
ABSTRACT
Much debate concerns whether any nonhuman animals share with humans the ability to infer others' mental states, such as desires and beliefs. In a recent eye-tracking false-belief task, we showed that great apes correctly anticipated that a human actor would search for a goal object where he had last seen it, even though the apes themselves knew that it was no longer there. In response, Heyes proposed that apes' looking behavior was guided not by social cognitive mechanisms but rather domain-general cueing effects, and suggested the use of inanimate controls to test this alternative submentalizing hypothesis. In the present study, we implemented the suggested inanimate control of our previous false-belief task. Apes attended well to key events but showed markedly fewer anticipatory looks and no significant tendency to look to the correct location. We thus found no evidence that submentalizing was responsible for apes' anticipatory looks in our false-belief task.
The ability to infer the mental states of others, such as their desires and beliefs, is known as theory of mind (ToM).Citation1 ToM is central to many key facets of human social life—including our unique forms of communication, cooperation, and culture—and consequently, researchers have long sought to determine whether ToM is also unique to humans.Citation2 Several decades of research have suggested that great apes, humans' closest living relatives, as well as some distantly related species (e.g., corvids) are able to infer and track the goals, perception, and knowledge that motivates others' actions.Citation3-5 However, until recently, despite a variety of investigations, there was no evidence that any nonhuman animals could understand others' false beliefs.Citation6-9 Attributing false beliefs requires recognizing that others' actions are guided not by reality but by their beliefs about reality, even when those beliefs are false. In humans, this ability has long been seen as the signature of a developed ToM.Citation10
Adapting a seminal eye-tracking paradigm from developmental psychology,Citation11 Krupenye, Kano, et al.Citation12 recently showed that great apes could pass a modified false belief test. Apes watched videos in which an actor searched for a goal object. In key events, the actor witnessed the object's hiding in one of 2 locations, briefly left the scene, and the object was moved and/or removed while the actor was gone. Thus, when the actor returned to search for the object, he now falsely believed that it was in its original location. Krupenye, Kano, et al.Citation12 used an infrared eye-tracker to noninvasively monitor apes' eye movements and map their gaze onto the films. Across 2 experiments, as the actor moved centrally toward the 2 potential hiding places, apes looked to the location where the actor had last seen the goal object, correctly anticipating that the actor would search for the object where he falsely believed it to be. These findings provided the first evidence that great apes may be able to predict others' actions by inferring their false beliefs.
These results remain open, however, to some alternative explanations (e.g., refs. Citation13, 14). One prominent account, offered by Heyes,Citation14-16 is that apes—as well as human infants and adults—solve this and other related tasks not by attributing mental states to the actor but by submentalizing. Submentalizing is “prediction of behavior by low-level, domain-general psychological processes.”Citation14 Commenting specifically on Krupenye, Kano, et al.Citation12 experiment 2, HeyesCitation14 suggested that rather than tracking where the actor last saw the object, and where he believed it to be, apes may have simply encoded low-level properties of the stimuli like “the appearance and disappearance of the [actor's] striking green shirt.” When the actor returned to the scene to search for his goal object, the green shirt could have served as a retrieval cue, eliciting a memory of the box that contained the object when the green shirt was last present. Then, “the orientation of the green object relative to the boxes and the brick [the goal object] before the green object's disappearance could have acted as a contextual cue priming the apes' visual search when the green object reappeared.”
HeyesCitation14 encouraged researchers to control for domain-general processes in ToM tests by implementing inanimate controls that mirror the perceptual features of test stimuli but reduce their agentic qualities. Some evidence from a previous anticipatory looking test involving an inanimate control already suggests that submentalizing is not responsible for apes' performance in ToM tasks.Citation17 However, to test Heyes'Citation14 proposal that retrieval and contextual cueing may explain apes' success in our false belief task, in the present study we implemented an inanimate control. The submentalizing hypothesis predicts that, given similar attention, retrieval and contextual cueing will elicit similar anticipatory looking patterns in the control as in the original study.
