
ABSTRACT
The soils beneath and surrounding mature Artocarpus mariannensis, Elaeocarpus joga, and Serianthes nelsonii trees were studied in northern Guam limestone forests to determine the role of these trees in maintaining spatial heterogeneity of biogeochemistry. The soils beneath S. nelsonii were nutrient-enriched compared to soils away from S. nelsonii. The soils beneath A. mariannensis were depauperate for some nutrients or were not different from the soils away from A. mariannensis for other nutrients. The soils beneath E. joga exhibited increases in some nutrients such as nitrogen, carbon, and phosphorus, but decreases in other nutrients such as potassium and calcium when compared to the soils away from E. joga trees. These three tree species influenced spatial heterogeneity in soil nutrient status in the order A. mariannensis < E. joga < S. nelsonii and their presence added greatly to surface soil heterogeneity. Iron, manganese, and pH exhibited the least variation within the paired sites. Calcium, magnesium, potassium, and zinc exhibited the greatest variation among the paired sites. These findings indicate that continuing loss of these trees from Guam’s forests will diminish the associated biogeochemical spatial heterogeneity.
1. Introduction
Late successional tree species in the forests of the Mariana Island Archipelago have been in a state of decline for many years. Three of these species are Artocarpus mariannensis Trécul, Elaeocarpus joga Merr., and Serianthes nelsonii Merr. The legume S. nelsonii that is endemic to the two southernmost islands of the archipelago, was already noted as rare when it was first described, and is currently listed as critically endangered. [Citation1] The threats to these species are similar to those of the other declining tree species, and efforts are underway to better understand these threats. A failure to regenerate and attrition of the remaining trees due to ongoing anthropogenic activities are some of the relevant conservation concerns. The contemporary forests are characterized by a skewed population age structure toward large, mature individuals. [Citation2–Citation5]
By the time one of the trees of these species occupies the mature emergent forest canopy, it has been influencing the habitat niche for decades, and is therefore ideal for studying the spatial role in ecosystem services. Long-lived tree species may increase ecosystem spatial chemical heterogeneity by orchestrating many biotic and abiotic factors. [Citation6–Citation9] Some of these include chronic extraction and sequestration of soil elements in plant organs, the extent of element resorption prior to organ senescence, litter quality effects on organic matter lability, and local amplification or suppression of soil microorganisms. The direct study of tree-specific soil legacy effects is a subset of literature on the Janzen–Connell model [Citation10,Citation11] and the literature on plant-soil feedbacks. [Citation12] In addition to residual effects on essential nutrients, these fields of study include allelochemicals and pathogens, and collectively may change plant community dynamics and nutrient cycling. While many of the case studies have focused on soils within large mono-specific stands, the influence of individual large canopy trees on biogeochemistry can be substantial, even in biodiverse tree communities. [Citation13,Citation14]
The niche properties of substrates associated with the focal tree species in comparison with nearby substrates away from the tree were determined by collecting soil samples associated with mature trees. My objective was to use established niche chemistry protocols to determine the soil chemical traits beneath the canopy of these three species and to compare these traits to the soils nearby but outside the influence of the focal tree species. Habitats dominated by legume species are known to exhibit soil nutrient relations that differ from non-legume habitats, [Citation15–Citation17] so I predicted the localized differences in soil chemical traits beneath the S. nelsonii tree would be greater in contrast with the general community traits than the other two species.
2. Materials and methods
The 17 study sites were located along the perimeter of the northern calcareous plateau of Guam within a longitudinal range of E 144.814° to 144.929° and a latitudinal range of N 13.464° to 13.649°. The tropical wet climate is Af under the Köppen-Geiger classification. The karst soils formed in slope alluvium, loess, and residuum overlying limestone (Clayey-skeletal, gibbsite, nonacid, isohyperthermic Lithic Ustorthents). [Citation18] The calcareous plateau has a maximum height of 180–185 m above sea level. The forest type has been classified as “typhoon forests” due to tropical cyclone frequency. [Citation19] The close proximity of paired sampling sites ensured homogeneous geological, edaphic, and disturbance histories for each of the 17 paired sites.
