
Abstract
Fungal plant pathogenicity is facilitated by effector proteins that are specifically expressed during infection and are responsible for suppressing plant defense mechanisms. Recent studies have elucidated the detailed molecular mechanisms of effector action throughout fungal infection. However, little is known about the trafficking and secretion of effectors in fungal hyphae during the initial stage of infection. Using state-of-the-art microscopy we have demonstrated that early endosome (EE) motility is required for effector production during fungal infection. Moreover, the MAPK Crk1 has been shown to travel on EEs and to function as a negative regulator of effector expression, suggesting that motile EEs are involved in signal transduction. Here I further discuss possible mechanisms whereby EE motility regulates effector expression in the initial stages of infection.
The corn smut fungus Ustilago maydis is a very well-studied plant pathogen.Citation1–3 Recent work in this fungus has revealed that there are several effectors essential for pathogenicity; for example, Cmu1 is a chorismate mutase that decreases the production of salicylic acid – a key defense signaling molecule,Citation4 and Pep1 and Pit2 are inhibitors of apoplastic plant peroxidases and proteases, respectively.Citation5,6 All such effectors are specifically expressed during the infection stage to reduce plant defense responses. In addition to investigation of microbe-plant infection mechanisms, cell biological analyses have also been carried out in U. maydis.Citation7 In fact, the fungus is nowadays considered as a model organism for understanding the molecular mechanisms of intracellular membrane trafficking, especially regarding early endosomes (EEs), which are unique organelles showing constant rapid and long-range motility that is regulated by bi-directional motor activity, kinesin-3 and dynein.Citation8,9 Despite the elucidation of the molecular mechanism of EE motility, there was little known about the physiological importance of this transport process. Indeed, it was a mystery as to why EEs are constantly in motion throughout hyphal cells given the metabolic burden inherent in this manner of active transport. Recent publications have shown that the translation machinery is delivered on motile EEs, which supports the even distribution of ribosomes inside hyphae.Citation10,11 However, there was still possibility that EE motility has more physiological roles, especially in the infection phase, where the complex molecular interactions between plant and fungus remain to be fully understood. Since elongated fungal hypha most likely needs long-range communication between the tip and the nucleus,Citation12 EEs might be responsible for transducing signals in a similar manner to mammalian cells, where such retrograde signaling is mediated by EE motility.Citation13-16 However, there were no reports of the involvement of fungal EE motility in signal transduction during plant infection.
First of all, to ascertain whether EEs are able to travel from the penetrating tip to the nucleus, we performed in planta microscopy of U. maydis expressing the EE marker Rab5a fused to photo-activatable GFP (paGFPCitation17). Using state-of-the-art microscopy techniques, we demonstrated that approximately one fourth of EEs display a retrograde run-length that is greater than the distance between the hyphal tip and the nucleus. Then, to dissect whether EE motility plays an important role in early infection stage, we developed a synthetic EE anchoring protein (EAP) that consists of rigor mutant of kinesin-1 motor domain, which is unable to move along the microtubule cytoskeleton, and the PX domain from the t-SNARE Yup1, which captures EEs.Citation18 EAP expression successfully stops EEs motility, but importantly does not affect motor-driven intercellular trafficking, such as the motility of kinesin-3 and dynein and the secretion of Mcs1, a cell-wall building enzyme localized to the plasma membrane. Using two promoters with different expression profiles in infected plant cells to express EAP, we demonstrated that EE motility in initial infection step is crucial for full virulence. In agreement with this, secretion of 3 already well-characterized effector proteins Cmu1, Pep1 and Pit2 was reduced. We further demonstrated that the transcription level of these effector genes was reduced, suggesting that EE motility is involved in the level of transcription of effector genes, which is vital for pathogenicity. To further support the obtained data, we employed 2 mutants: Δhok1 and Δrab5a. Hok1 is an adaptor protein essential for the attachment between motor proteins and EEs.Citation19,20 Rab5a is the small GTPase specifically localized to EEs and has an important role in their function.Citation21 Both Δhok1 and Δrab5a cells, compared with EAP expressing cells, displayed almost identical results in effector transcription and secretion and pathogenicity. Finally, to examine whether certain signaling molecules co-migrate on EEs, we focused on mitogen-activated protein kinase (MAPK) modules because they are widely conserved signaling components and are found on mammalian Rab5-positive EEs.Citation15 Among 6 candidate MAPK modules in U. maydis, we found that Crk1 shows co-motility with EEs and its null mutant displays enhanced transcription and secretion of effectors, suggesting that this MAPK is involved in negative regulation of effector production. Apart from Crk1, in order to tune the expression level of effector genes, there probably exists other molecules to upregulate effector production, although we could not find out. Since effector transcription is repressed in the absence of EE motility, it is reasonable to expect a reduction of effector secretion. In this respect, it is possible that effector secretion directly depends on EE-mediated trafficking - a scenario which we could not distinguish in the article. Now I propose one possible experiment to evaluate whether EE motility also contributes to effector secretion. If overexpression of an effector gene could induce effector secretion even in both the presence and absence of EAP expression, it is likely that EE motility is not responsible for effector secretion to the penetrating tip and there is another mechanism for effector secretion. Further research into whether EE motility is involved in effector secretion is needed.
