
Abstract
Trypanosoma brucei, the causative agent of African trypanosomiasis, is a unicellular parasite that possesses lysosome-related organelles known as acidocalcisomes. These organelles have been found from bacteria to human cells, and are characterized by their acidic nature and high calcium and polyphosphate (polyP) content. Our proteomic analysis of acidocalcisomes of T. brucei procyclic stages, together with in situ epitope-tagging and immunofluorescence assays with specific antibodies against selected proteins, established the presence of 2 H+ pumps, a vacuolar H+-ATPase and a vacuolar H+-pyrophosphatase, that acidify the organelles as well as of a number of transporters and channels involved in phosphate metabolism, cation uptake and calcium signaling. Together with recent work in other organisms, these results provide direct evidence that acidocalcisomes are especially adapted to accumulate polyP bound to cations and for calcium signaling.
Abbreviations
| CDF | = | cation diffusion facilitator |
| Ccc1 | = | Ca2+-sensitive cross-complementer 1 |
| IP3R | = | inositol 1, 4, 5-trisphosphate receptor |
| PolyP | = | polyphosphate |
| PPX | = | exopolyphosphatase |
| VIT | = | vacuolar iron transporter |
| VTC | = | vacuolar transporter chaperone |
Acidocalcisomes were first identified in trypanosomesCitation1,2 and later found from bacteria to human cells.Citation3 Although originally described in bacteria as metachromaticCitation4 or volutinCitation5 granules they were though to lack a surrounding membrane. The discovery that in some bacteriaCitation6,7 they possess a proton pump and are able to accumulate protons (H+) and calcium ions (Ca2+) established them as real organelles and led to the recognition that, together with lipid droplets,Citation8 they are the only organelles present in both prokaryotes and eukaryotes.Citation9
Acidocalcisomes belong to the group of lysosome-related organellesCitation10 and their acidification is driven by either a vacuolar H+-ATPase or a vacuolar H+-pyrophosphatase or, in the case of trypanosomes, by both.Citation9 However, a definitive demonstration of the co-localization of these proton pumps in T. brucei was not established until our recent work.Citation11 In this regard, acidocalcisomes of T. brucei share the property of being acidified by 2 proton pumps with the plant vacuole,Citation12 the plant-like vacuole of Toxoplasma gondii,Citation13 and the digestive food vacuole of the malaria parasites.Citation14 An acidic acidocalcisome pH is required to maintain polyP in its polymeric state because their alkalinization results in polyP hydrolysis.Citation15 PolyP is a polyanion of 3 to thousands of orthophosphate monomers and binding with organic (basic amino acids, polyamines) or inorganic (calcium, magnesium, sodium, potassium, zinc, iron) cations neutralizes its negative charges.
Here, we discuss our recent proteomic analysis of acidocalcisomes of T. bruceiCitation11 to further clarify their important role in phosphate and cation homeostasis, and Ca2+ signaling. Of the 2 best-studied life stages of this organism, the procyclic stages found in the vector, and the bloodstream forms found in the mammalian host, we chose the procyclic stages, which are richer in acidocalcisomes.Citation16 To isolate acidocalcisomes we used a modification of protocols described before.Citation17,18 Cells were broken by abrasion with silicon carbide in a mortar and after subcellular fractionation that included 2 iodixanol gradient centrifugations, a fraction highly enriched in acidocalcisomes was isolated, as established by marker enzyme assays, marker antibody immunoblotting and electron microscopy analyses ( and detailed in ref. Citation11). After SDS-PAGE of the proteins, bands were cut, digested in-gel with trypsin and peptides analyzed by LC-MS/MS. Bioinformatic analysis of the resulting peptides allowed the identification of a set of candidate proteins whose acidocalcisome localization was validated by in situ epitope-tagging and western blot analyses using specific antibodies (). Knockdown by RNA interference (RNAi) of the expression of 4 of the genes encoding these proteins (vacuolar H+-ATPase subunits a and d, vacuolar iron transporter, and zinc transporter) was done to investigate their requirement for cell growth in procyclic and bloodstream forms of the parasite. Growth was greatly inhibited in both life cycle stages after knockdown of the V-H+-ATPase subunits or of the vacuolar iron transporter.
