
Abstract
Although desalination by membrane processes is a possible solution to the problem of freshwater supply, related cost and energy demands prohibit its use on a global scale. Hence, there is an emerging necessity for alternative, energy and cost-efficient methods for water desalination. Cyanobacteria are oxygen-producing, photosynthetic bacteria that actively grow in vast blooms both in fresh and seawater bodies. Moreover, cyanobacteria can grow with minimal nutrient requirements and under natural sunlight. Taking these observations together, a consortium of five British Universities was formed to test the principle of using cyanobacteria as ion exchangers, for the specific removal of Na+ and Cl− from seawater. This project consisted of the isolation and characterisation of candidate strains, with central focus on their potential to be osmotically and ionically adaptable. The selection panel resulted in the identification of two Euryhaline strains, one of freshwater (Synechocystis sp. Strain PCC 6803) and one of marine origin (Synechococcus sp. Strain PCC 7002) (Robert Gordon University, Aberdeen). Other work packages were as follows. Genetic manipulations potentially allowed for the expression of a light-driven, Cl−-selective pump in both strains, therefore, enhancing the bioaccumulation of specific ions within the cell (University of Glasgow). Characterisation of surface properties under different salinities (University of Sheffield), ensured that cell–liquid separation efficiency would be maximised post-treatment, as well as monitoring the secretion of mucopolysaccharides in the medium during cell growth. Work at Newcastle University is focused on the social acceptance of this scenario, together with an assessment of the potential risks through the generation and application of a Hazard Analysis and Critical Control Points plan. Finally, researchers in Imperial College (London) designed the process, from biomass production to water treatment and generation of a model photobioreactor. This multimodal approach has produced promising first results, and further optimisation is expected to result in mass scaling of this process.
1. Introduction
In the year 2014, we are reaching the fulfilment of the “International Decade for Action WATER FOR LIFE—2005–2015” [Citation1]. One of the milestones of this project was to reduce by half people’s “sustainable access to Safe Drinking Water and Basic Sanitation” [Citation2]. At the same time, extreme weather events have increased the occurrence of droughts, most notably in arid regions in close proximity to the sea [Citation3,4]. In addition, developing countries are more prone to these effects [Citation5] and these factors put together allowed for the conception of “Biodesalination: From cell to tap” [Citation6]. This project envisages the utilisation of photosynthetic bacteria for the removal of specific ions from seawater, thus, providing a sustainable alternative to seawater desalination, when compared to current methods [Citation7,8].
The use of photosynthetic bacteria in the direct removal of Na+ has been previously demonstrated. For example, cyanobacteria, or fractions that they secrete in response to salt stress, have been shown to remove Na+ from aqueous solutions and soils, as well as enhancing seed germination rates [Citation9,10]. This in turn generated the first challenge in the “Biodesalination” scenario. In order to this process to be efficient, bacterial growth should supply the large volumes of water that need to be treated [Citation11]. Furthermore, biomass should be grown in a sustainable manner, utilising solar energy and the minimal addition of nutrients.
Algae could be used as alternatives to wastewater remediation, by sequestering CO2 and by capturing phosphates and nitrates in a form that can be further utilised. The disadvantages of using wastewater as a nutrient source are the presence of pathogens, therefore, limiting its use to sanitation and irrigation. With respect to cyanobacteria, use of wastewater is not a necessary requirement, due to their natural ability to form blooms in natural waters [Citation12,13]. Further investigation in the nutrient requirements of cyanobacteria, demonstrated that the “Redfield Ratio” (C:N:p = 106:16:1) allowed for optimal growth, while the equivalent ratio under light limitation was C:N:p = 117:25:1 [Citation14,15].
Understanding sodium homeostasis within a biological system was the next step in the process. Upon the exertion of ionic stress, cyanobacteria utilise a number of energy-demanding processes in order to export Na+ [Citation16,17]. In a “biodesalination” scenario, it is important that cells were starved of energy, therefore, halting active export of ions. Previous observations demonstrated that cyanobacteria accumulate osmolytes intra- or extra-cellularly in response to ionic stress [Citation18]. The compatible solutes accumulated by cyanobacteria also show hierarchy in their protective potential against salt stress [Citation19]. Furthermore, cyanobacteria are also capable of autofermentation. That is to say, they can catabolise internally stored carbohydrates, including osmolytes, in the dark [Citation20]. Taken together, these observations suggested that an energetically favourable way of growing cyanobacteria was administration of just sufficient light and nutrients for rapid growth, while allowing each cell enough energy (in the form of sugars) to counteract the stress exerted by salt. In addition, cells should be just depleted of energy and therefore not able to actively export Na+.
As discussed in a recently published case study on using cyanobacteria for biodesalination [Citation6], one of the bottlenecks in the production process is the efficient removal of the salt laden cells from the desalinated water without affecting the integrity of the cell membrane. Cell–liquid separation techniques such as sedimentation or flotation require, as a first step, the formation of robust cell aggregates. Research suggests that use of conventional metal salts as coagulants to trigger aggregate formation leads to widespread cell death and lysis during the separation process [Citation6]. Therefore, a thorough investigation of the cell surface characteristics and the alternative use of biopolymers as coagulants is merited.
This study was an examination of the potential of using biological systems for the large-scale desalination of seawater. This manuscript addresses: (1) the screening methods employed for the selection of micro-organisms suitable for this application. These included both literature searches and a biotechnological approach on strain screening and subsequent growth optimisation, (2) the identification of an analytical method capable of specifically detecting [Na+] and [Cl−] in the range between 10 and 100 mM and (3) the identification of the cell surface macromolecules on selected strains and an investigation of the aggregation potential based on physicochemical parameters of the cell surface and the extended Derjaguin, Landau, Verwey and Overbeek (XDLVO) theory, a classical theory of colloidal stability.
