Abstract
This research presents time-geographic density estimation (TGDE) as a new technique of animal home range analysis in geographic information science (GIS). TGDE combines methodologies of time geography and statistical density estimation to produce a continuous probability distribution of an object's spatial position over time. Once TGDE is applied to animal tracking data to create a density surface, home ranges and core areas can be delineated using specified contours of relative intensity (e.g., 95% or 50%). This article explores the use of TGDE for home range analysis using three data sets: a fixed-interval simulated data set and two variable-interval satellite tracks for a loggerhead sea turtle (Caretta caretta) corresponding to internesting and post-migration foraging periods. These applications are used to illustrate the influence of several parameters, including sample size, temporal sampling scheme, selected distance-weighted geoellipse function, and specified maximum velocity, on home range estimates. The results demonstrate how TGDE produces reasonable home range estimates even given irregular tracking data with wide temporal gaps. The advantages of TGDE as compared with traditional methods of home range estimation such as kernel density estimation are as follows: (1) intensities are not assigned to locations where the animal could not have been located given space and time constraints; (2) the density surface represents the actual uncertainty about an animal's spatial position during unsampled time periods; (3) the amount of smoothing applied is objectively specified based on the animal's movement velocity rather than arbitrarily chosen; and (4) uneven sampling intervals are easily accommodated since the density estimates are calculated based on the elapsed time between observed locations. In summary, TGDE is a useful method of home range estimation and shows promise for numerous applications to moving objects in GIS.
1. Introduction
Home range estimation is one of the most widely applied geographic information science (GIS)-based analyses in animal ecology. Home ranges define the spatial area occupied by an individual during its day-to-day activities, excluding any atypical excursions (Burt Citation1943). In practice, the home range is usually interpreted as the smallest area in which an animal spends 95% of its time (Kernohan et al. Citation2001). Core areas, which denote regions of more concentrated activity, are similarly defined as the minimum area enclosing 50% of an animal's movements (White and Garrott Citation1990). Home range analysis involves physically delineating these occupied areas based on sample points that record an animal's spatial position periodically over time. Numerous statistical and polygonal approaches have been proposed to estimate home ranges, including examples such as kernel density estimation (KDE) (Worton Citation1989), minimum convex polygons (Mohr Citation1947), local nearest-neighbor convex-hulls (Getz and Wilmers Citation2004), point peeling algorithms (Bath et al. Citation2006), and characteristic hull polygons (Duckham et al. Citation2008, Downs and Horner Citation2009).
Despite the large number of available techniques, most ecologists have favored KDE – a popular statistical data smoothing technique in GIS (Silverman Citation1986) – for home range estimation. It is often preferred, because it produces a continuous intensity surface that represents an animal's space-use pattern or ‘utilization distribution’ (Worton Citation1989). In this context, KDE is applied to animal tracking data, and then home ranges and core areas are delineated from the 95% and 50% contours of the resulting intensity surface. Despite KDE's status as the most widely applied and recommended home range estimator (e.g., Steury et al. Citation2010), some researchers recently have expressed concern about KDE's accuracy as a home estimator (Hemson et al. Citation2005, Downs and Horner Citation2008, Laver and Kelly Citation2008). For example, some studies have shown that KDE is sensitive to the shape of the point pattern of tracking data (Downs and Horner Citation2008) and tends to overestimate home ranges that are linear (Blundell et al. Citation2001, Row and Blouin-Demers Citation2006), have internal regions of unoccupied space (Getz and Wilmers Citation2004), or are of a patchy nature (Mitchell and Powell Citation2008). Proper selection of the smoothing parameter used with KDE has also been cited as a concern or limitation (e.g., Gitzen et al. Citation2006). However, perhaps most importantly, KDE has been shown to produce biased results when applied to tracking data in general, as KDE assumes points are independent, while tracking data consist of non-independent points which are linked by movement trajectories (Downs Citation2010). As such there has been recent interest in developing and applying trajectory-based approaches such as the Brownian-bridge method (Horne et al. Citation2007) or trajectory-based density estimation using Delaunay triangulation (Downs and Horner Citation2007) to the problem of home range estimation. However, neither of the two techniques has yet to gain popularity.