Figure 1. Flow chart shows steps for characterizing acidocalcisome proteome of Trypanosoma brucei. An integrated approach for functional analyses of the newly identified acidocalcisome proteins is proposed.
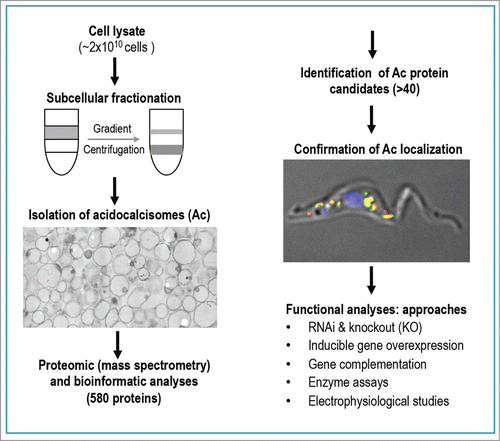
Of the 580 proteins identified with protein probability > 0.95, (listed inCitation11) over 40 were selected for further study on the basis of their presence in acidocalcisomes of other organisms, or on specific properties like presence of transmembrane domains or similarity to transporters of cations or metabolites known to be present in acidocalcisomes or lysosome-related organelles. Five proteins previously identified as localized to the acidocalcisomes by immunofluorescence assays (IFA) using epitope tagged proteins or specific antibodies, were found in the acidocalcisome proteome: vacuolar H+ pyrophosphatase (TbVP1),Citation19 plasma membrane-type vacuolar Ca2+-ATPase (PMCA, TbPMC1),Citation20 inositol 1,4,5-trisphosphate receptor (TbIP3R),Citation21 vacuolar transporter chaperone 4 (TbVtc4),Citation22 and vacuolar soluble pyrophosphatase (TbVSP1).Citation23 Four proteins were found in the acidocalcisome proteome and their localization validated by IFA using epitope-tagged proteins: vacuolar H+-ATPase subunits a and d (TbVAa and TbVAd), acid phosphatase (TbAP), and phosphate transporter (TbPho91). Four proteins that localize to acidocalcisomes or acidocalcisome-like organelles of other species, such as Zn transporter (TbZnT),Citation24 vacuolar iron transporter (TbVIT1),Citation25 polyamine transporter (TbPOT1),Citation26 and vacuolar transporter chaperone 1 (TbVtc1)Citation27,28 were not detected in the acidocalcisome proteome but were validated by IFA as localized to acidocalcisomes.Citation11 Finally, 5 proteins were detected in the acidocalcisome proteome but were localized to different organelles such as mitochondria (ABC transporter, TbABCT), nuclear membrane (hypothetical protein, TbNP), Golgi and lysosomes (Golgi /lysosome glycoprotein 1, TbGLP1), lysosomes (polyamine transporter 2, TbPOT2), and flagellar tip (cation/proton antiporter, TbFTP).Citation11
Although the acidocalcisome proteome contained 580 proteins most of them corresponded to proteins known to localize in other subcellular compartments.Citation11 This “contamination” is not unexpected because acidocalcisomes are usually observed in close contact with other organelles and intracellular structures such as the nucleus, mitochondria, subpellicular mitcrotubules, and lipid inclusionsCitation29 and parts of these structures and organelles could co-sediment with acidocalcisomes. In this regard, contact sites between organelles has been widely described.Citation30 Of the proteins validated as acidocalcisome proteins, only 2 are soluble, TbVSP1, and TbAP.