2. Materials and methods
2.1. Strain selection from literature
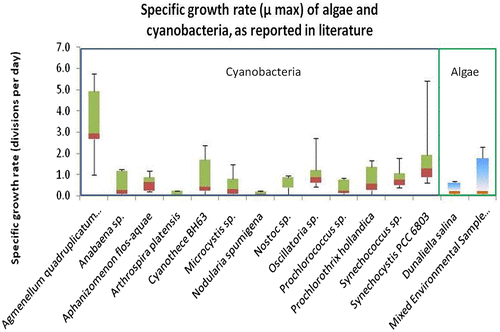
Strain selection was initiated with the literature screening for fast-growing algae and cyanobacteria. This was done in order to comply with the project’s requirements for a fast production of biomass. Results of this initial screening can be found in Fig. , while literature reviewed in this respect can be found in Appendix 1. In addition, the nutrient requirements for different strains were reviewed, together with the possibility of using different wastewaters for the supply of nutrients in the form of phosphates and nitrates. Finally, the effect of environmental conditions, such as temperature and light regime was also evaluated.
Fig. 1. Literature searches indicating the specific growth rate of different cyanobacteria, algae and mixed environmental samples. In the figure above, it is evident that cyanobacteria appeared to have overall higher growth rates, when compared to algae.
2.1.1. Growth optimisation of selected strains
Upon considering the growth parameters reviewed above, two euryhaline strains were selected for the practical screening and potential application in “biodesalination”. These were Synechocystis sp. PCC 6803 and Synechococcus sp. PCC 7002 [Citation21,22]. Growth of both strains was monitored in Erlenmeyer flasks containing BG11 [Citation23] or BG-11 Seawater (BG11-SW) for Synechococcus PCC 7002. BG11-SW was made by dissolving stock solutions of BG11 salts in filtered seawater (Stonehaven, Aberdeenshire, UK). Standard laboratory light was set at 25 μmol m−2 s−1, and ambient temperature was 22 ± 1°C, unless otherwise stated. Cells used for the inoculum were 3 weeks old, and cell numbers were adjusted to 2.5 million cells after inoculating 100 ml of sterile BG11. Light was provided by Osram Cool White lamps, while the effects of the duration of light (12:12, L:D or continuous light) and the effects of light intensity (25, 50, and 95 μmol m−2 s−1) were evaluated in a Fitotron growth chamber (Weiss Gallenkamp). Finally, the combination of effects of different temperatures in the growth of these strains was evaluated by incubating respective Erlenmeyer flasks containing each culture at mid-log and stationary phase (5 and 21 d following inoculation) in water baths at either 22 or 28°C.
2.2. Cell enumeration, viability and size estimation
Cell Numbers were evaluated by flow cytometry, using the Coulter Epics-XL-MCL (Beckman Coulter). Separation of live and dead cells was performed by distinguishing the emissions detected in the phycoerythrin and the fluorescein isothiocyanate channels. Size estimation (1–15 μm) was possible by setting up the Forward Scattering detection using the Flow Cytometry Size Calibration Kit (life Technologies).
2.3. Analytical chemistry
High-performance Liquid Chromatography was performed using the Waters Alliance 2695 Separations Module with the ZIC-HILIC, PEEK column (150 × 2.1 μm, 3.5 μm, 200 Å). Gradient separation of anions and cations was performed using the method described in the SeQuant Application Note 2700–41A, with 20 mM ammonium acetate (pH 3.0 with acetic acid) instead of ammonium formate. HPLC detection was performed using the Biotage ELSD-1080. The detection parameters were: nebuliser and evaporator temperature equal to 40°C, air-free nitrogen flow set at 2.5 slm. Chromatograms were acquired by Masslynx 4.0.
Standards were prepared by serial dilutions of freshly prepared 2 M NaCl (HPLC grade, Fisher) solution in MilliQ water. Next, this stock solution was further diluted (1:10) in 80:20 Acetonitrile: 20 mM ammonium acetate (pH 3.0) to final concentrations ranging from 1 to 150 mM. 10 μL of each standard and three blanks were run in triplicates.
2.4. Statistical analysis
Statistical analysis of acquired data was performed using Repeated Measurements ANOVA in SPSS (v21).
2.5. X-ray photoelectron spectroscopy
The X-ray photoelectron spectroscopy (XPS) analysis was carried out as described by previously [Citation30]. Synechocystis PCC 6803 was grown in BG-11 medium [Citation23] and Synechococcus PCC 7002 was grown in A + medium [Citation22] at 22°C in Erlenmeyer flasks with agitation at 150 rpm for adequate aeration. The light was set to 40 μE m−2 s−1 with a 12:12 light to dark cycle. The cells were harvested for 3 weeks post-inoculation by centrifugation at 4,500 g for 20 min at 4°C. The harvested cells were washed two times in distilled water and freeze-dried before analysis. The measurements were carried out on a KRATOS AXIS 165 Ultra photoelectron spectrometer at 10 kV and 20 mA using the AL Kα X-ray source (1,486.6 eV). The take-off angle was adjusted at 90° and data were collected for each sample (two biological replicates) at three randomly selected location (three technical replicates). The area corresponding to each sample was 400 μm in diameter. A survey scan was carried out (pass energy 20 eV, 0.1 eV step size) for C, O and N followed by a high-resolution scan (pass energy 20 eV, 0.1 eV step size) for C and O. The binding energies were normalised using the C1s peak at 284.6 eV. Casa XPS software (version 2.3.16) was used to carry out the spectral integration.