GIS offers a variety of approaches for analyzing the trajectories of moving objects (Trajcevski et al. Citation2004, Laube et al. Citation2005), although most of these have been untouched by ecologists. Most notable is the field of time geography, which provides a rigorous mathematical framework for documenting and analyzing the movement of objects over time (Miller Citation2005a). Methods of time geography are routinely applied to analyze movements of various mobile agents, such as people and vehicles (Kwan Citation2000, Kwan and Lee Citation2003, Miller Citation2005b, Neutens et al. Citation2007a, Citationb). While time-geographic techniques are equally applicable for animal locational data, apparently they are rarely if ever applied. As such, time geography may offer an innovative framework for analyzing animal movement data. In particular, time-geographic density estimation (TGDE), which generates a continuous probability density surface of an animal's movements, might offer an alternative home range estimator. TGDE produces an intensity surface that is directly analogous to that produced by traditional KDE, except that it is generated based on movement trajectories rather than assuming the data points are independent. Although Downs (Citation2010) suggested the method might be useful for analyzing animal movements, such an application has yet to appear in the literature.
As such, this research adapts TGDE to the problem of home range analysis. This article is outlined as follows. First, a brief overview of fundamental concepts of time geography is provided as necessary background for understanding the TGDE technique. Second, home range analysis is performed on a fixed-interval tracking data set of 600 control points to demonstrate the influence of several input parameters (sample size, distance-weighting function, and maximum velocity) on the results. Then, TGDE is applied to two satellite tracking data sets for a loggerhead sea turtle (Caretta caretta) (corresponding to internesting and post-migration foraging periods) in order to illustrate how the technique performs in practice, as well as to demonstrate its performance for variable-interval tracking data. The results are used to guide a practical discussion of the usefulness of the technique for home range analysis. Additionally, suggestions for future research applications of time-geographic techniques in ecology are provided in Section 4.
2. Materials and methods
2.1. Time-geographic density estimation
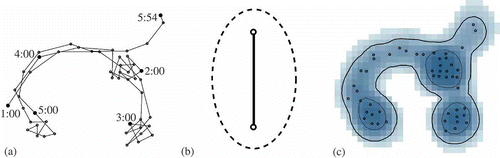
Hägerstrand (Citation1970) first conceptualized time geography as a framework for understanding human behavior and interactions. Miller (Citation2005a) later elaborated on the mathematical foundations for time geography, enabling a rigorous quantitative approach for analyzing the movement and interactions of objects in space and time. The fundamental elements of time geography are as follows: (1) sets of control points, (2) space–time paths, and (3) space–time prisms. A set of control points consists of a collection of observed point locations, sequentially ordered, that represent an object's spatial position over time. The space–time path documents the movement trajectory of the object, which is typically approximated by connecting adjacent control points with straight line segments. These paths can be plotted in three-dimensional space–time or, as illustrated in a, in two-dimensional space. The space–time prism delineates, for any two adjacent control points in a space–time path, the object's space–time volume during the tracking interval. In other words, the prism specifies all spatial locations accessible by the object at each time instance, as determined by constraints imposed by: the spatial locations of the control points; the elapsed time between points; and the object's maximum velocity. Mapped in two-dimensional space, a footprint of the space–time prism yields a geoellipse, which demarcates all spatial locations where the object could have been located during a specified time interval (b). For the specific mathematical formulations of the geometry of these elements, readers are referred to Miller (Citation2005a) and Miller and Bridwell (Citation2009).
Figure 1. A space–time path for a set of control points (a). A geoellipse fit to two consecutive control points. (b) A sample time-geographic density surface with home ranges and core areas delineated using 95% and 50% contours (c).