Several proteins were identified in both the T. bruceiCitation11 and the Cyanidioschyzon merolaeCitation18 acidocalcisome proteomes such as acid phosphatase, vacuolar H+-ATPase subunits a and d, vacuolar pyrophosphatase, zinc transporter, vacuolar iron transporter, and vacuolar transporter chaperone 1. There is also considerable overlap of acidocalcisome proteins with proteins localized in the yeast vacuole: V-H+-ATPase subunits, Ca2+-ATPase Pmc1p (ortholog of TbPMC1), Vtc1p and Vtc4p (orthologs of TbVtc1 and TbVtc4), Ca2+-sensitive cross-complementer 1 or Ccc1p (ortholog of TbVIT1), zinc transporter Zrc1p (ortholog of TbZnT), and phosphate transporter Pho91p (ortholog of TbPho91), and to the Arabidopsis thaliana vacuole: V-H+-ATPase subunits, vacuolar H+-pyrophosphatase, vacuolar iron transporter 1 (ortholog to TbVIT1), and metal tolerance protein 1 (ortholog to TbZnT). These results suggest a close relationship between these organelles.
The analysis of the acidocalcisome proteome reveals potential roles of the organelle in several metabolic and signaling pathwaysCitation11 (). A role in Ca2+ signaling is supported by the presence of mechanisms for Ca2+ uptake and release. Ca2+ uptake is driven by the plasma membrane-type Ca2+-ATPase (TbPMC1). This pump acts as a Ca2+/H+ countertransporting ATPase requiring an acidic intraorganellar pH for efficient Ca2+ uptake.Citation20,31 The mechanism for Ca2+ release is represented by the inositol 1,4,5-trisphosphate receptor (TbIP3R).Citation21 A previous study localized this channel to the acidocalcisomes using endogenous epitope tagging,Citation21 which was confirmed in this workCitation11 using specific antibodies. This appears to be in contrast to the apparent endoplasmic reticulum localization of the channel in T. cruzi,Citation32 although this has been disputed.Citation33
Figure 2. Schematic representation of the acidocalcisome of Trypanopsma brucei modified from ref. Citation11. The newly identified acidocalcisome proteins are marked in blue while other previously identified components are shown in black.
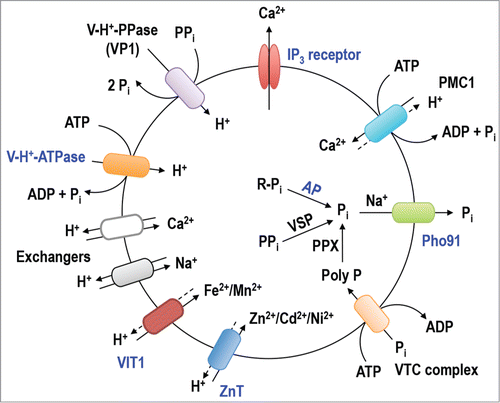
Acidocalcisomes are also important for phosphate metabolism (). In addition to enzymes involved in PPi and polyP hydrolysis, like the vacuolar soluble pyrophosphatase (TbVSP1)Citation23 and the exopolyphosphatase (PPX) activity previously reported in acidocalcisomes of T. cruzi,Citation15 our work confirmed the localization of 2 components of the vacuolar transporter chaperone (VTC) complex, TbVtc1Citation28 and TbVtc4,Citation22,34 in T. brucei acidocalcisomes.Citation11 The VTC complex, which was first described in the yeast vacuole, where it has 4 components (Vtc1–4p),Citation35 is involved in the synthesis and translocation of polyP to acidocalcisomesCitation22,34 and acidocalcisome-like vacuoles of yeastCitation35,36 and Chlamydomonas reinhardtii.Citation27 In addition to this Pi entry mechanism there is also evidence for a Pi release mechanism as suggested by the identification of a Na+/Pi symporter (TbPho91) with similarity to the Pho91p transporter localized in the yeast vacuole and responsible for Pi efflux.Citation37 Furthermore, an acidocalcisome acid phosphatase (TbAP) which could be involved in Pi release from organic molecules (R-Pi), including perhaps polyP, is also found in acidocalcisomes.Citation11