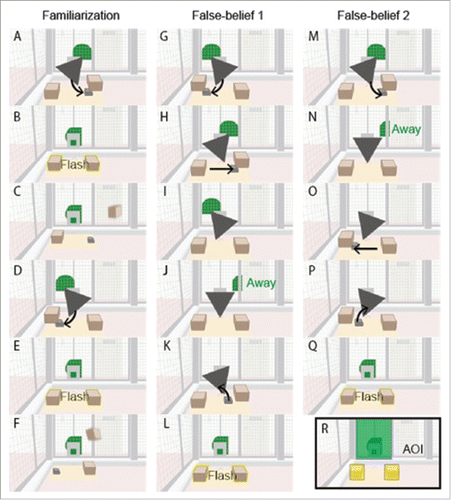
Apes (N = 43) were situated in a testing room and watched video stimuli on a monitor while an infrared eye-tracker noninvasively recorded their eye movements [60 Hz; X120 in WKPRC and X300 in KS; Tobii Technology AB, Stockholm, Sweden]. In the control test, we used the same methods as the original study, except that apes watched different stimuli (). In the original stimuli, a human actor sought a stone that a costumed gorilla-like character, King-Kong, had hidden in one of 2 boxes. Created based on of Heyes,Citation14 control stimuli were inanimate versions of the original in which the human actor was swapped out for a green semi-circle and the King-Kong antagonist for a gray triangle. The fore- and background features of the control stimuli were drawn as similarly as possible to the original stimuli. In the critical test trials, the green semi-circle was present while the goal object, a stone, hid under one of the 2 boxes. The green shape then left the scene and, while away, the stone was moved and/or removed. The key question was whether, upon the green shape's return, apes would spontaneously look to the Target box that contained the stone when the green shape was last present, as opposed to the other Distractor box.
Figure 1. Frames from the inanimate control stimuli. See the video online (https://youtu.be/J9hJBLcHc2A).
Apes closely tracked all key events (). However, despite a much larger sample, just 22 of 43 apes (51.16%) looked to one of the boxes during the anticipatory phase, as compared with 22 of 30 apes (73.33%) in the original study (). Moreover, unlike in the original study, among those that did make looks, there was no significant tendency to look first to the Target over the Distractor, overall (14 of 22 first looks to the target, p = 0.28, 2-tailed binomial test; ) or in each condition (FB1: 6 of 11 first looks to the target, p = 1.0; FB2: 8 of 11 first looks to the target, p = 0.23, 2-tailed binomial test). Additionally, an ANOVA investigating viewing times revealed no significant looking bias toward either the Target or Distractor, and a notably smaller effect size (F(1,37) = 0.31, p = 0.57, η2 = 0.008; ). The other main and interaction effects were also not significant (ps>0.05). Finally, analyzing only apes who participated in both the control and original studies (N = 26) revealed the same pattern of results; i.e. no significant bias in first looks (p = 0.33 in a binomial test) or viewing times (F(1,20) = 1.03, p = 0.32, η2 = 0.05 in ANOVA) in the control test.
Figure 2. Viewing times (msec ± SE) for key events during the belief-induction and the anticipatory-looking phases of the false-belief 1 (FB1) and false-belief 2 (FB2) conditions.
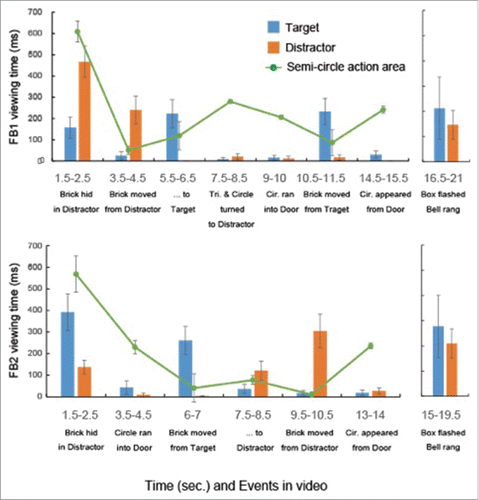
Table 1. Number of first looks in the original and control studies.