A recent advance in time geography is the development of a TGDE technique. TGDE, proposed by Downs (Citation2010), incorporates the fundamental elements of time geography with statistical density estimation in order to generate a continuous probability density surface for a moving object over time given a set of observed control points. The technique operates in a manner analogous to KDE except instead of applying a kernel to each data point (Silverman Citation1986), a distance-weighted geoellipse function is fit to each consecutive pair of control points in a space–time path. Mathematically, TGDE can be formulated as
If applied to animal locational data, such as that collected using GPS or satellite telemetry, TGDE would generate a probability distribution of the animal's location during the tracking interval. Assuming the home range is defined as the smallest area containing 95% of an animal's activity, it can be delineated using the 95% isopleth of the TGDE surface. This contouring procedure is identical to how ecologists estimate home ranges using density surfaces generated using traditional KDE (Worton Citation1989). Likewise, core areas of more intense activity can be delineated using the 50% contour. c illustrates 95% and 50% contours for a sample TGDE surface that delineate the home range and core area for a hypothetical animal.
2.2. Parameter selection
TGDE is a newly developed technique in GIS, and the influence of its input parameters has only begun to be explored (Downs Citation2010). The main factors affecting TGDE performance include: (1) the temporal tracking interval and sampling scheme; (2) the selected function of the geoellipse; and (3) specified maximum velocity. We explored the influence of these factors on home range estimates using a sample tracking data set for a hypothetical animal. We used an artificial data set in order to have strict control over the sampling interval while exploring the performance of the technique. The data set generated consisted of 600 control points that were assumed to have been collected using a fixed time interval between control points. The space–time path for the data set is shown in , but we note the path is approximated using straight lines and is not assumed to represent the actual trajectory traveled by the animal between control points. We computed home range and core area estimates for the data set using TGDE under scenarios consisting of three different sample sizes (600, 300, and 200 regularly sampled points), two geoellipse functions (uniform and linear decay), and two hypothetical maximum velocities. For all combinations of factors, home ranges and core areas were delineated using 95% and 50% contours of the resulting TGDE surfaces. The goal of this exercise was to demonstrate the technique and explore the influence of the input parameters on home range estimates before applying it to actual tracking data.
Figure 2. Space–time path for an artificial tracking data set of 600 points.

2.3. Application to sea turtle tracking data
We applied TGDE for home range estimation using ARGOS satellite tracking data in order to demonstrate the technique's performance using variable-interval sampling, as well as to illustrate its utility in practice. We used two tracks for a loggerhead sea turtle that corresponded to internesting and post-migration periods of activity. The turtle (named ‘Flo’) was captured on Casey Key beach (28.7°N, 82.3°W) in Florida, USA, and fit with a platform transmitter terminal (PTT), as previously described elsewhere (Girard et al. Citation2009, Tucker Citation2010). The internesting period, which occurs when a female occupies offshore areas between and immediately after successive nesting attempts (Godley et al. Citation2008), extended from 27 May 2009 to 17 July 2009. The turtle then migrated to its foraging grounds, where it was tracked from 19 August 2009 to 15 February 2010. In both cases, the locations were recorded opportunistically whenever the animal surfaced and communication between the device and satellite was successful. The resulting data sets were filtered to remove suspected erroneous locations using the STAT program (Coyne and Godley Citation2005), where we omitted fixes with location class Z (the lowest accuracy rating) or calculated swim speeds exceeding 4 km/h as per Hays et al. (Citation2001). The final internesting data set included 206 locations with a mean tracking interval of 3.4 h. The maximum gap was 25 h, with 24 gaps exceeding 12 h. The post-migration track was sampled less frequently with an average interval of 8.2 h over 529 locations. The maximum gap was 44.2 h, and 135 intervals surpassed 12 h. We applied TGDE to both data sets using a linear decay function with three maximum velocities for comparison. In all cases, home ranges and core areas were delineated using 95% and 50% contours of the resulting TGDE surfaces.
3. Results
3.1. Simulated data
The application of TGDE to the artificial, fixed-interval tracking data set illustrates the influence of input parameters on the resulting density surfaces, as well as home range and core area estimates derived from their contours. The results of each scenario are mapped in First, the exploratory analysis demonstrates the influence of the tracking interval (i.e., sample size) on the TGDE output. More precise estimates of home ranges are possible using more frequent tracking, as smaller potential path areas are generated given increased information about the animal's spatial position over time. For wider tracking intervals, there is greater uncertainty about the animal's location between control points, so larger potential path areas, and consequently home ranges and core areas, are produced. This suggests that TGDE results will always improve by increasing the temporal frequency of tracking data. If too wide of an interval is used in the analysis, the amount of uncertainty may be too great to precisely delineate home ranges and core areas.