Soil samples were collected from the S. nelsonii site in Aug 2015 and from the sites for the other two species in Aug 2017. Soil-sampling methods were patterned after Marler and Krishnapillai. [Citation20] The eight A. mariannensis and eight E. joga trees were mature components of the emergent canopy. Four transects were oriented in cardinal directions for each tree. Two sampling locations along each transect underneath each tree canopy included the location 1 m inside the periphery of the canopy, and half the distance between the bole and the periphery of the canopy. These eight subsamples were combined into one sample for each A. mariannensis and E. joga tree and defined as “home” soils. There were eight trees for these two species. Only one known mature S. nelsonii tree remains on Guam, so the eight home subsamples were not combined for these species and were treated as individual replications. Samples were also collected at fixed distances of 30 m and 60 m on each transect for a total of eight subsamples which were defined as “away” soils. If the prescribed site was within close proximity to a second tree of the same species, the location was moved closer or farther to ensure all away samples were not influenced by the focal species. Treatment of the replications was the same as for home soils. The litter layer was carefully brushed away to expose the mineral layer. One depth increment of 0–10 cm were sampled.
The soil for each replication was separated into three batches. One batch was dried at 50°C then total carbon (C) and nitrogen (N) contents were determined by dry combustion (FLASH EA1112 CHN analyzer; Thermo Fisher, Waltham, Mass., USA). Extractable essential nutrients other than phosphorus (P) were quantified following digestion with diethylenetriaminepentaacetic acid. [Citation21] Analysis was by inductively coupled plasma optical emission spectrometry (Spectro Genesis; SPECTRO Analytical Instruments, Kleve, Germany). Available P was determined by the Olsen method. [Citation22] Soil reaction was determined from saturated paste with an Accumet AB200 meter (Thermo Fisher Scientific, Singapore). A second batch of the fresh soil was analyzed for nitrate and ammonium following 2M potassium chloride extraction. [Citation23] The third batch of fresh soil was incubated in situ for 25 days using the buried bag method. [Citation24] Nitrate and ammonium were quantified at the end of the incubation period.
Several derived variables were calculated. Net nitrification rate was calculated by subtracting initial from final nitrate concentration and dividing by the incubation period. Net mineralization was calculated by subtracting initial nitrate plus ammonium from final nitrate plus ammonium concentration and dividing by the incubation period. Percent available nitrogen was calculated as ((nitrate+ammonium)/total nitrogen)*100. The stoichiometric relationship between carbon and nitrogen was quantified as C/N.
Most of the response variables did not meet parametric requirements primarily because of similarity among home soils and heterogeneity among the away soils. Therefore, the Mann-Whitney U test [Citation25] was employed to determine the differences between home and away soils, as this test does not require any assumption about distribution of the data.
3. Results
3.1. Artocarpus mariannensis
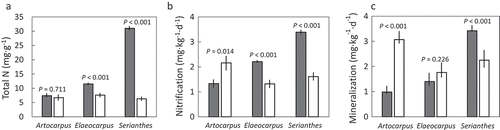
The soils away from mature A. mariannensis trees contained greater concentrations of P, potassium (K), calcium (Ca), magnesium (Mg), and manganese (Mn) than the soils beneath the trees (), according to the Mann-Whitney U test. The soils beneath A. mariannensis trees contained greater concentrations of copper and zinc than soils away from these trees. Ammonium, C, N, iron (Fe), nitrate, and pH were not influenced by sampling location (, ). Copper (Cu) and zinc (Zn) were the only elements that were increased by the presence of an A. mariannensis tree. The derived variables percent available N and C/N were also not influenced by sampling location (). The behavioral variables net nitrification and net mineralization were not influenced by proximity to A. mariannensis trees ().
Table 1. Proximity to Artocarpus mariannensis trees and soil chemical traits in Guam. Home soils were collected underneath mature trees. Away soils were collected 30 m and 60 m from focal trees. Means ± SE, n = 8.
Figure 1. The influence of proximity to Artocarpus mariannensis, Elaeocarpus joga, or Serianthes nelsonii trees on soil chemical traits in karst soils on Guam. Home soils (shaded bars) were collected underneath the trees, and away soils (white bars) were collected 30 m and 60 m from focal trees. (a) Total nitrogen; (b) Net nitrification; and (c) Net mineralization. P-value determined by Mann-Whitney U test.
3.2. Elaeocarpus joga
The soils away from mature E. joga trees contained greater concentrations of Ca, K, Mg, nitrate, and Zn than the soils beneath the trees (), according to the Mann-Whitney U test. The soils beneath E. joga trees contained greater concentrations of ammonium, C, Cu, Fe, and N than soils away from these trees (, ). Mn, P, and pH were not influenced by sampling location (). The derived variables percent available nitrogen and C/N were also not influenced by sampling location (). Net nitrification was greater beneath E. joga trees and net mineralization was not influenced by sampling location ().
