Abstract
Recent Government announcements have implied that wind power will play a major part in providing energy for the UK (BBC Citation2007). However, there is much concern that wind farms can have a significant impact on flying fauna (bats and birds) using the area, particularly at night. As part of an Environmental Impact Assessment, thorough appropriate surveys are necessary for quantifying and minimising any risk wind farms may cause flying fauna. Manual surveys that are commonly used are not always cost‐effective, efficient or practical. Remote systems based on motion detection are increasingly being used to monitor wildlife.
Fast‐moving airborne targets such as aeroplanes can falsely trigger motion‐detection based remote systems. As birds and bats repetitively flap their wings, this oscillating motion can be used to distinguish them from other airborne targets. Time periods between wing oscillations are not always constant, and hence the motion is not periodic. A method to detect cyclic motion based on similarity matrices is proposed, and synthetic and real data are used.
1. Background
Renewable technologies, such as those using wind power, may help meet the United Nations Environment Programme aims of sustainability (UNEP Citation2006). As a mature technology, renewable energy from wind is considered as one of the leading tools for tackling carbon emissions. Recently, the British Government has announced its intention to develop and install more wind farms to meet the demand for lower carbon generated energy (BBC Citation2007). This demand is likely to be met with offshore wind farms, with the UK expected to be the largest generator of offshore wind power worldwide in 2008 (BWEA Citation2007).
Offshore wind farms are likely to be one of the largest man‐made interferences in the seas around Europe (Exo et al. Citation2003). Onshore wind farms also pose problems, with poorly located wind turbines potentially causing collision risks, or harming and reducing habitat (RSPB Citation2005). Also infrastructure associated with wind farms can have a devastating effect (e.g. Kuvlesky et al. Citation2007).
To try and reduce the unnecessary impacts of structures such as wind farms on birds and bats, Environmental Impact Assessments (EIA), as part of a European Union directive, are required to be conducted during the proposal stage (DCLG Citation2000). As part of the EIA, natural heritage likely to be affected by a schedule one development (see Town and Country Planning regulations 1999) are identified. Identified natural heritage at risk have to then be assessed for potential impacts. For birds and bats, relevant surveys may be carried out to collect information on area activity, species numbers, area utilisation, etc. This data are then analysed by specialists who can provide the developer with information concerning the risk to the environment (DCLG Citation2000).
Research shows that many bird species, such as ducks, geese and waders are active at night as well as during the day. Bird groups such as passerines (e.g. Schmaljohann et al. Citation2007), waders (e.g. Gudmundsson Citation1994), geese (e.g. Alerstam et al. Citation1993) and ducks (e.g. Flock Citation1973) migrate at night. In addition to migrating and moving to roosts, birds may be engaged in other activities. For example, wader species such as Lapwing Vanellus vanellus feed at night to balance their energy budgets (Sheldon et al. Citation2004). Redshanks Tringa totanus have been found to forage for food more at night than during the day (Burton and Armitage Citation2005), and other shorebird species have been observed foraging at night (McNeil and Roberts 1992). As nocturnal behaviour may not always match diurnal behaviour (Gillings et al. Citation2005) nocturnal surveys are vital to supplement information regarding species behaviour at a potential development site.
Bats are also at threat from poorly located wind turbines. A number of causes of bat mortality at wind turbines have been speculated including migration, high altitude foraging flights, interference with ultrasound and wind turbine wind‐shear (Williams Citation2004, Bets 2007). Bats must be considered as part of an EIA, and appropriate surveys carried out (Betts Citation2007). Current bat detection techniques are based on bat detectors that do not provide the surveyor with a visual description of bat activity. The use of night vision equipment can help supplement understanding of bat activity.
Surveys of species at risk from developments are essential to the preservation of the natural environment, aiding sustainable development.