The submentalizing hypothesis predicts that, given similar attention, retrieval and contextual cueing will elicit similar anticipatory looking patterns in inanimate controls as in false belief test stimuli. In contrast to this prediction, we found that, although apes closely tracked key events as they did in the original study, they made markedly fewer looks to the Target or Distractor locations. Additionally, those that did make looks showed no significant tendency to look longer at the Target or the Distractor. Although somewhat more apes looked first to the Target than the Distractor (particularly in the FB2 condition), this tendency was not significant overall or in either condition. Moreover, and critically, the submentalizing hypothesis is only viable if it can account for successful performance across both conditions. Taken together, in the absence of the most prominent agentic features, apes did not automatically exhibit comparable anticipatory looking patterns to those exhibited in our false belief test. These results suggest that the domain-general processes implicated by the submentalizing hypothesis, namely retrieval and contextual cueing, are insufficient to explain apes' ability to correctly anticipate the actions of a mistaken agent in our task.Citation12 Determining the exact cognitive mechanism that facilitated their successful action prediction (e.g., perspective-taking,Citation7 rule-based behavior prediction,Citation13 etc.) and the nature of the representations involved in this mechanism,Citation18-21 however, remain key areas for future research.
Participants and methods
A total of 43 great apes (14 bonobos, Pan paniscus, 22 chimpanzees, Pan troglodytes, 7 orangutans, Pongo abelii) participated in this study, 26 of which had participated in the original study. Apes were born in captivity and lived with conspecifics in enriched naturalistic environments at the Wolfgang Kohler Primate Research Center (WKPRC) in Leipzig, Germany, and at Kumamoto Sanctuary (KS) in Kumamoto, Japan. Animal husbandry and research protocol complied with local guidelines, which strictly adhere to international standards and the national laws of Japan and Germany.
See referenceCitation12 for full details of the procedure of the original study, which the present study followed. Participants experienced 2 familiarization trials on one day and a single test trial on the next. In familiarization trials, while the green shape was present, the stone moved either under the left or right box (one familiarization of each type per subject), the green shape's orientation changed, and then the boxes flashed and slowly moved toward the green shape (4.5 seconds of boxes flashing and bell ringing; reflecting the anticipatory phase in which, in the original stimuli, the human actor reached toward the flashing boxes and then pulled in the tray that they sat on).
Test trials were one of 2 conditions: false-belief 1 (FB1) or false-belief 2 (FB2), with half of participants assigned to each condition (between-subjects design). In FB1, while the green shape was present, the stone moved first under one box and then under the other (and then the gray triangle turned to the first box). The green shape then left the scene and, while away, the stone was removed. In FB2, while the green shape was present, the stone moved under the first box. The green shape then left the scene and, while away, the stone moved to the second box and was then removed. In both conditions, the green shape then returned, changed its orientation, and the boxes began flashing and moving toward the green shape (again 4.5 seconds of boxes flashing and bell ringing). We counterbalanced across subjects the order of familiarization trials (stone on Left then Right or Right then Left) and whether, in the test trials, the Target was the left or right box.
As in the original study, the Target box was defined as the box where the green shape would falsely believe the stone to be located if it were an agent (i.e., the box where the stone was located before the green shape left the scene). The other box was the Distractor. Apes' eye-movement responses (first looks and viewing times) to each scene feature (e.g., Target and Distractor boxes) were coded automatically in the Tobii Studio software (ver. 3.2.1) based on Areas-Of-Interest (AOIs; see ). As in the original study, we used a binomial test to determine whether apes looked first to the Target box significantly above chance, and an ANOVA to determine whether viewing times differed significantly between Target/Distractor locations, conditions (FB1 vs FB2), or species.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Dr. Cecilia Heyes for discussing the details of our inanimate submentalizing control experiment reported here, and the staff at WKPRC and Kumamoto Sanctuary for their assistance performing the study.
Funding
Financial support came from Ministry of Education, Culture, Sports, Science and Technology (K-CONNEX to FK), Japan Society for Promotion of Science (KAKENHI 26885040, 16K21108 to FK), JSPS (KAKENHI 26245069, 24000001 to SH), and European Research Council (Synergy grant 609819 SOMICS to JC).