2.6. XDLVO modelling of aggregation
The XDLVO model was carried out as described previously [Citation31]. Briefly, the electrophoretic mobility of the cells was measured in 100 mM KCl at pH 8 using a Zeta potential analyser (Brookhaven Instruments, UK) [Citation30]. The measured electrophoretic mobilities were converted into apparent zeta potentials assuming the conditions for Helmholtz–Smoluchowski relationship are met. The contact angle of three liquids: water, formamide and diiodomethane with the bacterial lawns was measured using a tensiometer (Attension, UK) after drying the bacterial lawns until the plateau phase of drying is reached [Citation31]. Measured contact angles were converted into surface tension using the LW-AB approach [Citation32]. The interfacial free energy of aggregation of cells in an aqueous medium was calculated based on a sphere–sphere configuration [Citation32].
3. Results and discussion
3.1. Overview of the growth parameters
Upon combining this information and understanding the practical applications of “Biodesalination”, it was decided that growth optimisation of selected strains should conform to the following parameters. Initially, a temperature between 20 and 40°C would allow for the utilisation of these organisms in a large number of countries, where “biodesalination” could be applied. A pH range between 6 and 9 will facilitate the growth of most photosynthetic organisms researched to date. A salinity range of 0–0.5 M (NaCl), or 0–10 g L−1 of Na+, was considered to also reflect the possible range of salinities encountered over the utilisation of cyanobacteria for biodesalination. A light–dark cycle of 12:12 was considered to be a close approximation of the annual light availability in tropical countries. In respect to the nutritional requirements of the two selected organisms, natural seawater should be supplemented with 0–0.007 g L−1 phosphates (equivalent of 0–0.04 g L−1 K2HPO4) and 0–0.25 g L−1 nitrates (equivalent to 0–1.5 g L−1 NaNO3).
Based on some preliminary calculations, a moderate estimate of bacterial cell densities achieved to date (data not shown) was expected to be in the region of 1014–1015 cells L−1. With ion-transport proteins present at typical densities, one cell could transport approximately 107 of each ion per second across the membrane barrier. Therefore, 1 L of cell volume should be sufficient for the desalination of 2 L of water in 10 min. This would equate to a reduction of [Na+] and [Cl−] to 5% of the original seawater concentration.
3.2. Growth optimisation under standard laboratory conditions
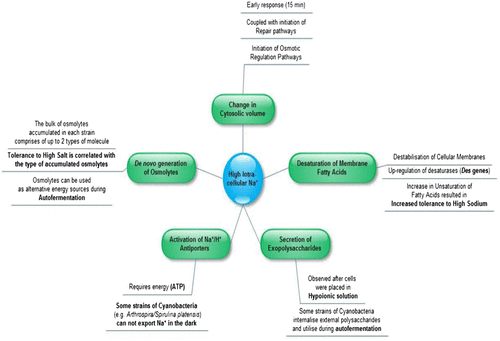
After setting the experimental parameters regarding the practicalities of “biodesalination”, two strains were selected from the literature, according to their ability to tolerate a wide array of different salinities [Citation21,22]. The reasoning behind this was that the selected strain would be exposed to 0–0.5 M NaCl, throughout its growth and during water treatment. The organisms originally selected included a cyanobacterial strain of freshwater origin (Synechocystis sp. Strain PCC 6803) and one of a marine background (Synechococcus sp. Strain PCC 7002, formerly Agmenellum quadruplicatum PR-6). Both strains have already been widely used as model organisms for understanding cyanobacterial photosynthesis and adaptation to ionic stress, and their genomes are fully sequenced [Citation24–27]. A general overview of the physiological adaptations of different cyanobacterial strains to salt stress (immediate transfer to a hyperionic or hypoionic solution) can be found in Fig. .
Fig. 2. Physiological adaptations of cyanobacteria to ionic stress caused by Na+. The mechanisms described above give a brief overview of early responses, as well as long-term adaptations to high salinity.
In order to achieve maximum growth of both organisms, a previously optimised medium, BG11, was used. BG11 is an affordable, eutrophic medium, widely used for laboratory experiments [Citation23]. Recipe stock solutions of minerals were either added to distilled water (BG11) or to filtered seawater from Stonehaven, Aberdeenshire (BG11-SW). An overview of the growth of both strains in these media can be found in Figs. and and Table . It is worth mentioning that when compared with data from the literature (Fig. ) the specific growth observed under laboratory conditions was lower than previously reported for Synechococcus PCC 7002. On the other hand, the specific growth rate of Synechocystis PCC 6803 was comparable with previously reported findings. Moreover, Synechocystis PCC 6803 grew faster under stronger light (95 μmol m−2 s−1), with both continuous and intermittent illumination. Synechococcus PCC 7002, demonstrated better overall growth under low light intensities (25 μmol m−2 s−1). Intermittent light appeared to be beneficial for overall growth in both strains, probably because it allowed time for light-independent reactions of photosynthesis and/or facilitated carbon concentrating mechanisms to capture CO2 and enhance photosynthesis [Citation28,29].
Fig. 3(a). Effect of intermittent light in the growth of selected cyanobacteria. Data indicated that Synechococcus PCC 7002 was growing faster than Synechocystis PCC 6803 in all light intensities tested. In contrast, Synechocystis PCC 6803 appeared to show better overall growth under high light intensities, when compared to Synechococcus PCC 7002. Error bars indicate standard deviation (n = 3).
