

ABSTRACT
Pancreatic islets play an essential role in regulating blood glucose levels. Age-dependent development of glucose intolerance and insulin resistance results in hyperglycemia, which in turn stimulates insulin synthesis and secretion from aged islets, to fulfill the increased demand for insulin. However, the mechanism underlying enhanced insulin secretion remains unknown. Glutamic acid decarboxylase 67 (GAD67) catalyzes the conversion of glutamate into γ-aminobutyric acid (GABA) and CO2. Both glutamate and GABA can affect islet function. Here, we investigated the role of GAD67 in insulin secretion in young (3 month old) and aged (24 month old) C57BL/6J male mice. Unlike young mice, aged mice displayed glucose-intolerance and insulin-resistance. However, aged mice secreted more insulin and showed lower fed blood glucose levels than young mice. GAD67 levels in primary islets increased with aging and in response to high glucose levels. Inhibition of GAD67 activity using a potent inhibitor of GAD, 3-mercaptopropionic acid, abrogated glucose-stimulated insulin secretion from a pancreatic β-cell line and from young and aged islets. Collectively, our results suggest that blood glucose levels regulate GAD67 expression, which contributes to β-cell responses to impaired glucose homeostasis caused by advanced aging.
Introduction
The pancreatic islets of Langerhans are indispensable endocrine microorgans that regulate glucose homeostasis. Age-dependent loss or dysfunction of the islets leads to elevated blood glucose levels.Citation1 Additionally, aging causes a progressive decrease in glucose tolerance and insulin sensitivity, which also promotes escalated blood glucose levels. Increased insulin synthesis and secretion occur in islet β-cells to reverse hyperglycemia.Citation2-Citation4 Although the genetic and biochemical mechanisms underlying compensatory insulin expression have been extensively investigated,Citation5 relatively fewer mechanisms are known about the factors that contribute to elevated insulin secretion.
Glutamic acid decarboxylases (GADs) catalyze the decarboxylation of glutamate to γ-aminobutyric acid (GABA) and CO2,Citation6 and are rate-limiting enzymes in the synthesis of GABA. Mammals possess two GAD isoforms, GAD65 and GAD67, with molecular weights of 65 and 67 kDa, respectively. GAD65 is expressed in rat and human islets, whereas GAD67 is predominant in mouse islets.Citation7 GAD65 and GAD67 localize primarily within the cytosol of β-cells; however, GAD65 can bind to vesicle membranes through palmitoylation of its N-terminal domain.Citation8-Citation10 Both GAD65 and GAD67 are recognized as autoantigens that induce β-cell destructive autoimmunity in type 1 diabetes.Citation11 GAD65-deficient mice showed normal glucose homeostasis,Citation12 whereas systemic GAD67 knockout mice die one day after birth.Citation13 Thus, the physiological or pathophysiological effects of a GAD67 knockout on insulin release remain unknown. Given that glutamate and GABA regulate β-cell proliferation and insulin release in mouse and human islets,Citation14,Citation15 we hypothesize that the catalytic activity of GAD67 influences β-cell function.
Previous studies evaluated GAD67 expression during prenatal, early postnatal, and young adult periods proposed,Citation16–Citation18 but the differential expression of GAD67 during aging has not been investigated. To examine a potential role of GAD67 in β-cell function during normal, non-diabetic aging, we measured the expression levels of GAD67 in young (3 month old) and aged (24 month old) C57BL/6J male mice. Our data demonstrated that GAD67 expression is highly elevated in the primary islets in aged relative to young mice. Furthermore, ex vivo experiments using islets from young and aged mice showed that GAD67 expression is glucose-dependent. Inhibition of GAD67 catalytic activity using a potent inhibitor of GAD, 3-mercaptopropionic acid (3-MPA), abolished glucose-stimulated insulin secretion (GSIS) in both β-TC6 β-cells and in islets isolated from young and aged mice. We propose that the enzymatic activity of GAD67 is important for compensatory insulin secretion to govern blood glucose levels caused by age-dependent deterioration of glucose homeostasis.
Results
Aged mice develop glucose intolerance and insulin resistance
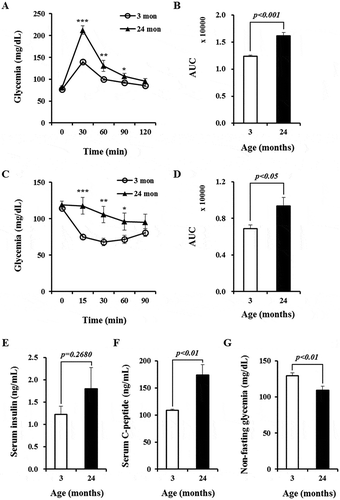
To assess the effects of aging on glucose metabolism and insulin sensitivity, we compared blood glucose homeostasis between young and aged mice. First, we performed glucose tolerance tests using a glucose load (1 g/kg). Relative to young mice, aged mice showed significantly higher blood glucose levels at 30, 60, and 90 minutes after glucose injection () and a dramatic difference (by 1.3-fold) in area under curve (AUC) (). These data suggest an age-dependent impairment of glucose tolerance. Similarly, insulin tolerance tests revealed that aged mice had significantly higher blood glucose levels at 15, 30, and 60 minutes after insulin injection () and greater AUC (by 1.4-fold, ) compared with young mice, indicating the development of insulin resistance with advancing age. Non-fasting serum insulin levels in aged mice were slightly higher but not significantly different than in young mice (). In addition, we measured serum C-peptide concentrations, which are considered a better measure of secreted insulin than serum insulin levels. Non-fasting serum C-peptide levels were significantly higher (by 1.5-fold) in aged mice than in young mice (), consistent with increased insulin secretion. Accordingly, non-fasting blood glucose levels were significantly lower (1.2-fold) in aged mice than in young mice (). These results suggest that aged mice secrete more insulin than young mice to compensate for the increased insulin demands caused by glucose intolerance and insulin resistance.