1.1 Current visual‐based methods
Manual‐based surveys are commonly used during nocturnal surveys of a site. This usually involves an experienced ornithologist sampling for bird activity at different locations on the site with the aid of night vision technology, such as a light intensifier or thermal imager (e.g. Desholm et al. Citation2004). This is not always practical, if a particular location requires continuous monitoring or the potential site is hazardous or difficult to survey, such as an offshore wind farm.
Remote systems provide a realistic alternative to manual surveys by storing data for later analysis. Remote systems can reduce the disturbance of the survey on fauna, provide a back up of data for review and reduces the number of personnel required on site (Sykes et al. Citation1995). Post‐survey analysis can be a lengthy, drawn out process. It requires personnel to watch through potentially large periods of inactivity until a target is spotted.
To improve the efficiency of remote systems, the use of motion or trigger detection could be used to remove the majority of periods with no target activity. This approach is becoming popular with remote monitoring, with two systems WT‐Bird and Thermal Animal Detection System (TADS), developed with wind farm monitoring in mind, operating on this principle (Desholm Citation2003, Wiggelinkhuizen et al. Citation2006).
Basic motion and trigger detection mechanisms are liable to start recording when the target of interest is not present. For example, fast moving clouds, aeroplanes and trees shaking in the wind can cause motion, as well as birds and bats flying past. We propose approaches below to improve the detection of birds and bats, ignoring other objects that may cause motion detection.
1.2 Detecting wing beat characteristics
Birds and bats fly by flapping their wings, making them distinguishable from other airborne objects. By detecting this exclusive characteristic, other objects that cause false motion detection can be ignored. Benefits include speedier analysis of video data or storage space savings, thereby increasing the practicality of remote surveys, and nocturnal ecological surveys in general.
Wing beat frequencies of fauna are of interest from a number of research viewpoints. Such examples include wing beat frequencies used as a method for calculating energy expenditure (e.g. Hambly et al. Citation2004, Engel et al. Citation2006, Schmidt‐Wellenburg et al. Citation2007, etc), bird identification in radar (e.g. Houghton and Blackwell Citation1972, Zhang et al. Citation2005,), and biomechanics (e.g. Norberg Citation2002).
The flapping motion exhibited by birds and bats can be described as either periodic; flapping motion repeats over time (i.e. temporally) with a fixed time period between flaps, or cyclic; flapping motion repeats temporally, but the duration between flaps varies. This non‐uniformity regarding target flight can occur for many reasons. The target fauna may be soaring (wings outstretched), taking off in flight, avoiding an obstacle or merely settling into flight (Tobalske Citation2007). Flapping patterns can also be unique to bird groups or species (Cornell Lab of Ornithology Citation2007).
Another advantage of being able to detect wing beat frequencies is that a single camera can be used. Stereo cameras can provide depth information, but can be affected by a number of limitations related to monitoring distant bird targets. These include errors involved in target correspondence from both images, either through target miss‐matching, quantisation and target distortion (Mohan et al. Citation1989, Rodriguez and Aggarwal Citation1990), and the very small disparity associated with distant targets (Shah Citation1997, Munoz‐Salinas et al. Citation2008). As bird targets can be a few pixels in size, the further away the bird target is, the greater the error in distance calculated from stereo cameras is likely to be. Wing beat patterns are constant irrespective of relative speed or distance, and may be a more robust method for identifying bird targets that are at distance away from the camera.
There are a number of periodicity and cyclic motion detection methods available. The main types, as classified by Cutler and Davis (Citation2000) are:
point correspondence analysis; | |||||
pixel periodicity analysis; | |||||
periodic motion feature analysis; | |||||
object similarity periodicity analysis; | |||||
moving object rigidity analysis. | |||||
An overview of the techniques found in these categories above can be found in Lazarevic et al. (Citation2007). The main review focus of the paper is on object similarity periodicity analysis based on similarity matrices and Seitz and Dyers' (Citation1997) contribution to point correspondence analysis.