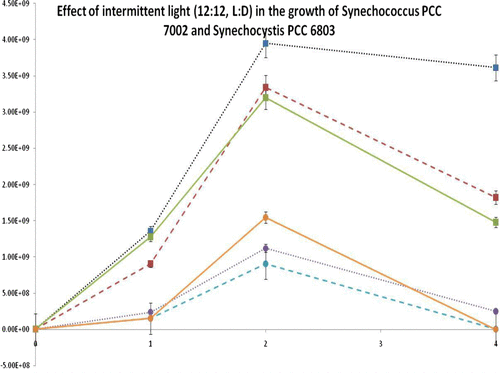
Fig. 3(b). Effect of continuous light in the growth of selected cyanobacteria. Both Synechococcus PCC 7002 and Synechocystis PCC 6803 appeared to reach maximum growth on day 21 (week 3). Moreover, Synechococcus PCC 7002 grew best at lower light intensities, while Synechocystis PCC 6803 showed faster growth under high light intensities. It is worth noting that the maximum cell densities achieved was ten-fold lower than when grown under intermittent light. Error bars indicate standard deviation (n = 3).
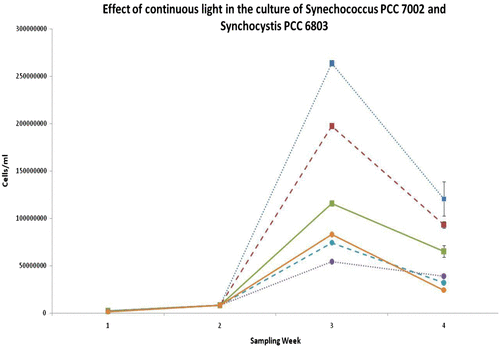
Table 1 Divisions per day as calculated by flow cytometry. The table below demonstrates growth rate (per day) for Synechococcus PCC 7002 and Synechocystis PCC 6803, under different illuminations. Red indicates highest growth observed for each organism over 3 weeks of culture. Green indicates lowest growth for each organism per period.
With the aim to meet the growth requirements of either strain, the availability of phosphates and nitrates in natural seawater [Citation30] needed to be assessed. Further supplementations of growth media may enhance sustainability and remediation potential of this process. A literature review revealed the Dissolved Organic Nitrate/Dissolved Inorganic Phosphate ratio in the Atlantic Ocean [Citation28], and the availability of phosphates and nitrates in different wastewaters. In brief, Dairy Lagoon Water was found to have a ratio of C:N = 3.5:1, C:p = 10:1, while domestic sewage has the following profile: C:N = 3.5:1; C:p = 20:1 [Citation29].
3.3. HPLC detection of Na+ and Cl−
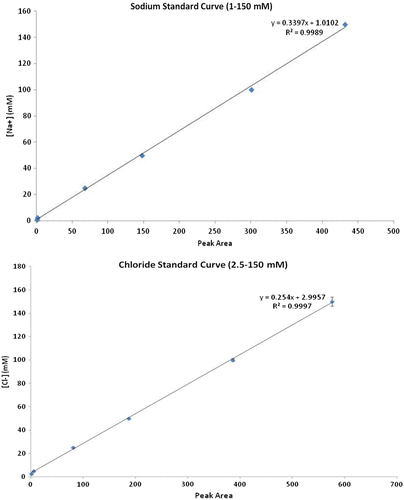
HPLC detection of Na+ and Cl− in water allowed for a detection range between 1 and 150 mM for both ions (Fig. ). On the other hand, large variation in statistical replicates (n = 3), below 10 mM and above 100 mM, suggested that our quantification range is within those limits (Appendix 2).
Fig. 4. HPLC detection standard curve for Na+ and Cl−. The limits of quantification for both equations were limited between 10 and 100 mM. Error bars indicate standard deviation (n = 3).
3.4. Cell–Water separation
Given the absolute requirement for a gentle yet effective method for the formation of cell aggregates during the cell–water separation stage, it was decided to study the natural cell–cell aggregation mechanisms in the two selected strains with a view to subsequently engineer these natural aggregation mechanisms for effective cell–water separation. Aggregation in bacteria is mediated by physicochemical forces such as van der Waals forces, electrostatic forces and acid–base interactions that arise from the macromolecules on the cell surface [Citation31]. Therefore, changing the cell surface macromolecular display can be an effective route to engineering aggregation in bacteria. As a preliminary step, in this study, the macromolecular display on the cell surface of the two selected strains, Synechocystis PCC 6803 and Synechococcus PCC 7002, was studied using XPS and the aggregation of each of the strains was modelled as particle–particle physicochemical interactions and described using the XDLVO theory.
3.5. Study of the cell surface macromolecules using XPS
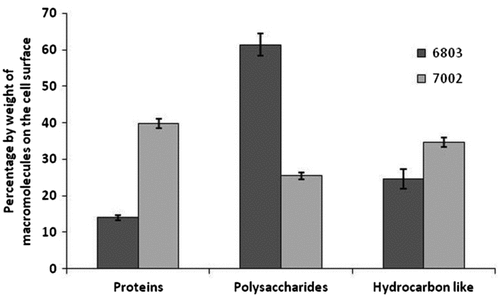
XPS analysis can identify the nature of the macromolecules on the cell surface of bacteria [Citation33]. It is a surface sensitive technique because the photoelectrons emerging from the sample, upon irradiation by X-rays, undergo inelastic scattering within the sample and therefore, only the photoelectrons from the surface of the sample will be detected. Typically, the information obtained by XPS can be attributed to arise from a depth of 8–10 nm on the cell surface [Citation30]. Synechococcus PCC 6803 and Synechocystis PCC 7002 were subjected to XPS analysis. The results (Fig. ) suggest that the cell surfaces of the two selected strains are very different. The surface of Synechocystis PCC 6803 is rich in polysaccharides whilst the surface of Synechococcus PCC 7002 predominantly contains proteins. The difference in the functional groups present on the cell surface will impact cell–cell interactions during aggregation.