Table 2. Proximity to Elaeocarpus joga trees and soil chemical traits in Guam. Home soils were collected underneath mature trees. Away soils were collected 30 m and 60 m from focal trees. Means ± SE, n = 8.
3.3. Serianthes nelsonii
The soils away from mature S. nelsonii trees contained greater concentrations of P than the soils beneath the trees (), according to the Mann-Whitney U test. The soils beneath S. nelsonii trees contained greater concentrations of ammonium, C, Ca, K, Mg, N, and Zn than soils away from these trees (, ). Cu, Fe, Mn, nitrate, and pH were not influenced by sampling location (). The derived variables percent available N and C/N were greater in away soils than in S. nelsonii soils (). The behavioral variables net nitrification and net mineralization were greater beneath S. nelsonii trees than away from the trees ().
Table 3. Proximity to Serianthes nelsonii trees and soil chemical traits in Guam. Home soils were collected underneath the only known mature tree remaining on Guam. Away soils were collected 30 m and 60 m from the tree. Means ± SE, n = 8.
4. Discussion
A contrast in how soils varied beneath A. mariannensis, E. joga, and S. nelsonii trees in Guam’s limestone forests has been described. As predicted, the S. nelsonii trees generated the most profound alterations of habitat biogeochemistry. For example, the concentrations of six of the 10 essential nutrients that were quantified were greater in home soils, and S. nelsonii was the only species with soils that differed for both of the calculated variables of percent available N and C/N. The soils beneath E. joga trees were different from surrounding soils for eight of the 10 essential elements, but the number of nutrients that were greater in home soils was the same as that in away soils. Artocarpus mariannensis trees were associated with soils where all of the important C and N traits were not influenced by sampling location, five of the 10 essential elements were greater in away soils, and only Cu and Zn were greater in home soils. The combined results indicate the magnitude of increased spatial heterogeneity in soil nutrient status for these trees was A. mariannensis < E. joga < S. nelsonii.
Drivers of the spatial changes caused by trees include differences in quality of microsite leaf litter inputs, nutrient inputs from enhanced or reduced animal visitations associated with spatial microsites, nutrients concentrated in stemflow then deposited at the base of individual trees, species-specific vertical and horizontal root contributions, and differential interactions among the tree community with soil microbes. [Citation7,Citation8,Citation26] Leaf litter quality is a function of the ability to absorb and sequester nutrients into leaves throughout their functional life, but also a function of how much of the resident nutrients are re-mobilized and returned to the stems prior to leaf abscission. [Citation27,Citation28] Heterogeneous soil C:N:P relations may profoundly influence the soil microorganism community. [Citation29]
The depauperate soils beneath A. mariannensis indicate a high proportion of leaf nutrients may be re-mobilized prior to leaf senescence in these species, and the tree may benefit edaphic consumer guilds with slow-growing conservation growth strategies. The nutrient-rich soils beneath S. nelsonii indicate greater leaf nutrient concentrations and lower nutrient re-mobilization during leaf senescence, and the tree may benefit edaphic consumer guilds with fast-growing exploitative growth strategies.
The difference in home and away soils occurred for all three tree species for Ca, K, Mg, and Zn. In contrast, this difference occurred for only one of the three tree species for Fe and Mn. The spatial stability of pH among sites is a function of the buffering capacity for this soil series. The lower horizons are almost pure calcium carbonate. [Citation18]
Studying the abiotic traits of soils that are under the influence of Guam’s sessile, long-lived tree species may improve our ability to conserve the forests and more fully understand the ecosystem services these trees provide. The increased spatial heterogeneity of biogeochemical traits that is delivered by these trees may have profound and lasting influences on soil food web biodiversity in these typhoon forests. This subject deserves further study. More importantly, the increased knowledge provides greater insight into the detrimental effects that are occurring with ongoing losses of these late successional trees due to anthropogenic disturbances.
Disclosure of Potential Conflicts of Interest
No potential conflict of interest were disclosed.
Acknowledgments
This research received no external funding. No take was associated with Serianthes nelsonii. Methods and results for Serianthes nelsonii approved by the Environmental Flight, Andersen Air Force Base.
References
- Wiles G, Williams E. Serianthes nelsonii. The IUCN Red List of Threatened Species. 2017: e.T30437A98715973. doi:10.2305/IUCN.UK.2017-3.RLTS.T30437A98715973.en.