Given the polyanionic nature of polyP it is not surprising that acidocalcisomes need to accumulate cations to compensate all those negative charges (). Previous studies revealed the presence of Na+/H+ and Ca2+/H+ exchangers in acidocalcisomes of procyclic forms of T. brucei,Citation38,39 and in the new reportCitation11 we describe the localization in acidocalcisomes of a putative Zn2+ transporter (TbZnT). TbZnT is homolog to a previously described T. cruzi zinc transporterCitation24 and member of the cation diffusion facilitator (CDF) family that includes transporters involved in Zn2+, Cd2+, Co2+, and/or Ni2+ transport in exchange for protons.Citation40 Acidocalcisomes also possess a protein (TbVIT1) with similarity to the vacuolar iron transporter (Vit1p) from yeastCitation25 and plantCitation41 vacuoles involved in Fe2+ and Mn2+ sequestration. Finally a putative polyamine transporter (TbPOT1) partially localizes to the acidocalcisomes.Citation11 Polyamines, which have several positive charges, are known to be abundant in the acidocalcisome-like vacuole of yeast.Citation42 Interestingly, it was recently reported that during zinc limitation acidocalcisomes of C. reinardtiiCitation43 also accumulate copper, which was proposed as a mechanism to avoid protein mismetallation.Citation44
In conclusion, acidocalcisomes are rich in polyP and cations and in addition to this storage function, most of its functions in trypanosomes could probably be attributed to their components including their role in pHCitation19 and osmoticCitation45 regulation, response to stress,Citation15 autophagy,Citation46 growth and infectivity.Citation45,47 As several of the newly identified acidocalcisome proteins (V-H+-ATPase subunits a and d, TbVIT1) are essential for growth of procyclic and bloodstream forms of T. brucei,Citation11 and acidocalcisomes possess proteins with no similarity to mammalian proteins like the vacuolar H+-pyrophosphatase, and the VTC complex, the newly identified pathways could be appropriate targets for chemotherapy.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
This work was supported by the US. National Institutes of Health (grant AI-077538 to R.D).
References
- Vercesi AE, Moreno SN, Docampo R. Ca2+/H+ exchange in acidic vacuoles of Trypanosoma brucei. Biochem J 1994; 304:227-33; PMID:7998937
- Docampo R, Scott DA, Vercesi AE, Moreno SN. Intracellular Ca2+ storage in acidocalcisomes of Trypanosoma cruzi. Biochem J 1995; 310:1005-12; PMID:7575396
- Docampo R, de Souza W, Miranda K, Rohloff P, Moreno SN. Acidocalcisomes – conserved from bacteria to man. Nat Rev Microbiol 2005; 3:251-261; PMID:15738951; http://dx.doi.org/10.1038/nrmicro1097
- Babes V. Beobachtungen über die metachromatischen körperchen, sporenbildung, verzwiegung, kolben- und kapsel-bildung pathogener bakterien. Zentralbl Bakteriol Parasitenkd Infektionskr Hyg 1895; 20:412-20
- Meyer A. Orientierende untersuchungen über verbreitung, morphologie und chemie des volutins. Bot Zeit 1904; 62:113-52
- Seufferheld M, Lea CR, Vieira M, Oldfield E, Docampo R. The H+-pyrophosphatase of Rhodospirillum rubrum is predominantly located in polyphosphate-rich acidocalcisomes. J Biol Chem 2004; 279:51193-202; PMID:15371423; http://dx.doi.org/10.1074/jbc.M406099200