Fig. 5. XPS analysis of Synechocystis PCC 6803 and Synechococcus PCC 7002. Values denote the distribution of macromolecules on the cell surface in the analysed sample volume. Error bars are standard deviation values.
3.6. XDLVO model of aggregation
The DLVO theory suggests that the aggregation is a summative consequence of the attractive van der Waals interactions and the attractive or repulsive electrostatic interactions. The extended DLVO theory (XDLVO) includes the effects of the short-range acid–base interactions arising from polar cell surface moieties, in addition to the forces considered in the DLVO theory [Citation31]. The result of the XDLVO model is a plot of the interaction energy versus the distance of approach between the particles. The XDLVO plot for Synechococcus PCC 6803 and Synechococcus PCC 7002 is given in Fig. . It is apparent from the model for both the strains that, as two cells approach each other, they have to actively overcome a significant energy barrier of greater than 1,000 kT before irreversible aggregation in the primary minimum can be realised. Therefore, the XDLVO model predicts that irreversible aggregation is not possible in either strains.
Fig. 6. XDLVO analysis. Aggregation energies as a function of separation distance for (A) Synechocystis PCC 6803 and (B) Synechococcus PCC 7002. G—interaction energy, GLW—interaction energy due to Lifschitz–van der Waals component, GEL—interaction energy due to electrostatic component, GAB—interaction energy due to acid–base component, GTOT—total interaction energy. Insets show the formation of the predicted secondary minimum at which reversible cell aggregation might occur.
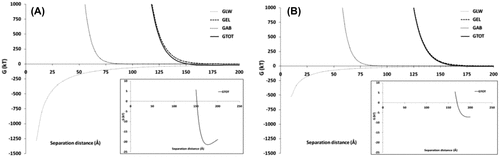
However, in Synechocystis PCC 6803, the model predicts the occurrence of reversible aggregation in the secondary minimum, with a net attractive interaction energy of 22 kT, when the cells are at a distance of approximately 20 nm from each other. The net attractive interaction energy at this distance occurs due the predominance of the van der Waals forces arising from the body of the cells and the virtual absence of the repulsive electrostatic and acid–base forces from the cell surface moieties. A similar reversible aggregation in the secondary minimum is not predicted for Synechococcus PCC 7002. From the model, it is likely that Synechocystis PCC 6803 might favour aggregate formation over Synechococcus PCC 7002. However, the model remains to be validated by experimental means.
4. Conclusions
This manuscript explores the possibility of using biological systems for the targeted removal of Na+ and Cl− from seawater. Utilisation of photosynthetic organisms in that respect allow for the concept of a cost and energy-efficient desalination, when compared to current alternatives. Upon consideration of the practicality of this scenario, a range of environmental and growth parameters were set and two suitable organisms were selected. These include a cyanobacterium of freshwater (Synechocystis PCC 6803) and one of marine origin (Synechococcus PCC 7002). Growth was monitored under standard laboratory conditions and optima were defined for each cyanobacterium. Furthermore, characterisation of surface properties and electrophoretic mobilities facilitated modelling of cell aggregation. The latter allows for the efficient removal of robust cells post-treatment, possibly without the addition of biopolymers as coagulants. Finally, development of an analytical protocol that allows for the detection of traces of Na+ and Cl−, will allow us to further scrutinise ion mobility between saline media and viable cells.
In this respect, tighter monitoring of the physiological changes due to ionic stress would further enhance the applicability of our process. Thus, it is very important that the effects of environmental conditions and nutrient availability are further researched in the biodesalination scenario. Namely, further investigation on how changes in salinity affect cell growth and viability. Could environmental or nutrient “switches” be used in large-scale water treatment? Literature studies and experimental data, suggest that use of biological systems could be a viable alternative to current desalination methods. On the other hand, there are still some challenges to be met. Deepening our understanding of biological processes will allow us to scale-up cyanobacterial growth, meeting the demands of water treatment scenarios. In addition, further examination of cell–medium separation methods in our experimental system will improve our prospect of using biological systems in seawater desalination.
Acknowledgements
The authors of this manuscript would like to thank the Engineering and Physical Sciences Research Council (EPSRC) for funding this “proof of principle” project.
Notes
Presented at the Conference on Desalination for the Environment: Clean Water and Energy 11–15, May 2014, Limassol, Cyprus
References
- International Decade for Action Water for Life 2005–2015: Available from: http://www.un.org/waterforlifedecade World Water Day web-site. Available from: www.unwater.org/wwd07. UN-Water web site. Available from: www.unwater.org. FAO web-site. Available from: www.fao.org. Available from: WHO Water, Sanitation and Health web-site. www.who.int/water_sanitation_health.
- A. Prüss-Üstün, R. Bos, F. Gore, J. Bartram, Safer Water, Better Health: Costs, Benefits and Sustainability of Interventions to Protect and Promote Health, World Health Organisation, Geneva, 2008.
- S. Dasgupta, B. Laplante, C. Meisner, D. Wheeler, J. Yan, A.D. Khawaji, I.K. Kutubkhanah, J.-M. Wie, The impact of sea level rise on developing countries: A comparative analysis, Clim. Change 93 (2009) 379–388.10.1007/s10584-008-9499-5
- J. Romm, Desertification: The next dust bowl, Nature 478 (2011) 450–451.10.1038/478450a
- M. Monirul, Q. Mirza, Climate change and extreme weather events: Can developing countries adapt? Clim. Policy 3 (2003) 233–248.