Figure 1. Glucose tolerance and insulin sensitivity are impaired in aged mice. (A) Glucose tolerance tests. n = 14 and 10 for mice aged 3 and 24 months, respectively. (B) Area under curve (AUC) for data presented in (A). (C) Insulin tolerance tests. n = 14 and 9 for mice aged 3 and 24 months, respectively. (D) AUC for data presented in (C). (E) Non-fasting serum insulin concentrations quantified by ELISA. n = 16 for mice aged 3 and 24 months, respectively. (F) Non-fasting serum C-peptide concentrations measured by ELISA. n = 16 for mice aged 3 and 24 months, respectively. (G) Non-fasting blood glucose levels in young and aged mice. n = 14 and 9 for mice aged 3 and 24 months, respectively. * p < 0.05, ** p < 0.01, and *** p < 0.001 for comparisons between mice aged 3 and 24 months.
GAD67 expression is elevated in islets during aging and in response to high glucose levels
To determine if GAD67 levels are influenced by age, we performed RT-qPCR and immunofluorescence. We observed an approximate 3.2-fold increase in Gad67 mRNA levels in aged islets compared to in young islets (). Further, β-cells from an aged islet displayed significantly higher levels of GAD67 protein than β-cells from a young islet (). We performed ELISA and found that aged islets contained approximately 1.3-times more GAD67 protein than young islets (). In contrast, Gad65 mRNA and GAD65 protein levels in the glucagon-secreting α-cells of an islet showed no significant difference between young and aged islets ( and Supplementary Figure 1).
Figure 2. Aging and high glucose concentration upregulate GAD67 expression in primary islets. (A) Gad67 mRNA expression normalized to Actb analyzed by RT-qPCR. Data are from three independent experiments. (B) Representative confocal projection images of insulin (red) and GAD67 (green) protein expression. Projection = 20 μm. Scale bars = 30 μm. (C) The mean fluorescence intensity of GAD67-positive signals per islet area according to age. n = 26 and 111 islets from 3 mice of each age group. (D) Gad65 mRNA expression normalized to Actb in young and aged islets analyzed by RT-qPCR. Data are from three independent experiments. (E) Representative confocal projection images of young and aged islets stained for nuclei (DAPI, blue), insulin (red) and GAD65 (green). Projection = 20 μm. Scale bars = 30 μm. (F) The mean fluorescence intensity of GAD65-positive signals per islet area according to age. n = 99 and 292 islets from 3 mice of each age group. (G) Quantification of GAD67 protein content in islets by ELISA. n = 5 and 4 mice of each age group. (H) Glucose-induced GAD67 expression in young and aged islets visualized by confocal microscopy. Projection = 20 μm. Scale bars = 30 μm. (I) The mean fluorescence intensity of GAD67-positive signals per islet area. n = 151 and 165 islets from 3 mice aged 3 months, n = 158 and 93 islets from 3 mice aged 24 months from each condition.
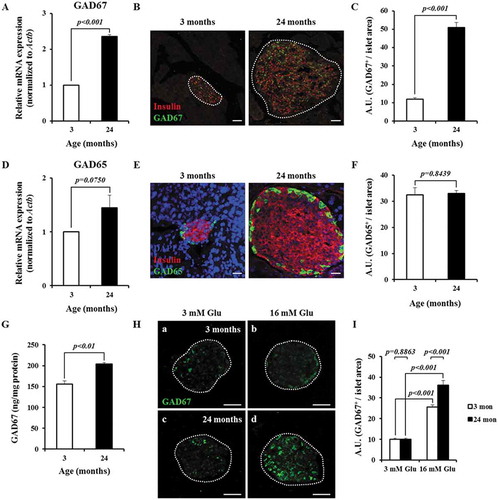
Next, we examined whether GAD67 expression is responsive to glucose, we incubated young and aged islets in 3 or 16 mM glucose for 48 h and analyzed GAD67 protein levels by immunofluorescence. GAD67 levels in young and aged islets cultured with 16 mM glucose were approximately 2.6- and 3.6-times greater than those in young and aged islets incubated in 3 mM glucose, respectively. Conversely, GAD65 levels in young and aged islets cultured with 16 mM glucose showed no significant difference compared to in young and aged islets incubated in 3 mM glucose (Supplementary Figure 2). These results suggest that elevated glucose concentration induces GAD67 expression in both young and aged islets, and that aged islets are more responsive than young islets ().