- Seufferheld M, Vieira MC, Ruiz FA, Rodrigues CO, Moreno SN, Docampo R. Identification of organelles in bacteria similar to acidocalcisomes of unicellular eukaryotes. J Biol Chem 2003; 278:29971-78; PMID:12783865; http://dx.doi.org/10.1074/jbc.M304548200
- Yang L, Ding Y, Chen Y, Zhang S, Huo C, Wang Y, Yu J, Zhang P, Na H, Zhang H, et al. The proteomics of lipid droplets: structure, dynamics, and functions of the organelle conserved from bacteria to humans. J Lipid Res 2012; 53:1245-53; PMID:22534641; http://dx.doi.org/10.1194/jlr.R024117
- Docampo R, Moreno SN. Acidocalcisomes. Cell Calcium 2011; 50:113-9; PMID:21752464; http://dx.doi.org/10.1016/j.ceca.2011.05.012
- Moreno SN, Docampo R. The role of acidocalcisomes in parasitic protists. J Eukaryot Microbiol 2009; 56:208-13; PMID:19527347; http://dx.doi.org/10.1111/j.1550-7408.2009.00404.x
- Huang G, Ulrich PN, Storey M, Johnson D, Tischer J, Tovar JA, Moreno SN, Orlando R, Docampo R. Proteomic analysis of the acidocalcisome, an organelle conserved from bacteria to human cells. PLoS Pathog 2014; 10:e1004555; PMID:25503798; http://dx.doi.org/10.1371/journal.ppat.1004555
- Maeshima M. Vacuolar H+-pyrophosphatase. Biochim Biophys Acta 2000; 1465:37-51; PMID:10748246; http://dx.doi.org/10.1016/S0005-2736(00)00130-9
- Miranda K, Pace DA, Cintron R, Rodrigues JC, Fang J, Smith A, Rohloff P, Coelho E, de Haas F, de Souza W, et al. Characterization of a novel organelle in Toxoplasma gondii with similar composition and function to the plant vacuole. Mol Microbiol 2010; 76:1358-75; PMID:20398214; http://dx.doi.org/10.1111/j.1365-2958.2010.07165.x
- Saliba KJ, Allen RJ, Zissis S, Bray PG, Ward SA, Kirk K. Acidification of the malaria parasite's digestive vacuole by a H+-ATPase and a H+-pyrophosphatase. J Biol Chem 2003; 278:5605-12; PMID:12427765; http://dx.doi.org/10.1074/jbc.M208648200
- Ruiz FA, Rodrigues CO, Docampo R. Rapid changes in polyphosphate content within acidocalcisomes in response to cell growth, differentiation, and environmental stress in Trypanosoma cruzi. J Biol Chem 2001; 276:26114-21; PMID:11371561; http://dx.doi.org/10.1074/jbc.M102402200
- Rodrigues CO, Scott DA, Docampo R. Characterization of a vacuolar pyrophosphatase in Trypanosoma brucei and its localization to acidocalcisomes. Mol Cell Biol 1999; 19:7712-23; PMID:10523660
- Salto ML, Kuhlenschmidt T, Kuhlenschmidt M, de Lederkremer RM, Docampo R. Phospholipid and glycolipid composition of acidocalcisomes of Trypanosoma cruzi. Mol Biochem Parasitol 2008; 158:120-30; PMID:18207579; http://dx.doi.org/10.1016/j.molbiopara.2007.12.001
- Yagisawa F, Nishida K, Yoshida M, Ohnuma M, Shimada T, Fujiwara T, Yoshida Y, Misumi O, Kuroiwa H, Kuroiwa T. Identification of novel proteins in isolated polyphosphate vacuoles in the primitive red alga Cyanidioschyzon merolae. Plant J 2009; 60:882-93; PMID:19709388; http://dx.doi.org/10.1111/j.1365-313X.2009.04008.x
- Lemercier G, Dutoya S, Luo S, Ruiz FA, Rodrigues CO, Baltz T, Docampo R, Bakalara N. A vacuolar-type H+-pyrophosphatase governs maintenance of functional acidocalcisomes and growth of the insect and mammalian forms of Trypanosoma brucei. J Biol Chem 2002; 277:37369-76; PMID:12121996; http://dx.doi.org/10.1074/jbc.M204744200
- Luo S, Rohloff P, Cox J, Uyemura SA, Docampo R. Trypanosoma brucei plasma membrane-type Ca2+-ATPase 1 (TbPMC1) and 2 (TbPMC2) genes encode functional Ca2+-ATPases localized to the acidocalcisomes and plasma membrane, and essential for Ca2+ homeostasis and growth. J Biol Chem 2004; 279:14427-39; PMID:14724285; http://dx.doi.org/10.1074/jbc.M309978200