- J.M. Amezaga, A. Amtmann, C.A. Biggs, T. Bond, C.J. Gandy, A. Honsbein, E. Karunakaran, L. Lawton, M.A. Madsen, K. Minas, M.R. Templeton, Biodesalination: A case study for applications of photosynthetic bacteria in water treatment, Plant Physiol. 164 (2014) 1661–1676.
- M. Elimelech., W.A. Phillip, The future of seawater desalination: Energy, technology, and the environment, Science 333 (2011) 712–717.10.1126/science.1200488
- R.L. McGinnis, M. Elimelech, Energy requirements of ammonia-carbon dioxide forward osmosis desalination, Desalination 207 (2007) 370–382.10.1016/j.desal.2006.08.012
- M. Arora, A. Kaushik, N. Rani, C.P. Kaushik, Effect of cyanobacterial exopolysaccharides on salt stress alleviation and seed germination, J. Environ. Biol. 31 (2010) 701–704.
- N.K. Singh, D.W. Dhar, Cyanobacterial reclamation of salt-affected soil, in: E. Lichtfouse (Ed.), Genetic Engineering, Biofertilisation, Soil Quality and Organic Farming, first ed., vol. 4, Springer, Dijon, pp. 245–249, 2010
- M. Henze, P. Harremoës, J. Cour Jansen, E. Arvin, Wastewater Treatment, Springer, pp. 11–28, 2002.10.1007/978-3-662-04806-1
- D. Justic, N.N. Rabalais, E. Turner, Stoichiometric nutrient balance and origin of coastal eutrophication, Mar. Pollut. Bull. 30 (1995), 41–46.
- S.I. Rubin, T. Takahashi, D.W. Chipman, J.G. Goddard, Deep sea research part I: Oceanographic Research Papers, 45 (1998) 1211–1234.10.1016/S0967-0637(98)00021-1
- A.C. Redfield, The biological Control of chemical factors in the environment, Am. Sci. 46 (1958) 205–221.
- W.S. Broecker, G.M. Henderson, The sequence of events surrounding Termination II and their implications for the cause of glacial-interglacial CO2 changes, Paleoceanography 13 (1998) 352–364.10.1029/98PA00920
- N. Empadinhas, M.S. da Costa, Osmoadaptation mechanisms in prokaryotes: Distribution of compatible solutes, Int. Microbiol. 11 (2008) 151–161.
- H. Gimmler, Primary sodium plasma membrane ATPases in salt-tolerant algae: Facts and fictions, J. Exp. Bot. 51 (2000) 1171–1178.10.1093/jexbot/51.348.1171
- E. Blumwald, R.J. Mehlhorn, L. Packer, Ionic osmoregulation during salt adaptation of the Cyanobacterium Synechococcus 6311, Plant Physiol. 73 (1983) 377–380.10.1104/pp.73.2.377
- R.H. Reed, L.M. Borowitzka, M.A. Mackay, J.A. Chudek, R. Foster, S.R.C. Warr, D.J. Moore, W.D.P. Stewart, Organic solute accumulation in osmotically stressed cyanobacteria, FEMS Microbiol. Lett. 39 (1986) 51–56.10.1111/fml.1986.39.issue-1-2
- D. Carrieri, D. Momot, I.A. Brasg, G. Ananyev, O. Lenz, D.A. Bryant, G.C. Dismukes, Boosting autofermentation rates and product yields with sodium stress cycling: Application to production of renewable fuels by cyanobacteria, Appl. Environ. Microbiol. 76 (2010) 6455–6462.10.1128/AEM.00975-10
- D.L. Richardson, R.H. Reed, W.D.P. Stewart, Synechocystis PCC6803: A euryhaline cyanobacterium, FEMS Microbiol. Lett. 18 (1983) 99–102.10.1111/fml.1983.18.issue-1-2
- Ma. Ludwig, D.A. Bryant, Synechococcus sp. Strain PCC 7002 Transcriptome: Acclimation to temperature, salinity, oxidative stress and mixotrophic growth conditions, Front. Microbiol. 3, (2012), 1–14.
- R.Y. Stanier, R. Kunisawa, M. Mandel, G. Cohen-Bazire, Purification and properties of unicellular blue-green alga (Order Chroococcales), Becteriol. Rev. 35 (1971) 171–205.
- S. Fulda, S. Mikkat, F. Huang, J. Huckauf, K. Marin, B. Norling, M. Hagemann, Proteome analysis of salt stress response in the cyanobacterium Synechocystis sp. strain PCC 6803, Proteomics 6 (2006) 2733–2745.10.1002/(ISSN)1615-9861
- K. Marin, Y. Kanesaki, D.A. Los, I. Suzuki, M. Hagemann, Gene expression profiling reflects physiological processes in salt acclimation of Synechocystis sp. Strain PCC 6803, Plant Physiol. 136 (2004) 3290–3300.10.1104/pp.104.045047
- S.I. Allakhverdiev, A. Sakamoto, Y. Nishiyama, Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II in Synechococcus sp., Plant Physiol. 123 (2000) 1047–1056.10.1104/pp.123.3.1047
- S.C. Singh, P. Sinha, D.P. Häder, Role of lipids and fatty acids in stress tolerance in cyanobacteria, Acta Protozool. 41 (2002) 297–308.
- M. Vidal, C.M. Duarte, S. Agustí, Dissolved organic nitrogen and phosphorus pools and fluxes in the Central Atlantic Ocean, Limnol. Oceanogr. 44 (1999) 106–115.10.4319/lo.1999.44.1.0106
- I. Woertz, A. Feffer, T. Lundquist, Y. Nelson, Algae grown on dairy and municipal wastewater for simultaneous nutrient removal and lipid production for biofuel feedstock, J. Environ. Eng. 135 (2009) 1115–1122.10.1061/(ASCE)EE.1943-7870.0000129
- J. Mukherjee, E. Karunakaran, C.A. Biggs, Using a multifacteted approach to determine the changes in bacterial cell surface properties influenced by a biofilm lifestyle, Biofouling 28 (2012) 1–14.