Elevated GAD67 expression does not cause autoimmune destruction in aged islets
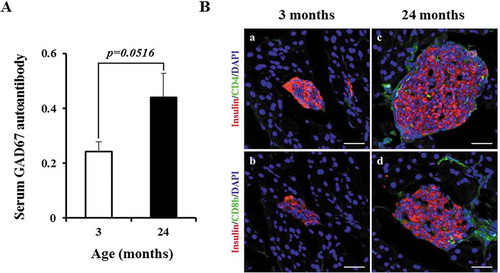
Given that GAD67 is a known islet autoantigen and biomarker for type 1 diabetes,Citation19,Citation20 we examined serum levels of the GAD67 autoantibody. As determined by ELISA, GAD67 autoantibody levels were about 1.8-fold higher in aged mice than in young mice (). We analyzed the entire pancreata by immunofluorescent staining for CD4 and CD8b molecules to determine whether islets had been infiltrated by T lymphocytes. We did not observe CD4+ or CD8b+ cells in representative confocal fluorescent images of young islets (Figure 3Ba and b). Although a few CD4+ or CD8b+ cells were present in aged islets (Figure 3Bc and d), they were rarely detected and islet architecture was intact. Together, these results suggest that elevated GAD67 expression does not appear to damage aged islets in nondiabetic C57BL/6J mice.
Figure 3. Aged islets do not exhibit β-cell destruction. (A) Serum levels of GAD67 IgG autoantibodies with aging. Y axis represents optical density values. n = 3 mice of each age group. (Ba–d) Representative confocal images of young and aged islets stained for nuclei (DAPI, blue), insulin (red), CD4 (green in a and c) and CD8b (green in b and d). n = 3 mice of each age group. Scale bars = 50 μm.
Inhibition of GAD67 activity abolishes glucose-stimulated insulin secretion (GSIS) in β-cells and young and aged islets
Because GAD67 expression is significantly upregulated in response to increased glucose concentrations, we examined whether GAD67 activity affects the capacity of β-cells and islets to secrete insulin. We treated the β-TC6 cell line with 3-mercaptopropionic acid (3-MPA), a potent competitive inhibitor of GADs that is commonly used to modify GAD activity in rodent pancreatic islets and brain,Citation21-Citation23 and did not observe effects on cell viability below 50 μM 3-MPA (Supplementary Figure 3A and B). Similarly, we tested primary islets isolated from young and aged mice. Treatment of primary islets with 1 mM 3-MPA, which inhibits >90% of GAD activity in islets,Citation23 did not affect cell viability (Supplementary Figure 3C-F). To verify that GAD activity was truly inhibited by 3-MPA, we measured GABA levels in β-TC6 cells and islets cultured with 25 mM glucose in the presence or absence of 3-MPA. Indeed, treatment with 3-MPA led to reduced GABA levels in β-TC6 cells, as well as young and old islets, consistent with inhibition of GAD67 activity ().
Figure 4. Suppression of GAD67 activity blocks GSIS from β-cells and young and aged islets. (A) Effect of 3-MPA (0, 10, and 50 μM) on GSIS from β-TC6 cells. (B) GSIS from 3 month old islets with or without 1 mM 3-MPA. n = 3 mice. (C) GSIS from 24 month old islets with or without 1 mM 3-MPA. n = 3 mice. (D) Comparison of GSIS indices determined in figure (A). (E) Comparison of GSIS indices measured in figure (B). (F) Comparison of GSIS indices measured in (C). Insulin concentrations were determined by ELISA. GSIS indices were calculated as the ratio of secreted insulin level stimulated with 25 mM glucose to that stimulated with 3 or 5 mM glucose. High (25 mM) glucose-induced secreted GABA concentration normalized by total protein levels from β-TC6 cells (G), young (H) and aged (I) islets in the absence or presence of 3-MPA. Three independent experiments for β-TC6 cells and three mice of each age group were included.
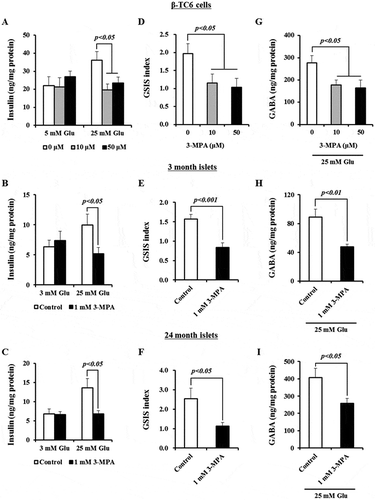
We observed increased insulin secretion in β-TC6 cells, as well as in young and aged primary islets, cultured with high vs. low glucose concentrations, but this increase was abolished in the presence of 3-MPA (). Further, the GSIS index of β-TC6 cell lines, and of young and aged islets, cultured with 25 mM glucose declined upon the addition of 3-MPA (). Together, these results indicate that blocking GAD67 activity restrains GSIS from β-cells as well as young and aged islets.