Seitz and Dyer (Citation1997) define cyclic motion by removing the temporal constraint of periodic motion, and consider motion cyclic if it repeats, regardless of the time taken to repeat the motion. This is defined as:
Cutler and Davis (Citation2000) check objects for periodicity by firstly aligning the objects by their tracking results. The aligned objects are then differenced against each other. The absolute values found are added to a similarity matrix. The closer the value is to zero, the more likely the frames are similar to each other. Periodic motion is detected by checking one‐dimensional (1D) power spectral density using
Plotnik and Rock (Citation2002) also use a similarity matrix, but values in the matrix are calculated using a normalised sum of squared differences between each frame. As with Cutler and Davis, periodic motion is detected using Equation Equation(2), and a target is identified if the autocorrelation of the similarity matrix matches a pre‐defined lattice.
Branzan‐Albu et al. (Citation2005) is also based on Cutler and Davis (Citation2000), but values for the similarity matrix are calculated by correlating objects with each other, and the peak correlation value used in the matrix. A global threshold is applied to the matrix, followed by a number of standard image processing morphological operators. These processes reduce the similarity matrix to a pattern based around the identity diagonal.
A key advantage of object similarity periodicity analysis based on similarity matrices, such as the methods outlined above, is that low‐resolution images that may be blurry can be used (Cutler and Davis Citation2000). This reduces the importance of observable features such as wing size or bird shape, as the overall change of the target is compared over time, and not specific areas.
The similarity matrix based methods outlined above tend to identify a change from one type of periodic motion to another, for example, from walking to running. This approach fails to detect cyclic motion occurring within the motion. Another potential issue arises based on the number of object frames available when flying fauna cross the camera viewshed. Many flying fauna targets cross the camera viewshed within a second (pers. obs.). Considering typical video frame rates of 25 frames per second (Phase Alternating Line), the potential number of target images available is low. There may not be enough frames for lattice matching techniques used by Cutler and Davis (Citation2000) and Plotnik and Rock (Citation2002), and the morphological operations used by Branzan‐Albu et al. (2006) may cause most of the similarity matrix information to be lost.
2. Proposed method
The methods outlined above all use similarity matrices as part of the process for finding periodicity based on spectral frequency analysis. It is proposed that the similarity matrix can be used to identify cyclic motion based on the pattern formed by the matrix.
A similarity matrix is a way of determining relationships between signals, or a single signal (self‐similarity). This method can provide a visual or numeric representation of correspondence within the signal, displaying a relationship between signals or within a signal. Applications for similarity matrices include genetic (e.g. Borowski et al. Citation2000) and music analysis (e.g. Muller and Kurth Citation2006). An illustration of how a similarity matrix can be used is presented as follows.
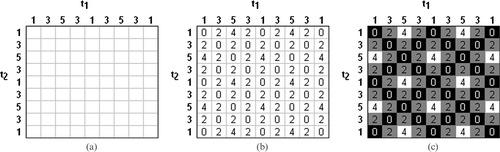
An oscillating numerical series of {1, 3, 5, 3, 1, 3, 5, 3, 1} is plotted against itself in a similarity matrix (Figure ). The result of the difference for each term over time is placed in each element (Figure ). The lower the value, the more similar the terms of the series are, and the larger the value, the more different they are. A pattern within the matrix suggests a periodic or cyclic relationship is present within the series. Finally, providing a shading reference to the elements, using black to represent zero and white to represent the maximum difference (four in this instance), a visualisation of the pattern is presented (Figure ). Measuring the number of elements between crossover peaks of similarity (black elements) identifies how often the series repeats (every four elements in this instance).
Figure 1 Similarity matrix illustration using a number series.(a) The series plotted against itself. (b) Series terms differenced over time. (c) Colour‐shading of elements to provide a visual interpretation.