- T. Das, B.P. Krom, H.C. van der Mei, H.J. Busscher, P.K. Sharma, DNA-mediated bacterial aggregation is dictated by acid—Base interactions, Soft Matter 7 (2011) 2927–2935.10.1039/c0sm01142h
- P.K. Sharma, H. Rao, Adhesion of Paenibacillus polymyxa on chalcopyrite: Surface thermodynamics and extended DLVO theory, Colloids Surf. B: Biointerfaces 29 (2003) 21–38.
- P.G. Rouxhet, N. Mozes, P.B. Dengis, Y.F. Dufrêne, P.A. Gerin, M.J. Genet, Application of X-ray photoelectron spectroscopy to microorganisms, Colloids Surf., B 2 (1994) 347–369.10.1016/0927-7765(94)80049-9
- L.I. Falcon, S. Pluvinage, E.J. Carpenter, Growth kinetics oellular N2-fixing cyanobacterial isolates in continuous culture in relation to phosphorus and temperature, Mar. Ecol. Prog. Ser. 285 (2005) 3–9.
- H. Seki, H. Ozawa, S. Ichimura, Temperature dependence of filament length of Anabaena spiroides Klebahn var. Crassa Lemm. Hydrobiologia. 83 (1981) 419–423.
- C.D. Collins, C.W. Boylen, Physiological responses of Anabaena variabilis (cyanophyceae) to instantaneous exposure to various combinations of light intensity and temperature, J. Phycol. 2 (1982) 206–211.
- C. van Baalen, D.S. Hoare, E. Barndt, Heterotrophic growth of blue-green algae in dim light, J. Bacteriol. 105 (1971) 685–689.
- D.A. Paone, S.E. Stevens, Nitrogen starvation and the regulation of Glutamine Synthetase in Agmenellum quadruplicatum, Plant. Physiol. 67 (1981) 1097–1100.
- N. Imamura, Studies of the water blooms an Lake Kasumigaura, Verh. Internat. Verein. Limnol. 21 (1981) 652–658.
- R.H. Foy, C.E. Gibson, R.V. Smith, The influence of daylength, light intensity and temperature on the growth rates of planktonic blue-green algae, Br. Physiol. J. 11 (1976) 151–163.
- P. Fay, S.A. Kulasooriya, Tetrazolium reduction and nitrogenise activity in heterocystous blue-green algae, Arch. Mikrobiol. 114 (1973) 701–705.
- W.F. Vincent, W.B. Silvester, Growth of blue-green algae in the Manukau (New Zealand) oxidation ponds. I. Growth potential of oxidation pond water and comparative optima for blue-green algal growth, Water Res. 13 (1979) 711–716.
- W.T. De Nobel, H.C.P. Hatthijs, E. Von Elert, L.R. Mur, Comparison of the light-limited growth of the nitrogen-fixing cyanobacteria Anabaena and Aphanizomenon, New Phytol. 138 (1998) 579–587.
- J.H. Yoon, S.S. Choi, T.H. Park, The cultivation of Anabaena variabilis in a bubble column operating under bubbly and slug flows, Bioresour. Technol. 110 (2012) 430–436.
- J.A. Rother, P. Fay, Sporulation and the development of planktonic blue-green algae in two Salopian meres, Proc. Royal Soc. London B 196 (1977) 317–332.
- U. Uehlinger, Experimental studies on the autoecology of Aphanizomenon flos-aquae, Arch. Hydrobiol. Suppl. 60 (1981) 260–288.
- J.T. Leema, R. Kirubagaran, N.V. Vinithkumar, P.S. Dheenan, S. Karthikayulu, High value pigment production from Arthrospira (Spirulina) platensis cultured in seawater, Bioresour. Technol. 101 (2010) 9221–9227.
- K.J. Reddy, B. Haskell, D.M. Sherman, L.A. Sherman, Unicellular, aerobic Nitrogen-fixing cyanobacteria of the genus Cyanothece, J. Bacteriol. 175 (1993) 1284–1292.
- M. Giordiano, Interactions between C and N metabolism in Dunaliella salina cells cultured at elevated CO2 and high N concentrations, J. Plant. Phys. 158 (2001) 577–581.
- J.J. Ortega-Calvoh, X. Arino, L.J. Stal, C. Saiz-Jimenez, Cyanobacterial Sulfate accumulation from Black Crust of a historic building, Geomicr. J. 12 (1994) 15–22.
- E. Esquenazi, M. Daly, T. Bahrainwala, W.H. Gerwick, P.C. Dorrestein, Ion mobility mass spectrometry enables the efficient detection and identification of halogenated natural products from cyanobacteria with minimal sample preparation, Bioorg. Med. Chem. 15 (2011) 6639–6644.
- M.F. Watanabe, S. Oishi, Effects of environmental factors on toxicity of a cyanobacterium (Microcystis aeruginosa) under culture conditions, Appl. Envir. Micr. 49 (1985) 1342–1344.
- A. Nicklisch, J.G. Kohl, Growth kinetics of Microcystis aeruginosa (Kutz) Kutz as a basis for modelling its population dynamics, Int. Revue Ges. Hydrobiol. 68 (1983) 317–326.
- A.J. van der Westhuizen, J.N. Eloff, Effect of temperature and light on the toxicity and growth of the blue-green alga Microcystis aeruginosa (UV – 006), Planta. 163 (1985) 55–59.