Discussion
Here, we show that apparently healthy, non-diabetic C57BL/6J mice develop impaired glucose metabolism and insulin resistance during aging. Age-associated enlargement in islet size along with increased numbers of insulin-positive β-cells may represent distinct features of compensatory islets that develop in response to impaired glucose tolerance and elevated insulin resistance, as seen in obese humans and rodents, where β-cell replication is the major mechanism underlying the mass expansion of islets.Citation24,Citation25 Pancreatic β-cells in 2-year-old mice retain their capacity for compensatory replication, despite a decline in the basal proliferation rate.Citation26 Our previous report showed that the intrinsic ability of islet β-cells to produce and release insulin does not decline with age in mice and humans.Citation4 In agreement with β-cells being relatively competent in aged islets, serum insulin and C-peptide levels in aged mice were higher than those in young mice. Aging-dependent gene expression and functional changes in mouse and human β-cells were previously investigated. While several transcription factors associated with β-cell function such as Pdx1, Nkx6-1, and Foxa2 were further activated in aged mouse β-cells, but not in aged human β-cells, the tumor suppressor protein p16InkCitation4a (encoded by Cdkn2a) revealed the elevated gene expression in both aged mouse and human β-cells.Citation27 It was also reported that p16InkCitation4a-induced senescence in mouse and human β-cells promotes GSIS.Citation28 Based on these reports, they suggest that many factors can affect β-cell function with age.
In this study, we found that Gad67 mRNA and GAD67 protein levels in mouse pancreatic islets increased with advancing age. Importantly, elevated glucose concentration lead to increased GAD67 protein levels in islets isolated from young and aged mice. A previous study reported that glucose enhances GAD67 expression in cultured rat islets.Citation29 Given that upregulation of GAD67 in aged islets is an adaptation to higher glucose, which can be reversed after incubation with 3 mM glucose for 48 h, GAD67 would be lower in aged islets than young islets in the context of lower fed glycemia. Conversely, we observed that GAD67 in aged islets is even higher than that in young islets. It was reported that GAD67 is regulated by PDX1, an insulin transcription factor.Citation30 Therefore, we postulated that the glucose-derived autocrine insulin/AKT/FOXO1/PDX1 signaling pathway might contribute to the elevated levels of GAD67 in islets in response to aging and/or glucose.Citation31-Citation33 GAD67 inhibition did not affect basal insulin secretion from the cultured β-TC6 cell line and young and aged islets, but blocked GSIS. The selectivity of 3-MPA is not specific to GAD67 inhibition and it also reduces fatty acid oxidation by inhibiting long- and medium-chain acyl-CoA dehydrogenase.Citation35 Glucose-derived malonyl-CoA increases cytosolic long-chain (LC) acyl-CoA via inhibition of fatty acid oxidation. Malonyl-CoA and LC acyl-CoA esters are metabolic coupling factors in glucose- and nutrient-induced insulin secretion from the hamster pancreatic HIT-T15 β-cells.Citation36,Citation37 Therefore, under the condition where 3-MPA blocks fatty acid oxidation, increased cytosolic LC acyl-CoA would enhance GSIS. However, we observed a significant decrease in GSIS from both young and aged islets treated with 3-MPA (), suggesting that the decline of GSIS may not result solely from reduced fatty acid oxidation by 3-MPA.
To further assess if GAD67 inhibition using 3-MPA affects insulin granule exocytosis, we performed KCl (30 mM)-induced insulin secretion (KIIS) with β-TC6 cells and young islets. As a result, KIIS in β-TC6 cells and young islets was significantly inhibited by 3-MPA (Supplementary Figure 4), suggesting that inhibition of GSIS in β-cells and young and aged islets by 3-MPA is attributed to the disturbance of insulin granule exocytosis. On the contrary, overexpression of GAD65 in miceCitation38 and INS-1E β-cells and rat isletsCitation39 deteriorates GSIS. Overexpression of GAD65 in β-cells might cause increased trafficking of cytosolic GAD67:GAD65 heterodimers to Golgi membranes or peripheral vesicles, via the membrane anchoring or palmitoylation/depalmitoylation cycle of GAD65,Citation40 which would reduce cytosolic levels of GAD67 and, in turn, GSIS. Taken together, these findings suggest that GAD67 has a role in compensatory insulin secretion during aging, probably through regulation of cellular glutamate or GABA contentCitation38,Citation39 or insulin granule exocytosis, although the molecular mechanism underlying modulation of GSIS remains to be determined.
We acknowledge that there are limitations of this study. First, we did not use the different genetic background of mice and only studied male mice, in which GAD67 is the major isomer, whereas GAD65 is preferentially expressed in human islets. Second, dynamics of insulin secretion and CaCitation2+ measurements by islet perifusion would provide further insights of the roles of GAD67/3-MPA in GSIS. Nonetheless, our results demonstrated that GAD67 levels increase in islets with aging and also indicated an important functional role for GAD67 in compensatory insulin secretion from aged islets. Therefore, the results of this study provide new insight into blood glucose regulation during aging and, more specifically, how the escalation of GAD67 in aged islets contributes to the systemic glucose intolerance and insulin resistance that occur with aging.
Materials and methods
Pancreatic β-cell line culture
The β-TC6 mouse insulinoma cells (ATCC, CRL-11506) were cultured in ATCC-formulated Dulbecco’s modified Eagle’s medium (DMEM) containing 4 mM L-glutamine and 25 mM glucose in an atmosphere of 5% CO2, 95% air at 37°C.Citation41 The medium was supplemented with 15% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin. All experiments were performed using β-TC6 cells between passages 35 and 42.