References
- Premack D, Woodruff G. Does the chimpanzee have a theory of mind. Behavioral Brain Sci 1978; 1(4):515-26; https://doi.org/10.1017/S0140525X00076512
- Tomasello M, Carpenter M, Call J, Behne T, Moll H. Understanding and sharing intentions: The origins of cultural cognition. Behavioral Brain Sci 2005; 28(05):675-91; PMID:16262930; https://doi.org/10.1017/S0140525X05000129
- Call J, Tomasello M. Does the chimpanzee have a theory of mind? 30 years later. Trends Cognitive Sci 2008; 12(5):187-92; PMID:18424224; https://doi.org/10.1016/j.tics.2008.02.010
- Clayton NS, Dally JM, Emery NJ. Social cognition by food-caching corvids. The western scrub-jay as a natural psychologist. Philos Trans R Soc Lond B Biol Sci 2007; 362(1480):507-22; https://doi.org/10.1098/rstb.2006.1992
- Bugnyar T, Reber SA, Buckner C. Ravens attribute visual access to unseen competitors. Nat Commun 2016; 7:10506; PMID:26835849; https://doi.org/10.1038/ncomms10506
- Call J, Tomasello M. A nonverbal false belief task: The performance of children and great apes. Child Dev 1999; 70(2):381-95; PMID:10218261; https://doi.org/10.1111/1467-8624.00028
- Hare B, Call J, Tomasello M. Do chimpanzees know what conspecifics know? Animal Behaviour 2001; 61(1):139-51; PMID:11170704; https://doi.org/10.1006/anbe.2000.1518
- Kaminski J, Call J, Tomasello M. Chimpanzees know what others know, but not what they believe. Cognition 2008; 109(2):224-34; PMID:18849023; https://doi.org/10.1016/j.cognition.2008.08.010
- Krachun C, Carpenter M, Call J, Tomasello M. A competitive nonverbal false belief task for children and apes. Dev Sci 2009; 12(4):521-35; PMID:19635080; https://doi.org/10.1111/j.1467-7687.2008.00793.x
- Wellman HM, Cross D, Watson J. Meta-analysis of theory-of-mind development: The truth about false belief. Child Dev 2001; 72:655-84; PMID:11405571; https://doi.org/10.1111/1467-8624.00304
- Southgate V, Senju A, Csibra G. Action anticipation through attribution of false belief by 2-year-olds. Psychol Sci 2007; 18(7):587-92; PMID:17614866; https://doi.org/10.1111/j.1467-9280.2007.01944.x
- Krupenye C, Kano F, Hirata S, Call J, Tomasello M. Great apes anticipate that other individuals will act according to false beliefs. Science 2016; 354(6308):110-4; PMID:27846501; https://doi.org/10.1126/science.aaf8110
- Perner J, Ruffman T. Infants' insight into the mind: How deep? Science 2005; 308:214-6; PMID:15821079; https://doi.org/10.1126/science.1111656
- Heyes C. Apes Submentalise. Trends Cognitive Sci 2017; 21(1):1-2.
- Heyes C. False belief in infancy: a fresh look. Dev Sci 2014; 17(5):647-59; PMID:24666559; https://doi.org/10.1111/desc.12148
- Heyes C. Submentalizing: I am not really reading your mind. Perspect Psychol Sci 2014; 9(2):131-43; PMID:26173251; https://doi.org/10.1177/1745691613518076
- Kano F, Call J. Great apes generate goal-based action predictions: an eye-tracking study. Psychol Sci 2014; 25(9):1691-8; PMID:25022278; https://doi.org/10.1177/0956797614536402
- Gruber T, Zuberbühler K, Clément F, van Schaik C. Apes have culture but may not know that they do. Front Psychol 2015; 6:91.
- Baillargeon R, Scott RM, He Z. False-belief understanding in infants. Trends Cognit Sci 2010; 14(3):110-8; PMID:20106714; https://doi.org/10.1016/j.tics.2009.12.006
- Apperly IA, Butterfill SA. Do humans have two systems to track beliefs and belief-like states? Psychol Rev 2009; 116(4):953; PMID:19839692; https://doi.org/10.1037/a0016923
- Martin A, Santos LR. What Cognitive Representations Support Primate Theory of Mind? Trends Cogn Sci 2016; 20(5):375-82; PMID:27052723; https://doi.org/10.1016/j.tics.2016.03.005