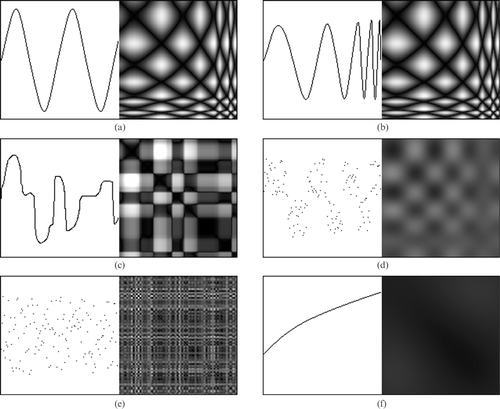
A number of scenarios have been produced to represent different types of periodic and cyclic motion. In each scenario, the vertical values of the black pixels represent 1D data of an event, while the horizontal of the scenario represents the time at which that event occurred. Looking at the scenario overall shows what action is happening. A similarity matrix has been generated for each scenario using the following:
Looking at simulated data put into a similarity matrix there are intuitive, observable differences between cyclic motion and randomly generated noise, as can be seen in Figure . Where cyclic motion is present, the pattern in the similarity matrix generated suggests something interesting is happening. Where there is no cyclic motion, there is either no obvious intuitive activity as in Figure or no evidence of repeating motion, as in Figure .
Figure 2 Examples of cyclic motion and their similarity matrices.(a) A periodic sinusoidal wave. (b) Sinusoidal wave that increases frequency with time. (c) Representation of erratic flapping motion that has similar repetitive motion. (d) Random points that reasonably fit a square wave that increases frequency with time. (e) Randomly generated points. (f) Point values that gradually increase over time. All matrices are made up of 150 points.
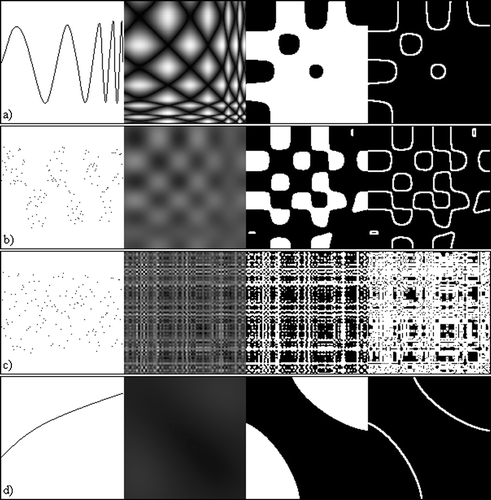
The next step forward is to interpret an observer's intuition of activity from a similarity matrix to a method for identifying it. One of the key assumptions made for identifying cyclic motion is that high amplitudes of cycles are most likely to be found at low frequencies. Noise in the similarity matrix due to frame capture, spurious pixels and poor segmentation is most likely to occur at higher frequencies. However, it is important to try and avoid dismissing cyclic motion with poor temporal matches between repeating motion, which may occur using Equation Equation(2). The following is proposed. The peak power amplitude frequency was found, and a low pass top hat filter was applied, effectively smoothing the image, removing periodicity that is likely to have been generated by noise. An Otsu threshold (Otsu Citation1979) is applied to the image. A 3×3 Sobel edge detector is then applied to distinguish between a discernable pattern in the similarity matrix from cyclic motion (mostly background pixels, few edge pixels) and noise or a nonsense similarity matrix (high proportion of edge pixels, or a very low number of edge pixels). Figure illustrates the above using a number of 1d data inputs. Figure use cyclic data, whilst figure (c) and (d) use non‐cyclic 1d data.
Figure 3 Cyclic motion detection stages. From left to right, original simulated data, similarity matrix, similarity matrix after low pass filter and Otsu threshold applied, similarity matrix after edge detection applied.(a) Sinusoidal wave that increases frequency with time. (b) Random points that fit a square wave that increases frequency with time. (c) Randomly generated points. (d) Point values that generally increase over time. All matrices are made up to 150 points.