Animals and animal care
Male C57BL/6J mice were from KRIBB-IACUC and housed in a specific pathogen-free animal facility (22°C ± 3°C; 50% ± 10% relative humidity). All animal care and surgical procedures were approved by the Animal Care and Use Committee of DGIST.
Primary islet isolation and maintenance
Pancreatic islets were isolated from the pancreata of young (3 month old) and aged (24 month old) mice following standard islet isolation procedures.Citation42 Mice were anesthetized and then sacrificed by exsanguination. Collagenase type V (Sigma, C9263) was dissolved in Hank’s balanced salts solution to a 1 mg/ml concentration and then was transfused into the common bile duct. The partially digested pancreas was excised and incubated for 10–15 min at 37°C in a water bath with shaking every 3 min. Islets were picked under a stereoscope and maintained in RPMI-1640 medium supplemented with 10% FBS, L-glutamine, 5 mM glucose, penicillin (100 units/ml), and streptomycin (0.1 mg/ml) at 37°C in a 5% CO2 incubator overnight.
Intraperitoneal glucose and insulin tolerance tests
Intraperitoneal glucose and insulin tolerance tests were performed as described previously.Citation4 Briefly, for glucose and insulin tolerance tests, mice were fasted overnight (16 h) and for 6 h with free access to water, respectively. Mice were injected intraperitoneally with glucose (1 g/kg body weight) and insulin (Sigma, I9278; 0.75 U/kg body weight). Blood glucose levels were measured at 0, 30, 60, 90, and 120 min for glucose tolerance tests and at 0, 15, 30, 60, and 90 min for insulin tolerance tests from a cut tail vein using an Accu-Chek® Nano blood glucose meter (Roche).
Glucose-stimulated insulin secretion (GSIS) tests
β-TC6 cells were seeded in a 12-well plate (2 × 10Citation5 cells/well) in triplicate and incubated for 48 h. Cells were incubated with DMEM containing 1% FBS, 5.6 mM glucose, and different doses of 3-MPA (0, 10, and 50 μM) for 24 h, washed once with Krebs-Ringer Bicarbonate Buffer (KRBB; 114 mM NaCl, 4.4 mM KCl, 1 mM MgSO4, 29.5 mM NaHCO3, 1.28 mM CaCl2, 0.1% BSA, and 10 mM HEPES, adjusted to pH 7.4) without glucose, starved with KRBB plus 5 mM glucose and different doses of 3-MPA for 5 h, washed once with KRBB without glucose, and stimulated with KRBB (1 ml per well) containing 5 mM and 25 mM glucose and 30 mM KCl for 1 h. Supernatants were then collected for insulin measurement. For islet experiments, five size-matched islets per well were seeded in triplicate in a 24-well plate in KRBB. Mouse islets were starved for 5 h at 37ºC in KRBB without glucose (400 μl per well) and incubated for 1 h at 37ºC with 25 mM glucose or 30 mM KCl and 1 mM 3-MPA (Sigma, M5801) dissolved in DMSO for each condition. All supernatant samples (400 μl per well) were stored at −80ºC before measurement. Secreted insulin levels from β-TC6 cells and young and aged islets were normalized by their total protein levels.
Assessment of 3-MPA toxicity
Prior to examining the effect of 3-MPA on β-cells or islet function, cell viability assays were performed to identify 3-MPA concentrations that had minimal effects on cell viability. The β-TC6 cell line and young and aged islets were maintained in the absence and presence of 3-MPA, as described above. The concentrations of 3-MPA used were 10, 50, and 200 μM for β-TC6 cells (Supplementary Figure 3A and B); 1 mM for young (Supplementary Figure 3C and D) and aged islets (Supplementary Figure 3E and F). Live/dead cells were stained using LIVE/DEAD® kit (Thermo Fisher Scientific, L3224) containing 2 μM calcein AM and 4 μM EthD-1 at room temperature for 30 min. Images were taken using a Leica DMI 3000B fluorescence microscope for β-TC6 cells, and with an Olympus FV1000 confocal laser scanning microscope with 50 μm projection for young and aged islets. Cell viability (%) was calculated as the relative percentage of the total area of live cells divided by the total number of dead cells in a field of view: β-TC6 cells, n = 6 wells, total 12 images per condition; young islets, n = 257 and 263 islets per condition from 3 mice; and aged islets, n = 245 and 241 islets per condition from 2 mice).
Quantitative reverse transcription real-time PCR (RT-qPCR)
Total RNA (5.6–11.7 μg) was extracted from approximately 1000 islets using a Hybrid-RTM RNA Purification Kit (GeneAll, 305–101). cDNA was synthesized using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, 4368814) from 2 μg total RNA following the manufacturer’s instructions. Primers were designed using Primer Express Software for Real-Time PCR Version 3.0. Quantitative real time PCR was performed using an ABI 7500 Real Time PCR System and the SYBRTM Green PCR Master Mix (Applied Biosystems, 4309155). Amplification conditions were 2 min at 50°C, 10 min at 95°C, 1 min at 60°C; followed by 40 cycles of 15 s at 95°C, 1 min at 60°C, 30 s at 95°C, and 15 s at 60°C. Reaction specificity was confirmed by melting curve analysis.Citation41 Primers used in this study were shown in Supplementary Table 1. Actb was used as an internal standard.
