T, Wada H, et al. Modified two-layer preservation method (M-Kyoto/PFC) improves islet yields in islet isolation. Am J Transplant. 2006;6:496–504. doi:10.1111/j.1600-6143.2006.01223.x.
- Noguchi H, Ikemoto T, Naziruddin B, Jackson A, Shimoda M, Fujita Y, Chujo D, Takita M, Kobayashi N, Onaca N, et al. Iodixanol-controlled density gradient during islet purification improves recovery rate in human islet isolation. Transplantation. 2009;87:1629–35. doi:10.1097/TP.0b013e3181a5515c.
- Noguchi H, Naziruddin B, Shimoda M, Fujita Y, Chujo D, Takita M, Peng H, Sugimoto K, Itoh T, Kobayashi N, et al. Evaluation of osmolality of density gradient for human islet purification. Cell Transplant. 2012;21:493–500. doi:10.3727/096368911X605402.
- Miyagi-Shiohira C, Kobayashi N, Saitoh I, Watanabe M, Noguchi Y, Matsushita M, Noguchi H. The Evaluation of Islet Purification Methods That Use Large Bottles to Create a Continuous Density Gradient. Cell Med. 2017;9:45–51. doi:10.3727/215517916X693131.
- Ricordi C, Gray DW, Hering BJ, Kaufman DB, Warnock GL, Kneteman NM, Lake SP, London NJ, Socci C, Alejandro R, et al. Islet isolation assessment in man and large animals. Acta Diabetol Lat. 1990;27:185–95. doi:10.1007/BF02581331.
- Shapiro AM, Ricordi C, Hering BJ, Auchincloss H, Lindblad R, Robertson RP, Secchi A, Brendel MD, Berney T, Brennan DC, et al. International trial of the Edmonton protocol for islet transplantation. N Engl J Med. 2006;355:1318–30. doi:10.1056/NEJMoa061267.