Investigations of effectors expression and production by quantitative analyses and state-of-the-art microscopy have elucidated that EE motility is essential for the initial infection (). However, this finding has raised several new questions, primarily: what the initial signaling molecule(s) sensed at the plasma membrane to trigger effector expression and are there more signal transducing factors on motile EEs, apart from Crk1? Recent work demonstrated that Sho1 and Msb2 are sensor proteins to induce the formation of penetrating structure appressoria, which is mediated by plant hydrophobic surface.Citation22 For appressoria formation, hydroxyl fatty acid is also an important factor,Citation23 but how it is sensed by the fungus is not known. Importantly, Sho1 interacts with a MAPK Kpp6, which affects virulence.Citation24 We could not observe any motility of Kpp6 in our study, thus this MAPK probably does not depend on EE motility to regulate effector expression. Also, we do not know how signaling elements can be moved from the moving organelles into the nucleus. Further dissection of the complex interplay of signal transduction between the plasma membrane through EEs to the nucleus will provide crucial information about how fungi successfully infect plants, which will also be beneficial for understanding the detailed molecular mechanisms of other microbe-plant interactions.
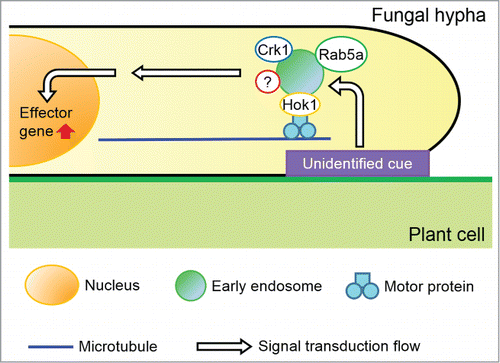
Figure 1. A model depicting effector production via early endosome motility. Initial step is signaling perception from the plant surface by unidentified mechanism, which is transduced to early endosomes (EEs). Motile EEs, harboring Rab5a, Hok1, the MAPK Crk1 and likely unknown protein as signal transducing molecules, travel from the hyphal tip to the nucleus, where effector genes are expressed. Thereafter, effector proteins are delivered to the penetrating hyphal tip to be secreted into the apoplastic space between fungal and plant cell walls or even into plant cells to suppress plant defense responses. If EE motility is impaired, signaling that is crucial for effector expression cannot be transduced to the nucleus, likely even with perception of initial cue from the plasma membrane to EEs, resulting in no effector production and less virulence.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
I would like to thank Professor Gero Steinberg for his supervision on this research project held in Biosciences, University of Exeter, UK. I am grateful to Dr. Adokiye Berepiki for his critical and constructive comments on the manuscript.
References
- Kämper J, Kahmann R, Bölker M, Ma LJ, Brefort T, Saville BJ, Banuett F, Kronstad JW, Gold SE, Müller O, et al. Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 2006; 444:97-101; PMID:17080091; http://dx.doi.org/10.1038/nature05248
- Brefort T, Doehlemann G, Mendoza-Mendoza A, Reissmann S, Djamei A, Kahmann R. Ustilago maydis as a pathogen. Annu Rev Phytopathol 2009; 47:423-45; PMID:19400641; http://dx.doi.org/10.1146/annurev-phyto-080508-081923
- Djamei A, Kahmann R. Ustilago maydis: dissecting the molecular interface between pathogen and plant. PLoS Pathog 2012; 8:e1002955; PMID:23133380; http://dx.doi.org/10.1371/journal.ppat.1002955
- Djamei A, Schipper K, Rabe F, Ghosh A, Vincon V, Kahnt J, Osorio S, Tohge T, Fernie AR, Feussner I, et al. Metabolic priming by a secreted fungal effector. Nature 2011; 478:395-8; PMID:21976020; http://dx.doi.org/10.1038/nature10454
- Doehlemann G, van der Linde K, Assmann D, Schwammbach D, Hof A, Mohanty A, Jackson D, Kahmann R. Pep1, a secreted effector protein of Ustilago maydis, is required for successful invasion of plant cells. PLoS Pathog 2009; 5:e1000290; PMID:19197359; http://dx.doi.org/10.1371/journal.ppat.1000290
- Doehlemann G, Reissmann S, Assmann D, Fleckenstein M, Kahmann R. Two linked genes encoding a secreted effector and a membrane protein are essential for Ustilago maydis -induced tumor formation. Mol Microbiol 2011; 81:751-66; PMID:21692877; http://dx.doi.org/10.1111/j.1365-2958.2011.07728.x