Measurement of insulin, C-peptide, GAD67 antigen, GAD67 autoantibody and GABA concentrations by ELISA
ELISA was performed using commercial ELISA kits shown in Supplementary Table 2, and experiments were conducted according to the manufacturer’s instructions. All cell lysates were prepared using RIPA buffer (Thermo Fisher Scientific, NCI9900KR) containing HaltTM Protease Inhibitor Single-Use Cocktail (Thermo Fisher Scientific, 1860932) and HaltTM Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific, 78426) and their total protein levels were measured using PierceTM BCA Protein Assay Kit (Thermo Fisher Scientific, 23225) according to the manufacturer’s instructions.
Immunohistochemistry
Pancreata from young (3 months old) and aged (24 months old) mice were fixed in 4% paraformaldehyde overnight, incubated overnight in a sucrose gradient (10, 20, and 30% sucrose in PBS, respectively), and frozen in Tissue-Tek Optimal Cutting Temperature (OCT) compound (Sakura Finetek, 4583) before cryosectioning (−20°C). Pancreas sections (20 μm) were rinsed with PBS-Triton X-100 (0.3%) and incubated in blocking solution (PBS-Triton X-100 and CAS-BlockTM (Thermo Fisher Scientific, 00–8120)). Thereafter, sections were immunostained with primary antibodies diluted in blocking solution for 48 h at RT and with secondary antibodies diluted in PBS for 16 h at RT.Citation4 The antibodies used for immunohistochemistry were shown in Supplementary Table 3. Cell nuclei were labeled with DAPI. Slides were mounted with ProLongTM Gold Antifade Mountant (Thermo Fisher Scientific, P36934). All projected confocal images (20 μm) of islets were taken using an Olympus FV1000 confocal laser scanning microscope. Images were analyzed using Image J software.
Measurement of glucose-induced GAD65 and GAD67 expression
Young (3 months old) and aged (24 months old) islets were incubated with RPMI-1640 medium containing 10% FBS and 3 or 16 mM glucose for 48 h. Collected islets were fixed in 4% paraformaldehyde overnight, washed in PBS once, seeded in 1.5% agarose gel, and frozen in Tissue-Tek OCT compound before cryosectioning (−20°C). After permeabilization and blocking processes as described above, islet sections (20 μm) were immunostained using anti-GAD65 and anti-GAD67 antibodies and GAD65 and GAD67 content was quantified by immunohistochemistry.
Statistical analysis
All data are presented as mean ± SEM obtained from at least three independent experiments. Statistical significance was analyzed by unpaired or paired Student t-test (Excel, Microsoft). P-values of < 0.05 were considered statistically significant.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
Supplemental Material
Download MS Word (3.8 MB)Acknowledgments
We thank Ms. Su Hyun Jung and Hye Soo Kim for their help with confocal microscopy. We also thank Drs. Joana Almaça and Seong-Kyoon Choi for their comments regarding the experimental methods.
Supplementary material
Supplemental data for this article can be accessed on the publisher’s website.
Additional information
Funding
References
- Chang AM, Halter JB. Aging and insulin secretion. Am J Physiol Endocrinol Metab. 2003;284:E7–12. doi:10.1152/ajpendo.00366.2002.
- Defronzo RA. Glucose intolerance and aging: Evidence for tissue insensitivity to insulin. Diabetes. 1979;28:1095–1101.
- Jackson RA. Mechanisms of age-related glucose-intolerance. Diabetes Care. 1990;13(Suppl 2):9–19. doi:10.2337/diacare.13.2.S9.
- Almaça J, Molina J, Arrojo E Drigo R, Abdulreda MH, Jeon WB, Berggren P-O, Caicedo A, Nam HG. Young capillary vessels rejuvenate aged pancreatic islets. Proc Natl Acad Sci U S A. 2014;111:17612–17617. doi:10.1073/pnas.1414053111.
- Okada T, Liew CW, Hu J, Hinault C, Michael MD, Krtzfeldt J, Yin C, Holzenberger M, Stoffel M, Kulkarni RN. Insulin receptors in β-cells are critical for islet compensatory growth response to insulin resistance. Proc Natl Acad Sci U S A 2007; 104:8977–8982.
- Erdö SL, Wolff JR. γ-Aminobutyric acid outside the mammalian brain. J Neurochem. 1990;54:363–372.
- Kim J, Richter W, Aanstoot HJ, Shi Y, Fu Q, Rajotte R, Warnock G, Baekkeskov S. Differential expression of GAD65 and GAD67 in human, rat, and mouse pancreatic islets. Diabetes. 1993;42:1799–1808.
- Reetz A, Solimena M, Matteoli M, Folli F, Takei K, De Camilli P. GABA and pancreatic beta-cells: colocalization of glutamic acid decarboxylase (GAD) and GABA with synaptic-like microvesicles suggests their role in GABA storage and secretion. EMBO J. 1991;10:1275–1284. doi:10.1002/j.1460-2075.1991.tb08069.x.