Figure 3. Application of time-geographic density estimation to an artificial tracking data set using multiple sample sizes, geoellipse functions, and maximum velocity (v) parameters.

Second, the choice of a geoellipse function also influences the results of the time-geographic technique, although this is only to a minor degree compared with the tracking interval, given this particular sample data set. As illustrated in , the chosen function controls how the density is distributed within the computed potential path area. Since the uniform function distributes the density equally within each geoellipse, it produces more even density surfaces than the linear decay function which assigns higher densities nearer approximated space–time paths. Consequently, the uniform function produces slightly larger home range and core area estimates than for the linear decay scenarios. As such, the relative accuracy of the two functions in practice would depend on how well the approximated space–time path represents the actual movement trajectory of the animal; when the path is more accurate, then a linear decay function would be more applicable.
Additionally, TGDE results for the simulated data illustrate the influence of the choice of the maximum velocity parameter. Downs (Citation2010) suggested that a theoretical or observed maximum velocity should be specified. Here, we applied to hypothetical values that were reasonable given that all control points fell within the computed potential path areas. The scenarios in highlight how the animal's estimated home ranges and core areas are considerably larger when a higher velocity is specified. This is theoretically always the case, since higher velocities will produce larger geoellipses and consequently larger potential path areas which lead to larger home range estimates. These results indicate that the maximum velocity parameter plays an important role in home range estimation by TGDE, especially since this value is specified by the user rather than being an artefact to how the data were collected.
3.2. Sea turtle data
The application of the TGDE to sea turtle data documents how the method performs for irregularly sampled control points at different intervals. a illustrates the approximated space–time path and variable tracking interval for the internesting period, while b–d report the results for each specified velocity of 1.5, 1.0, and 0.75 km/h, respectively. reports similar results for the post-migration track with velocities of 2.0, 1.5, and 1.0 km/h. For both periods, note how the resulting density surfaces and delineated contours compensate for the uneven tracking intervals. The home range contours are more precisely drawn around locations recorded in more frequent succession, while they extend wider in areas with large temporal gaps in sampling since there is greater uncertainty about where the animal occurred during unsampled times. This feature is most evident in the northern part of the turtle's internesting range, where few locations were recorded but they represented relatively large amounts of time. As such, the animal's location during unsampled time periods is more precisely drawn during times of frequent tracking and more approximated using large geoellipses during infrequently sampled periods.
Figure 4. Space–time path (a) and time-geographic home range estimates (b-d) using three maximum velocity (v) parameters for a loggerhead sea turtle during the internesting period.
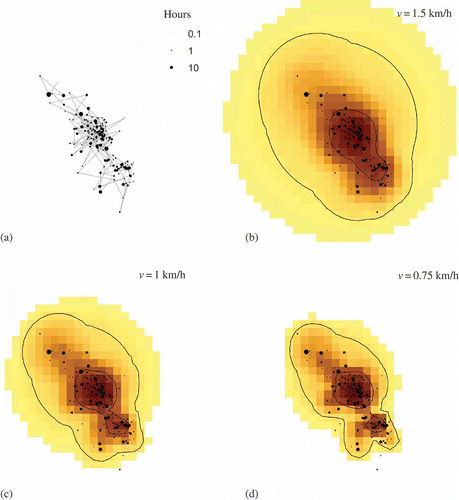
Figure 5. Space–time path (a) and time-geographic home range estimates (b-d) using three maximum velocity (v) parameters for a loggerhead sea turtle during the post-migration foraging period.