2.1 Applying the method to real data
Clips of flying fauna were used to generate similarity matrices. Fauna included: a bat, Harris Hawk Parabuteo unicinctus, Golden Plover Pluvialis apricaria (captured using a cooled thermal imager connected to a USB digital frame grabber) and Rook Corvus frugilegus (captured using a digital video camera). All frames from all clips were converted to 256 greyscale level images prior to any processing.
There are a number of background subtraction techniques available. The method applied in this paper is based on an adaptive running average (Heikkila and Silven Citation1999). More advanced methods are available, but this approach provided good results, as rapid illumination changes, reflectance and shadow do not affect thermal imagers.
For the bat and Harris Hawk clips, the targets were segmented from the background by differencing the current frame from an updated background model. The background model was updated using
The Golden Plover and Rook were hand‐segmented frame‐by‐frame from the video clips. The Golden Plover crossed the camera viewshed in 19 frames while the Rook crossed the viewshed in 18 frames.
All of the fauna apart from the Rook flew in a linear manner across the viewshed. The Rook changed direction of flight and its body positioning during its flight across the viewshed.
Regardless of the segmentation method used, all targets were aligned by their centre of gravity. As with the simulated data, each target's similarity matrix was created using Equation Equation(3).
Once the similarity matrices had been generated, a low pass filter, as described previously was applied to the matrices, along with the Otsu threshold.
3. Results
The visual results of each stage of the cyclic motion detection method applied to the birds and bat data are shown in Figure . The similarity matrices appear to visually show flapping data. Figure shows the series of clipped frames as the bat target crosses the camera viewshed. The bat extends its wings near the start of the frame sequence and near the end. The similarity matrix for the bat (Figure ) shows this with the largest dissimilarity coinciding with the full extension of the wings.
Figure 4 Cyclic motion detection for flying fauna.(a) Bat, (b) Golden Plover, (c) Harris Hawk, (d) Rook. Left image – similarity matrix; middle image – lowpass filtered image; right image – Otsu threshold image. All images have been enlarged by 400%.

Figure 5 Segmented frames of bat target crossing the camera viewshed.
The Golden Plover similarity matrix shows a more rapid wing beat compared to the bat. This can be seen with the grid‐like structure within the matrix. The Harris Hawk similarity matrix shows a flap, followed by a soaring period, followed by a flap. The poor definition in the first flap in the matrix is due to poor segmentation in a couple of the frames. The Rook similarity matrix has some texturing, but it is difficult to ascertain cyclic or periodic motion within the matrix. However, there are still some ‘clumps’ visible in the threshold image.
4. Discussion and conclusion
Effective bird and bat surveys are essential to improving the sustainability of new developments. The use of motion detection can be used to develop and widen the application of remote surveys for monitoring bird and bat activity. Further enhancements can be made to fauna detection by capitalising on distinct flapping features, without necessarily requiring distinguishable features such as wing and body shape, hence operating on low‐resolution images.
Birds do not always fly with periodicity. Flapping variation can depend on types of flight, time length during wing oscillation and time length between wing oscillations. If target motion changes are used as a tracking method, it is important to consider cyclic oscillations in addition to periodicity.
It has been shown that cyclic motion, as well as periodic motion, can be represented in similarity matrices. A pattern can be manually observed from the matrix when an object exhibits cyclic or periodic motion. This is in contrast to when only noise is present, or the object does not oscillate while moving.
This approach has been applied to video clips of flying fauna. As with the simulated data, the similarity matrices showed distinctive patterns for each fauna example. This is useful where a limited number of frames are available. A limitation is likely to arise when there are too few frames to generate a similarity matrix. This situation is most likely to occur when a bird target is flying past the camera at a close distance. More distant targets are expected to generate a number of frames that will generate a satisfactory similarity matrix.