- Christgau S, Schierbeck H, Aanstoot HJ, Aagaard L, Begley K, Kofod H, Hejnaes K, Baekkeskov S. Pancreatic β cells express two autoantigenic forms of glutamic acid decarboxylase, a 65-kDa hydrophilic form and a 64-kDa amphiphilic form which can be both membrane-bound and soluble. J Biol Chem. 1991;266:21257–21264.
- Christgau S, Annstoot HJ, Schierbeck H, Begely K, Tullin S, Hejnaes K, Baekkeskov S. Membrane anchoring of the autoantigen GAD65 to microvesicles in pancreatic β-cells by palmitoylation in the NH2-terminal domain. J Cell Biol. 1992;118:309–320. doi:10.1083/jcb.118.2.309.
- Baekkeskov S, Aanstoot HJ, Christgau S, Reetz A, Solimena M, Cascalho M, Folli F, Richter-Olesen H, De Camilli P. Identification of the 64K autoantigen in insulin-dependent diabetes as the GABA-synthesizing enzyme glutamic acid decarboxylase. Nature. 1990;347:151–156. doi:10.1038/347151a0.
- Kash SF, Johnson RS, Tecott LH, Noebels JL, Mayfield RD, Hanahan D, Baekkeskov S. Epilepsy in mice deficient in the 65-kDa isoform of glutamic acid decarboxylase. Proc Natl Acad Sci U S A. 1997;94:14060–14065. doi:10.1073/pnas.94.25.14060.
- Asada H, Kawamura Y, Maruyama K, Kume H, Ding RG, Kanbara N, Kuzume H, Sanbo M, Yagi T, Obata K. Cleft palate and decreased brain γ-aminobutyric acid in mice lacking the 67-kDa isoform of glutamic acid decarboxylase. Proc Natl Acad Sci U S A. 1997;94:6496–6499. doi:10.1073/pnas.94.12.6496.
- Soltani N, Qiu H, Aleksic M, Glinka Y, Zhao F, Liu R, Li Y, Zhang N, Chakrabarti R, Ng T, et al. GABA exerts protective and regenerative effects on islet beta cells and reverses diabetes. Proc Natl Acad Sci U S A. 2011;108:11692–11697. doi:10.1073/pnas.1102715108.
- Purwana I, Zheng J, Li X, Deurloo M, Son DO, Zhang Z, Liang C, Shen E, Tadkase A, Feng ZP, et al. GABA promotes human β-cell proliferation and modulates glucose homeostasis. Diabetes. 2014;63:4197–4205. doi:10.2337/db14-0153.
- Pleau JM, Esling A, Van Acker C, Dardenne M. Glutamic acid decarboxylase (GAD 67) gene expression in the pancreas and brain of the nonobese diabetic mouse. Biochem Biophys Res Commun 1996; 224:747–753.
- Pleau JM, Throsby M, Esling A, Dardenne M. Ontogeny of glutamic acid decarboxylase gene expression in the mouse pancreas. Biochem Biophys Res Commun 1997; 233:227–230.
- Feng MM, Xiang YY, Wang S, Lu WY. An autocrine γ-aminobutyric acid signaling system exists in pancreatic β-cell progenitors of fetal and postnatal mice. Int J Physiol Pathophysiol Pharmacol. 2013;5:91–101.
- Honeyman MC, Cram DS, Harrison LC. Glutamic acid decarboxylase 67–reactive T cells: a marker of insulin-dependent diabetes. J Exp Med. 1993;177:535–540. doi:10.1084/jem.177.2.535.
- Elliott JF, Qin HY, Bhatti S, Smith DK, Singh RK, 605 Dillon T, Lauzon J, Singh B. Immunization with the larger isoform of mouse glutamic acid decarboxylase (GAD67) prevents autoimmune diabetes in NOD mice. Diabetes. 1994;43:1494–1499. doi:10.2337/diab.43.12.1494.
- Katoh J, Taniguchi H, Ogura M, Kasuga M, Okada Y. A convulsant, 3-mercaptopropionic acid, decreases the level of GABA and GAD in rat pancreatic islets and brain. Experientia. 1995;51:217–219. doi:10.1007/BF01931099.
- Okada Y, Taniguchi H. Effect of glutamate decarboxylase- and GABA transaminase-inhibitors on the GABA (γ-aminobutyric acid) level in the isolated pancreatic islets and the hippocampal slice of the rat in vitro. Proc Japan Acad Ser B. 1980;56:46–50. doi:10.2183/pjab.56.46.
- Fernandèz-Pascual S, Mukala-Nsengu-Tshibangu A, Martín-Del-Río R, Tamarit-Rodriguez J. Conversion into GABA (γ-aminobutyric acid) may reduce the capacity of L-glutamine as an insulin secretagogue. Biochem J. 2004;379:721–729. doi:10.1042/bj20031826.
- Saisho Y, Butler AE, Manesso E, Elashoff D, Rizza RA, Butler PC. β-Cell mass and turnover in humans: effects of obesity and aging. Diabetes Care. 2013;36:111–117. doi:10.2337/dc12-0421.
- Roat R, Rao V, Doliba NM, Matschinsky FM, Tobias JW, Garcia E, Ahima RS, Imai Y. Alterations of pancreatic islet structure, metabolism and gene expression in diet-induced obese C57BL/6J mice. PLoS One. 2014;9:e86815. doi:10.1371/journal.pone.0086815.