- T.G. downing, C. Meyer, M.M. Gehringer, M. van de Verter, Microcystin content of Microcystis aeruginosa is modulated by nitrogen uptake rate relative to specific growth rate or carbon fixation rate, Environ. Toxicol. 20 (2005) 257–262.
- N.E. Erdmann, E. Zuther, S. Abarzua, Comparative studies on the photoproduction of nonhydrogenous resources by cyanobacteria, Cur. Micr. 25 (1992) 83–87.
- G.H.J Kruger, J.N. Eloff, The effect of temperature on specific growth rate and activation energy of Microcystis and Synechococcus isolates relevant to the onset of natural blooms, J. Limnol. Soc. S. Africa 4 (1978) 9–20.
- M. Takahashi, P. Bienfang, Size structure of phytoplankton biomass and photosynthesis in subtropical Hawaiian waters, Mar. Biol. 76 (1983) 203–2011.
- D.J. Douglas, Microautoradiography-based enumeration of photosynthetic pikoplankton with estimates of carbon-specific growth rates, Mar. Ecol. Prog. 14 (1984) 223–228.
- R. Iturriaga, B.G. Mitchell, Chroococcoid cyanobacteria: A significant component in the food web dynamics of the open ocean, Marine. Ecol. Prog. 28 (1986) 291–297.
- M.R. Landry, L.W. Haas, V.L. Fageness, Dynamics of microbial plankton communities: Experiments in Kaneohe Bay, Hawaii, Mar. Ecol. Prog. Ser. 16 (1984) 127–133.
- A. Musial, M. Plinski, Influence of salinity on the growth of Nodularia spumigena Mertens, Ocean. Hydrobiol. St. 2 (2003) 45–52.
- P.J. Bottomley, C. Van Baalen, Characteristics of heterotrophic growth in the blue-green algae Nostoc sp. strain Mac, J. Gen. Microbiol. 107 (1978) 309–318.
- G.Ahlgren, Growth of Oscillatoria agardhii in chemostat culture. 2. Dependence of growth constants on temperature, Mitt. Int. Verein. Theor. Angew. Limnol. 21 (1978) 88-102.
- R.H. Foy, R.V. Smith, The role of carbohydrate accumulation in the growth of planktonic Oscillatoria species, Br. Phycol. J. 15 (1980) 139–150.
- L. Van Liere, L.R. Mur, Growth kinetics of Oscillatoria agardhii Gomont in continuous culture, limited in its growth by the light energy supply, Microbiol. 115 (1979) 153–160.
- E.R. Zinser, Z.I. Johnson, A. Coe, E. Karaca, D. Veneziano, S.W. Chisholm, Influence of light and temperature on Prochlorococcus ecotype distributions in the Atlantic Ocean, Limnol. Oceanogr. 52 (2007) 2205–2220.
- S. Bertilsson, O. Berglund, D.M. Karl, S.W. Chisholm, Elemental composition of marine Prochlorococcus and Synechococcus: Implications for the ecological stoichiometry of the sea, Limnol. Oceanogr. 48 (2003) 1721–1731.
- L.R. Moore, S.W. Chisholm, Photophysiology of the marine cyanobacterium Prochlorococcus: Ecotypic differences among cultured isolates, Limnol. Oceanogr. 44 (1999) 628–638.
- T. Burger-Wiersma, L.J. Stal, L.R. Mur, Prochlorothrix hollandicagen. Nov., sp. nov., a filamentous oxygenic photoautotrophic prokaryote containing chlorophyls a and b: Assignment to Prochlorotriceae fam. Nov. And order Prochlorales Florenzano, Balloni, and Materassi 1986, with emendation of the ordinal description, Int. J. System. Bacter. 39 (1989) 250–257.
- N.S.R. Agawin, S. Agusti, Abundance, frequency of dividing cells and growth rates of Synechococcus sp. (cyanobacteria) in the stratified Northwest Mediterranean Sea, J. Plankton Res. 19 (1997) 1599–1615.
- I. Tuominen, M. Pollinari, E. Tyystjarvi, T. Tyystjarvi, The SigB sigma factor mediates high-temperature responses in the cyanobacterium Synechocystis sp. strain PCC 6803, FEBS Lett. 580 (2005) 319–323.
- A.K. Singh, T.C. Summerfield, H. Li, L.A. Sherman, The heat shock response in the cyanobacterium Synechocystis sp. strain PCC 6803 and regulation of gene expression by HrcA and SigB, Arch Microbiol. (2006) 273–286.
- N.S.R. Agawin, C.M. Duarte, S. Agusti, Growth and abundance of Synechococcus sp. in a Mediterranean Bay: Seasonality and relationship with temperature, Mar. Ecol. Prog. Ser. 170 (1998) 45–53.
- L. Campbell, E.J. Carpenter, Estimating the grazing pressure of heterotrophic nanoplankton on Synechococcus spp. Using the sea water dilution and selective inhibitor techniques, Mar. Ecol. Prog. Ser. 33 (1986) 121–129.
- B.J. Binder, Y.C. Liu, Growth rate regulation of rRNA content of a marine Synechococcus (Cyanobacterium) strain, Appl. Environ. Microbiol. 61 (1998) 708–717.
- E. Breitbarth, A. Oschlies, J. LaRoche, Physiological constraints on the global distribution of Trichodesmium – Effect of Temperature on diazotrophy, Biogeosciences 4 (2007) 53–61.
- L. Prufert-Bebout, H.W. Paerl, C. Lasson, Growth, nitrogen fixation, and spectral attenuation in cultivated Trichodesmium species, Appl. Environ. Microbiol. 59 (1993) 1367–1375.