Bird species have varying flap patterns as well as different wing beat frequencies. It is possible to an extent to determine these flap patterns for a species from the similarity matrix. This may potentially be useful for identifying fauna species groups, further improving the efficiency of automated remote survey systems. Appropriate image processing methods such as smoothing and thresholding applied to similarity matrices may be able to provide a suitable template for target identification based on the matrix's pattern. This may displace some of the errors associated with stereo vision, and reduces the cost associated with an additional thermal, making small‐scale nocturnal fauna surveys more practical.
Automating the segmentation task would make the proposed method more practical in real‐time applications. A suggested approach could include the use of a Kalman filter to distinguish genuine moving targets from noise, applied to blobs found after the running average method as described previously has been applied. Target centre of gravity can be used as the tracking point. As targets can vary in size, all blobs may have to be treated as potential targets until the Kalman filter ‘rejects’ it by being unable to locate the target at the next ‘predicted’ location. Specific target segmentation can be achieved through background subtraction. How well this method segments a target compared to hand segmentation, and the robustness of the cyclic motion detection based on sub‐optimal segmentation would need to be explored.
5. Future work
Edge detection has yet not been applied to the real bird images, but this will be undertaken as future work.
Research will be undertaken into how processed similarity matrices can be used for identifying species groups. In addition to species group identification, determining the minimum number of frames required for generating matrices will be investigated.
Acknowledgements
This study is part of the joint EngD project with Brunel University and RPS. The project receives funding from the Engineering and Physical Sciences Research Council. The authors are also grateful for the help provided by Dr Joseph Au from the School of Engineering and Design within Brunel University.
References
- Alerstam , T. , Gudmundsson , G. A. and Larsson , B. 1993 . Flight tracks and speeds of Antarctic and Atlantic seabirds – radar and optical measurements. . Philosophical Transactions of the Royal Society of London Series B‐Biological Sciences , 340 (1) : 55 – 67 .
- Betts , A. 2007 . Are British bats at risk from windfarms? . British Wildlife , 17 (5) : 339 – 345 .
- Borowski , T. , Krol , M. , Broclawik , E. , Baranowski , T. C. , Strekowski , L. and Mokrosz , M. J. 2000 . Application of similarity matrices and genetic neural networks in quantitative structure‐activity relationships of 2‐ or 4‐(4‐methylpiperazino)pyrimidines: 5‐HT2A receptor antagonists. . Journal of Medicinal Chemistry , 43 (10) : 1901 – 1909 .
- Branzan‐Albu , A. , Yazdi , M. and Bergevin , R. 2005 . Detection of cyclic human activities based on the morphological analysis of the inter‐frame similarity matrix. . Real‐Time Imaging , 11 (3) : 219 – 232 .
- BBC. 2007. Wind ‘could power all UK homes’ [online], British Broadcasting Corporation. Available from: http://news.bbc.co.uk/1/hi/uk_politics/7135930.stm [Accessed 20 January 2008]
- BWEA. 2007. BWEA press release – Hutton to announce major offshore wind push [online], British Wind Energy Association. Available from: http://www.bwea.com/media/news/071207.html [Accessed 20 January 2008]
- Burton , N. H. K. and Armitage , M. J. S. 2005 . Differences in the diurnal and nocturnal use of intertidal feeding grounds by Redshank Tringa totanus. . Bird Study , 52 (2) : 120 – 128 .
- Cornell Lab of Ornithology. 2007. All about birds: flight pattern [online]. Cornell Laboratory of Ornithology. Available from: http://www.birds.cornell.edu/AllAboutBirds/birding123/identify/flight [Accessed 6 May 2008]
- Cutler , R. and Davis , L. S. 2000 . Robust real‐time periodic motion detection analysis and applications. . IEEE Transactions on Pattern Analysis and Machine Intelligence , 22 (8) : 781 – 796 .