- Steinberg G, Perez-Martin J. Ustilago maydis, a new fungal model system for cell biology. Trends Cell Biol 2008; 18:61-7; PMID:18243705; http://dx.doi.org/10.1016/j.tcb.2007.11.008
- Steinberg G. Motors in fungal morphogenesis: cooperation versus competition. Curr Opin Microbiol 2011; 14:660-7; PMID:22030446; http://dx.doi.org/10.1016/j.mib.2011.09.013
- Steinberg G. Endocytosis and early endosome motility in filamentous fungi. Curr Opin Microbiol 2014; 20C:10-18; PMID:24835422; http://dx.doi.org/10.1016/j.mib.2014.04.001
- Baumann S, König J, Koepke J, Feldbrügge M. Endosomal transport of septin mRNA and protein indicates local translation on endosomes and is required for correct septin filamentation. EMBO Rep 2014; 15:94-102; PMID:24355572; http://dx.doi.org/10.1002/embr.201338037
- Higuchi Y, Ashwin P, Roger Y, Steinberg G. Early endosome motility spatially organizes polysome distribution. J Cell Biol 2014; 204:343-57; PMID:24493587; http://dx.doi.org/10.1083/jcb.201307164
- Steinberg G. On the move: endosomes in fungal growth and pathogenicity. Nat Rev Microbiol 2007; 5:309-16; PMID:17325725; http://dx.doi.org/10.1038/nrmicro1618
- Sorkin A, Von Zastrow M. Signal transduction and endocytosis: close encounters of many kinds. Nat Rev Mol Cell Biol 2002; 3:600-14; PMID:12154371; http://dx.doi.org/10.1038/nrm883
- Howe CL, Mobley WC. Signaling endosome hypothesis: A cellular mechanism for long distance communication. J Neurobiol 2004; 58:207-16; PMID:14704953; http://dx.doi.org/10.1002/neu.10323
- Miaczynska M, Pelkmans L, Zerial M. Not just a sink: endosomes in control of signal transduction. Curr Opin Cell Biol 2004; 16:400-6; PMID:15261672; http://dx.doi.org/10.1016/j.ceb.2004.06.005
- Miaczynska M, Bar-Sagi D. Signaling endosomes: seeing is believing. Curr Opin Cell Biol 2010; 22:535-40; PMID:20538448; http://dx.doi.org/10.1016/j.ceb.2010.05.007
- Schuster M, Kilaru S, Fink G, Collemare J, Roger Y, Steinberg G. Kinesin-3 and dynein cooperate in long-range retrograde endosome motility along a nonuniform microtubule array. Mol Biol Cell 2011; 22:3645-57; PMID:21832152; http://dx.doi.org/10.1091/mbc.E11-03-0217
- Wedlich-Söldner R, Bölker M, Kahmann R, Steinberg G. A putative endosomal t-SNARE links exo- and endocytosis in the phytopathogenic fungus Ustilago maydis. EMBO J 2000; 19:1974-86; PMID:10790364
- Bielska E, Schuster M, Roger Y, Berepiki A, Soanes DM, Talbot NJ, Steinberg G. Hook is an adapter that coordinates kinesin-3 and dynein cargo attachment on early endosomes. J Cell Biol 2014; 204:989-1007; PMID:24637326; http://dx.doi.org/10.1083jcb.201309022
- Zhang J, Qiu R, Arst HN Jr, Peñalva MA, Xiang X. HookA is a novel dynein-early endosome linker critical for cargo movement in vivo. J Cell Biol 2014; 204:1009-26; PMID:24637327; http://dx.doi.org/10.1083/jcb.201308009
- Fuchs U, Hause G, Schuchardt I, Steinberg G. Endocytosis is essential for pathogenic development in the corn smut fungus Ustilago maydis. Plant Cell 2006; 18:2066-81; PMID:16798890; http://dx.doi.org/10.1105/tpc.105.039388
- Lanver D, Mendoza-Mendoza A, Brachmann A, Kahmann R. Sho1 and Msb2-related proteins regulate appressorium development in the smut fungus Ustilago maydis. Plant Cell 2010; 22:2085-101; PMID:20587773; http://dx.doi.org/10.1105/tpc.109.073734
- Mendoza-Mendoza A, Berndt P, Djamei A, Weise C, Linne U, Marahiel M, Vranes M, Kämper J, Kahmann R. Physical-chemical plant-derived signals induce differentiation in Ustilago maydis. Mol Microbiol 2009; 71:895-911; PMID:19170880; http://dx.doi.org/10.1111/j.1365-2958.2008.06567.x
- Lanver D, Berndt P, Tollot M, Naik V, Vranes M, Warmann T, Münch K, Rössel N, Kahmann R. Plant surface cues prime Ustilago maydis for biotrophic development. PLoS Pathog 2014; 10:e1004272; PMID:25033195; http://dx.doi.org/10.1371/journal.ppat.1004272