- Raulerson L, Rinehart A. Trees and shrubs of the Northern Mariana Islands. Saipan, Commonwealth of the Northern Mariana Islands: Coastal Resources Management; 1991.
- Wiles GJ, Schreiner IH, Nafus D, et al. The status, biology, and conservation of Serianthes nelsonii (Fabaceae), an endangered Micronesian tree. Biol Conserv. 1996;76:229–239.
- Schreiner IH. Demography and recruitment of selected trees in the limestone forest of Guam in relation to introduced ungulates. Micronesica. 1997;30:169–181.
- Ritter MW, Naugle CM. Population characteristics, germination and proposed management of Elaeocarpus joga Merr. On Guam: a Regionally Endemic Tree Micronesica. 1999;31:275–281.
- Van Der Heijden MG, Bardgett RD, Van Straalen NM. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett. 2008;11:296–310.
- Van der Putten WH, Bardgett RD, Bever JD, et al. Plant-soil feedbacks: the past, present and future challenges. J Ecol. 2013;101:265–276.
- Dawud SM, Raulund-Rasmussen K, Ratcliffe S, et al. Tree species functional group is a more important driver of soil properties than tree species diversity across major European forest types. Funct Ecol. 2017;31:1153–1162.
- Teste FP, Kardol P, Turner BL, et al. Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science. 2017;355:173–176.
- Janzen DH. Herbivores and the number of tree species in tropical forests. Am Natur. 1970;104:501–529.
- Connell JH. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In: Den Boer PJ, Gradwell GR, editors. Dynamics of populations. Wageningen, Netherlands: Centre for agricultural publishing and documentation; 1971. p. 298–312.
- Kulmatiski A, Beard KH, Stevens JR, et al. Plant soil feedbacks: A meta-analytical review. Ecol Lett. 2008;11:980–992.
- Andivia E, Fernández M, Alejano R, et al. Tree patch distribution drives spatial heterogeneity of soil traits in cork oak woodlands. Ann For Sci. 2015;72:549–559.
- Kooch Y, Tarighat FS, Hosseini SM. Tree species effects on soil chemical, biochemical and biological features in mixed Caspian lowland forests. Trees. 2017;31:863–872.
- Chimphango SBM, Potgieter G, Cramer MD. Differentiation of the biogeochemical niches of legumes and nonlegumes in the Cape Floristic Region of South Africa. Plant Ecol. 2015;216:1583–1595.
- Marler TE, Dongol N, Cruz GN. Leucaena leucocephala and adjacent native limestone forest habitats contrast in soil properties on Tinian Island. Communic Integr Biol. 2016;9:e1212792.
- Marler TE. Soil chemistry following afforestation of barren coastal soils in Southern Guam does not conform to that of continuously vegetated surfaces. J Coastal Zone Management. 2017;20:2.
- Young FJ. Soil survey of Territory of Guam. U. S. Dept. of Agric. Soil Conservation Service; Washington DC, USA; 1988.
- Stone BC. America’s Asiatic flora: the plants of Guam. Am Sci. 1971;59:308–319.
- Marler TE, Krishnapillai MV. Cycas micronesica trees alter local soil traits. Forests. 2018;9:565.
- Berghage RD, Krauskopf DM, Warncke DD, et al. Micronutrient testing of plant growth media extractant, identification and evaluation. Commun Soil Sci Plant Anal. 1987;18:1089–1109.
- Olsen S, Cole C, Watanabe F, et al. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. USDA Circular Nr 939, US Gov Print Office, Washington DC, U.S.A.; 1954.
- Cataldo DA, Haroon M, Schrader LE, et al. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun Soil Sci Plant Anal. 1975;6:71–80.
- Eno CF. Nitrate production in the field by incubating the soil in polyethylene bags. Proc Soil Sci Soc Amer. 1960;24:277–279.
- Mann HB, Whitney DR. On a test of whether one of two random variables is stochastically larger than the other. Ann Math Stat. 1947;18:50–60.
- Ehrenfeld JG, Ravit B, Elgersma K. Feedback in the plant soil system. Annu Rev Environ Resour. 2005;30:75–115.
- Killingbeck KT. Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology. 1996;77:1716–1727.
- Killingbeck KT. Nutrient resorption. In: Noodén LD, editor. Plant cell death processes. Amsterdam: Elsevier; 2004. p. 215–226.
- Shen F, Wu J, Fan H, et al. Soil N/P and C/P ratio regulate the responses of soil microbial community composition and enzyme activities in a long-term nitrogen loaded Chinese fir forest. Plant Soil. 2019;436:91–107.