- DCLG. 2000. Environmental impact assessment: guide to procedures [online], Department for Communities and Local Government. Available from: http://www.communities.gov.uk/documents/planningandbuilding/rtf/158046.rtf [Accessed 23 January 2008]
- Desholm , M. 2003 . Thermal Animal Detection System (TADS). Development of a method for estimating collision frequency of migrating birds at offshore wind turbines NERI Technical Report No. 440, National Environmental Research Institute, Denmark, 27
- Desholm , M. , Fox , A. D. and Beasley , P. 2004 . Best practice guidance for the use of remote techniques for observing bird behaviour in relation to offshore wind farms Report request commissioned by the Collaborative Offshore Wind Research into the Environment (COWRIE)
- Engel , S. , Biebach , H. and Visser , G. H. 2006 . Metabolic costs of avian flights in relation to flight velocity: a study in Rose Coloured Starlings (Sturnus rosesus, Linnaeus). . Journal of Comparative Physiology B‐Biochemical Systemic and Environmental Physiology , 176 (5) : 415 – 427 .
- Exo , K. M. , Huppop , O. and Garthe , S. 2003 . Birds and offshore wind farms: a hot topic in marine ecology. . Wader Study Group Bulletin , 100 (1) : 50 – 53 .
- Flock , W. L. 1973 . Radar observations of bird movements along the arctic coast of Alaska. . The Wilson Bulletin , 85 (3) : 259 – 275 .
- Gillings , S. , Fuller , R. J. and Sutherland , W. J. 2005 . Diurnal studies do not predict nocturnal habitat choice and site selection of European Golden‐Plovers (Pluvialis apricaria) and Northern Lapwings (Vanellus vanellus). . Auk , 122 (4) : 1249 – 1260 .
- Gudmundsson , G. A. 1994 . Spring migration of the Knot Calidris c. canutus over southern Scandinavia, as recorded by radar. . Journal of Avian Biology , 25 (1) : 15 – 26 .
- Hambly , C. , Harper , E. J. and Speakman , J. R. 2004 . The energy cost of loaded flight is substantially lower than expected due to alterations in flight kinematics. . Journal of Experimental Biology , 207 (22) : 3969 – 3976 .
- Heikkila , J. and Silven , O. 1999 . “ A real‐time system for monitoring of cyclists and pedestrians. ” . In Proceedings of the 2nd IEEE workshop on visual surveillance , 74 – 81 . Piscataway, NJ : IEEE . 26 June 1999, Fort Collins, CO, USA
- Houghton , E. W. and Blackwell , F. 1972 . Use of bird activity modulation waveforms in radar identification , Malvern, Worcester : Royal Radar Establishment .
- Kuvlesky , W. P. , Brennan , L. A. , Morrison , M. L. , Boydston , K. K. , Ballad , B. M. and Bryant , F. C. 2007 . Wind energy development and wildlife conservation: challenges and opportunities. . Journal of wildlife Management , 71 (8) : 2487 – 2498 .
- Lazarevic , L. , Southee , D. , Harrison , D. , Osmond , J. , Frost , D. and Wade , M. 2007 . “ Approaches to measure wing beat frequencies with computer vision. ” . In Proceedings of the 5th international conference on design & manufacture for sustainable development , Loughborough : Loughborough University, CD‐ROM ISBN 978‐1‐84402‐058‐4 . 10–11 July 2007, Loughborough
- McNeil , R. and Roberts , M. 1988 . Nocturnal feeding strategies of some shorebird species in a tropical environment. . Acta Congressus Internationalis Ornithologici , XIX : 2328 – 2336 .
- Mohan , R. , Medioni , G. and Nevatia , R. 1989 . Stereo error detection, correction, and evaluation. . IEEE Transactions on Pattern Analysis and Machine Intelligence , 11 (2) : 113 – 120 .
- Munoz‐Salinas , R. , Garcia‐Silvente , M. and Carnicer , R. 2008 . Adaptive multi‐modal stereo people tracking without background modelling. . Journal of Visual Communication and Image Representation , 19 (2) : 75 – 91 .