- Huang G, Bartlett PJ, Thomas AP, Moreno SN, Docampo R. Acidocalcisomes of Trypanosoma brucei have an inositol 1,4,5-trisphosphate receptor that is required for growth and infectivity. Proc Natl Acad Sci U S A 2013; 110:1887-92; PMID:23319604; http://dx.doi.org/10.1073/pnas.1216955110
- Lander N, Ulrich PN, Docampo R. Trypanosoma brucei vacuolar transporter chaperone 4 (TbVtc4) Is an acidocalcisome polyphosphate kinase required for in vivo infection. J Biol Chem 2013; 288:34205-16
- Lemercier G, Espiau B, Ruiz FA, Vieira M, Luo S, Baltz T, Docampo R, Bakalara N. A pyrophosphatase regulating polyphosphate metabolism in acidocalcisomes is essential for Trypanosoma brucei virulence in mice. J Biol Chem 2004; 279:3420-5; PMID:14615483; http://dx.doi.org/10.1074/jbc.M309974200
- Ferella M, Nilsson D, Darban H, Rodrigues C, Bontempi EJ, Docampo R, Andersson B. Proteomics in Trypanosoma cruzi–localization of novel proteins to various organelles. Proteomics 2008; 8:2735-49; PMID:18546153; http://dx.doi.org/10.1002/pmic.200700940
- Li L, Chen OS, McVey Ward D, Kaplan J. CCC1 is a transporter that mediates vacuolar iron storage in yeast. J Biol Chem 2001; 276:29515-9; PMID:11390404; http://dx.doi.org/10.1074/jbc.M103944200
- Hasne MP, Coppens I, Soysa R, Ullman B. A high-affinity putrescine-cadaverine transporter from Trypanosoma cruzi. Mol Microbiol 2010; 76:78-91; PMID:20149109; http://dx.doi.org/10.1111/j.1365-2958.2010.07081.x
- Aksoy M, Pootakham W, Grossman AR. Critical function of a Chlamydomonas reinhardtii putative polyphosphate polymerase subunit during nutrient deprivation. Plant Cell 2014; 26:4214-29; PMID:25281687; http://dx.doi.org/10.1105/tpc.114.129270
- Fang J, Rohloff P, Miranda K, Docampo R. Ablation of a small transmembrane protein of Trypanosoma brucei (TbVTC1) involved in the synthesis of polyphosphate alters acidocalcisome biogenesis and function, and leads to a cytokinesis defect. Biochem J 2007; 407:161-70; PMID:17635107; http://dx.doi.org/10.1042/BJ20070612
- Miranda K, Benchimol M, Docampo R, de Souza W. The fine structure of acidocalcisomes in Trypanosoma cruzi. Parasitol Res 2000; 86:373-384; PMID:10836511; http://dx.doi.org/10.1007/s004360050682
- Helle SC, Kanfer G, Kolar K, Lang A, Michel AH, Kornmann B. Organization and function of membrane contact sites. Biochim Biophys Acta 2013; 1833:2526-41; PMID:23380708; http://dx.doi.org/10.1016/j.bbamcr.2013.01.028
- Scott DA, Docampo R. Characterization of isolated acidocalcisomes of Trypanosoma cruzi. J Biol Chem 2000; 275:24215-21; PMID:10816577; http://dx.doi.org/10.1074/jbc.M002454200
- Hashimoto M, Enomoto M, Morales J, Kurebayashi N, Sakurai T, Hashimoto T, Nara T, Mikoshiba K. Inositol 1,4,5-trisphosphate receptor regulates replication, differentiation, infectivity and virulence of the parasitic protist Trypanosoma cruzi. Mol Microbiol 2013; 87:1133-1150; PMID:23320762; http://dx.doi.org/10.1111/mmi.12155
- Docampo R, Moreno SN, Plattner H. Intracellular calcium channels in protozoa. Eur J Pharmacol 2014; 739:4-18; PMID:24291099; http://dx.doi.org/10.1016/j.ejphar.2013.11.015
- Ulrich PN, Lander N, Kurup S, Reiss L, Brewer J, Soares Medeiros LC, Miranda K, Docampo R. The aciodcalcisome vacuolar transporter chaperone 4 catalyzes the synthesis of polyphosphate in insect-stages of Trypanosoma brucei and T. cruzi. J Eukaryot Microbiol 2014; 61:155-65; PMID:24386955; http://dx.doi.org/10.1111/jeu.12093