- Stolovich-Rain M, Hija A, Grimsby J, Glaser B, Dor Y. Pancreatic beta cells in very old mice retain capacity for compensatory proliferation. J Biol Chem. 2012;287:27407–27414. doi:10.1074/jbc.M112.350736.
- Avrahami D, Li C, Zhang J, Schug J, Avrahami R, Rao S, Stadler MB, Burger L, Schübeler D, Glaser B, et al. Aging-dependent demethylation of regulatory elements correlates with chromatin state and improved β cell function. Cell Metab. 2015;22:619–632. doi:10.1016/j.cmet.2015.07.025.
- Helman A, Klochendler A, Azazmeh N, Gabai Y, Horwitz E, Anzi S, Swisa A, Condiotti R, Granit RZ, Nevo Y, et al. p16 Ink4a-induced senescence of pancreatic beta cells enhances insulin secretion. Nat Med. 2016;22:412–420. doi:10.1038/nm.4054.
- Pedersen AA, Videbaek N, Skak K, Petersen HV, Michelsen BK. Characterization of the rat GAD67 gene promoter reveals elements important for basal transcription and glucose responsiveness. DNA Seq. 2001;11:485–499. doi:10.3109/10425170109041332.
- Pedersen AA, Petersen HV, Videbaek N, Skak K, Michelsen BK. PDX-1 mediates glucose responsiveness of GAD67, but not GAD65, gene transcription in islets of Langerhans. Biochem Biophys Res Commun. 2002;295:243–248. doi:10.1016/S0006-291X(02)00674-5.
- Ohsugi M, Cras-Méneur C, Zhou Y, Warren W, Bernal-Mizrachi E, Permutt MA. Glucose and insulin treatment of insulinoma cells results in transcriptional regulation of a common set of genes. Diabetes. 2004;53:1496–1508.
- Kitamura T, Nakae J, Kitamura Y, Kido Y, Biggs WH, Wright CVE, White MF, Arden KC, Accili D. The forkhead transcription factor Foxo1 links insulin signaling to Pdx1 regulation of pancreatic β cell growth. J Clin Invest. 2002;110:1839–1847. doi:10.1172/JCI200216857.
- Martinez SC, Cras-Méneur C, Bernal-Mizrachi E, Permutt MA. Glucose regulates Foxo1 through insulin receptor signaling in the pancreatic islet β-cell. Diabetes. 2006;55:1581–1591. doi:10.2337/db05-0678.
- Humphrey RK, Yu SM, Flores LE, Jhala US. Glucose regulates steady-state levels of PDX1 via the reciprocal actions of GSK3 and AKT kinases. J Biol Chem. 2010;285:3406–3416. doi:10.1074/jbc.M109.006734.
- Sabbagh E, Cuebas D, Schulz H. 3-Mercaptopropionic acid, a potent inhibitor of fatty acid oxidation in rat heart mitochondria. J Biol Chem. 1985;260:7337–7342.
- Martens GA, Vervoort A, Van De Casteele M, Stangé G, Hellemans K, Van Thi HV, Schuit F, Pipeleers D. Specificity in beta cell expression of L-3-hydroxyacyl-CoA dehydrogenase, short chain, and potential role in down-regulating insulin release. J Biol Chem. 2007;282:21134–21144. doi:10.1074/jbc.M700083200.
- Prentki M, Vischer S, Glennon MC, Regazzi R, Deeney JT, Corkey BE. Malonyl-CoA and long chain acyl-CoA esters as metabolic coupling factors in nutrient-induced insulin secretion. J Biol Chem. 1992;267:5802–5810.
- Shi Y, Kanaani J, Menard-Rose V, Ma YH, Chang PY, Hanahan D, Tobin A, Grodsky GM, Baekkeskov S. Increased expression of GAD65 and GABA in pancreatic beta-cells impairs first-phase insulin secretion. Am J Physiol Endocrinol Metab. 2000;279:E684–94. doi:10.1152/ajpendo.2000.279.3.E684.
- Rubi B, Ishihara H, Hegardt FG, Wollheim CB, Maechler P. GAD65-mediated glutamate decarboxylation reduces glucose-stimulated insulin secretion in pancreatic beta cells. J Biol Chem. 2001;276:36391–36396. doi:10.1074/jbc.M104999200.
- Kanaani J, Cianciaruso C, Phelps EA, Pasquier M, Brioudes E, Billestrup N, Baekkeskov S. Compartmentalization of GABA synthesis by GAD67 differs between pancreatic beta cells and neurons. PLoS One. 2015;10:e0117130. doi:10.1371/journal.pone.0117130.
- Lee KM, Jung GS, Park JK, Choi SK, Jeon WB. Effects of Arg-Gly-Asp-modified elastin-like polypeptide on pseudoislet formation via up-regulation of cell adhesion molecules and extracellular matrix proteins. Acta Biomater.2013;9:5600-5608. doi: 10.1016/j.actbio.2012.10.038.
- Li DS, Yuan YH, Tu HJ, Liang QL, Dai LJ. A protocol for islet isolation from mouse pancreas. Nat Protoc. 2009;4:1649–1652. doi: 10.1038/nprot.2009.150.