- Muller , M. and Kurth , F. 2006 . “ Enhancing similarity matrices for music audio analysis. ” . In Proceedings of the international conference on acoustic, speech and signal processing , 9 – 12 . Piscataway, NJ : IEEE . May 2006, Toulouse
- Norberg , U. M. L. 2002 . Structure, form, and function of flight in engineering and the living world. . Journal of Morphology , 252 (1) : 52 – 81 .
- Otsu , N. 1979 . A thresholding selection method from gray‐level histogram. . IEEE Transactions on Systems, Man, and Cybernetics , 9 (1) : 62 – 66 .
- Plotnik , A. M. and Rock , S. M. 2002 . Quantification of cyclic motion of marine animals from computer vision. . Oceans , 3 (1) : 29 – 31 .
- Rodriguez , J. J. and Aggarwal , J. K. 1990 . Stochastic analysis of stereo quantization error. . IEEE Transactions on Pattern Analysis and Machine Intelligence , 12 (5) : 467 – 470 .
- RSPB. 2005. Wind farms [online], Royal Society for the Protection of Birds. Available from: http://www.rspb.org.uk/ourwork/policy/windfarms/index.asp [Accessed 19 April 2007]
- Schmaljohann , H. , Liechti , F. and Bruderer , B. 2007 . Daytime passerine migrants over the Sahara – are these diurnal migrants or prolonged flights of nocturnal migrants? . Ostrich , 78 (2) : 357 – 362 .
- Schmidt‐Wellenburg , C. A. , Biebach , H. , Daan , S. and Visser , G. H. 2007 . Energy expenditure and wing beat frequency in relation to body mass in free flying Barn Swallows (Hirundo rustica). . Journal of Comparative Physiology B‐Biochemical Systemic and Environmental Physiology , 177 (3) : 327 – 337 .
- Seitz , S. M. and Dyer , C. R. 1997 . View‐invariant analysis of cyclic motion. . International Journal of Computer Vision , 25 (3) : 231 – 251 .
- Shah, M., 1997. Fundamentals of computer vision [online], Computer Science Department, University of Florida. Available from: http://www.cs.ucf.edu/courses/cap6411/book.pdf [Accessed 5 May 2008]
- Sheldon , R. , Bolton , M. , Gillings , S. and Wilson , A. M. 2004 . Conservation management of Lapwing Vanellus vanellus on lowland arable farmland in the UK. In ecology and conservation of lowland farmland birds II: the road to recovery. . Ibis , 146 (Suppl. 2) : 41 – 49 .
- Sykes Jr , P. W. , Ryman , W. E. , Kepler , C. B. and Hardy , J. W. 1995 . A 24‐hour remote surveillance system for terrestrial wildlife studies. . Journal of Field Ornithology , 66 (2) : 199 – 211 .
- Tobalske , B. W. 2007 . Biomechanics of bird flight. . Journal of Experimental Biology , 210 (18) : 3135 – 3146 .
- UNEP. 2006. Environment for development [online], United Nations Environment Programme. Available from: http://www.unep.org/PDF/ABOUT_UNEP_ENGLISH.pdf [Accessed 19 April 2007]
- Wiggelinkhuizen , E. J. , Rademakers , L. W. M. M. , Barhorst , S. A. M. and den Boon , H. J. 2006 . Bird collision monitoring system for multi‐megawatt wind turbines WT‐Bird: prototype development and testing. ECN Wind Energy ECN‐E–06‐027, Energy research Centre of the Netherlands, 57
- Williams , W. 2004 . When blade meets bat – unexpected bat kills threaten future wind farms. . Scientific American , 290 (2) : 20 – 21 .
- Zhang , P. F. , Liu , S. and Xu , Q. 2005 . Identifying Doppler velocity contamination caused by migrating birds. Part I: feature extraction and quantification. . Journal of Atmospheric and Oceanic Technology , 22 (8) : 1105 – 1113 .