- Hothorn M, Neumann H, Lenherr ED, Wehner M, Rybin V, Hassa PO, Uttenweiler A, Reinhardt M, Schmidt A, Seiler J, et al. Catalytic core of a membrane-associated eukaryotic polyphosphate polymerase. Science 2009 324:513-6; PMID:19390046; http://dx.doi.org/10.1126/science.1168120
- Gerasimaite R, Sharma S, Desfougeres Y, Schmidt A, Mayer A. Coupled synthesis and translocation restrains polyphosphate to acidocalcisome-like vacuoles and prevents its toxicity. J Cell Sci 2014; 127:5093-104; PMID:25315834; http://dx.doi.org/10.1242/jcs.159772
- Hurlimann HC, Stadler-Waibel M, Werner TP, Freimoser FM. Pho91 Is a vacuolar phosphate transporter that regulates phosphate and polyphosphate metabolism in Saccharomyces cerevisiae. Mol Biol Cell 2007; 18:4438-45; PMID:17804816; http://dx.doi.org/10.1091/mbc.E07-05-0457
- Vercesi AE, Docampo R. Sodium-proton exchange stimulates Ca2+ release from acidocalcisomes of Trypanosoma brucei. Biochem J 1996; 315:265-70; PMID:8670117
- Vercesi AE, Grijalba MT, Docampo R. Inhibition of Ca2+ release from Trypanosoma brucei acidocalcisomes by 3,5-dibutyl-4-hydroxytoluene: role of the Na+/H+ exchanger. Biochem J 1997; 328:479-82; PMID:9371704
- MacDiarmid CW, Gaither LA, Eide D. Zinc transporters that regulate vacuolar zinc storage in Saccharomyces cerevisiae. EMBO J 2000; 19:2845-55; PMID:10856230; http://dx.doi.org/10.1093/emboj/19.12.2845
- Kim SA, Punshon T, Lanzirotti A, Li L, Alonso JM, Ecker JR, Kaplan J, Guerinot ML. Localization of iron in Arabidopsis seed requires the vacuolar membrane transporter VIT1. Science 2006; 314:1295-8; PMID:17082420; http://dx.doi.org/10.1126/science.1132563
- Tomitori H, Kashiwagi K, Asakawa T, Kakinuma Y, Michael AJ, Igarashi K. Multiple polyamine transport systems on the vacuolar membrane in yeast. Biochem J 2001; 353:681-8; PMID:11171066; http://dx.doi.org/10.1042/0264-6021:3530681
- Ruiz FA, Marchesini N, Seufferheld M, Govindjee, Docampo R. The polyphosphate bodies of Chlamydomonas reinhardtii possess a proton-pumping pyrophosphatase and are similar to acidocalcisomes. J Biol Chem 2001; 276:46196-203; PMID:11579086; http://dx.doi.org/10.1074/jbc.M105268200
- Hong-Hermesdorf A, Miethke M, Gallaher SD, Kropat J, Dodani SC, Chan J, Barupala D, Domaille DW, Shirasaki DI, Loo JA, et al. Subcellular metal imaging identifies dynamic sites of Cu accumulation in Chlamydomonas. Nat Chem Biol 2014; 10:1034-42; PMID:25344811; http://dx.doi.org/10.1038/nchembio.1662
- Huang G, Fang J, Sant'Anna C, Li ZH, Wellems DL, Rohloff P, Docampo R. Adaptor protein-3 (AP-3) complex mediates the biogenesis of acidocalcisomes and is essential for growth and virulence of Trypanosoma brucei. J Biol Chem 2011; 286:36619-30; PMID:21880705; http://dx.doi.org/10.1074/jbc.M111.284661
- Li FJ, He CY. Acidocalcisome is required for autophagy in Trypanosoma brucei. Autophagy 2014; 10:1978-88; PMID:25484093; http://dx.doi.org/10.4161/auto.36183
- Galizzi M, Bustamante JM, Fang J, Miranda K, Soares Medeiros LC, Tarleton RL, Docampo R. Evidence for the role of vacuolar soluble pyrophosphatase and inorganic polyphosphate in Trypanosoma cruzi persistence. Mol Microbiol 2013; 90:699-715; PMID:24033456; http://dx.doi.org/10.1111/mmi.12392