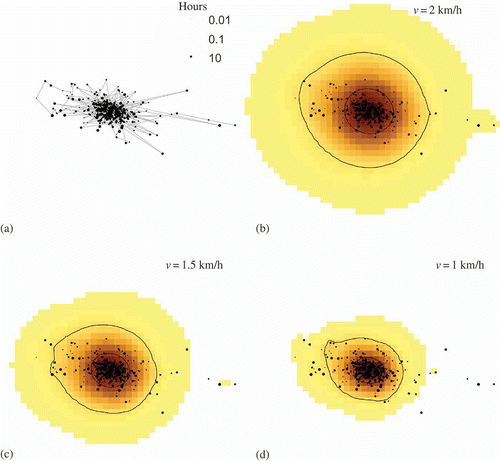
Additionally, the results for the loggerhead sea turtle further emphasize the influence of the maximum velocity parameter on time-geographic home range estimates. As for the artificial fixed-interval data, larger velocities produced larger home range and core area estimates. For the data sets used in this study, it is impossible to determine which velocity produced the most accurate results since the true home range of the animals remains unknown. However, for the actual tracking data sets, it is possible to judge the relative merits of each choice of velocity; the smallest estimate is not necessarily the most accurate even if it provides the ‘best fit’ to the points by visual inspection. A simple diagnostic is the number of locations that occur outside the computed potential path area. For the internesting period, all points fell within the potential path area for the 1.5 and 1.0 km/h velocities, and two points occurred outside for the lowest velocity. For the post-migration period, all points were located in the potential path area at 2 km/h, only two occurred outside at 1.5 km/h, and a large number fell outside for 1 km/h. Points occur outside the potential path area when the observed distance between consecutive control points exceeds the maximum possible given the specified velocity, in which case no intensity is computed for that geoellipse. For the points located outside the potential path area, either the turtle was traveling faster than the specified velocity or that the spatial position of the control point was incorrectly recorded. If the data are filtered for accuracy beforehand, then the occurrence of multiple points falling outside this area is a clear indication that the velocity parameter was set too low since the animal must have traveled at higher speeds than specified.
For the internesting period, 1 km/h appears to produce the most reasonable home range and core area estimates; one point occurs at the edge of the computed potential path area, and all but two points occur within the delineated home range. Since the home range contains 95% of activity, not all points should necessarily be included within its boundary.However, for the post-migration period, a velocity parameter of 1 km/h is too small, since many points occur outside the potential path area. These outlying points – as well as others within the range – are not included in the density computations since they were located too far from their neighbors in the space–time path based on the specified velocity; consequently numerous points are excluded from the delineated home range, further indicating the parameter is set too low. The higher velocities both seem to produce reasonable home range estimates since all but a few points occur within the potential path area. Since speeds of ∼2 km/h were computed for about 25% of the locations in the post-migration data set, that velocity may produce the most reasonable estimate even though the home range boundary appears more generalized than for the 1.5 km/h velocity. Additionally, a large number of points fall outside the computed home range for the 1.5 km/h velocity, also indicating the estimated area is possibly too small. This evidence suggests that a velocity parameter of 2 km/h produced the most reasonable home range estimates for the post-migration period given those explored. However, the results indicate the time-geographic technique still produced a reasonable post-migration home range estimate given a tracking interval with very wide gaps in sampling compared with the internesting period which was delineated more precisely.
4. Discussion and conclusions
This research presents TGDE as a novel methodology for estimating animal home ranges from GPS and satellite tracking data. The advantages of TGDE for home range analysis compared with traditional KDE are that (1) intensities are not assigned to locations where the animal could not have been located; (2) the density surface represents the actual uncertainty about an animal's spatial position during unsampled time periods; (3) the amount of smoothing applied is objectively specified based on the animal's movement velocity rather than arbitrarily chosen; and (4) uneven sampling intervals are easily accommodated since the size of each geoellipse function is derived from space–time constraints imposed by the control points. The exploratory analysis performed using the fixed-interval simulated and variable-interval sea turtle tracking data indicate the time-geographic technique can produce meaningful home range estimates under a range of scenarios. While somewhat limited in scope, these applications do illustrate how the TGDE results are sensitive to a number of factors, including the tracking interval and sampling scheme, geoellipse function, and specified maximum velocity. There are several implications of this research.
First, the results of both the artificial and the sea turtle data sets document how the precision of home ranges estimated by TGDE necessarily improve as the temporal frequency of sampling is increased. When the tracking interval is rapid, precise home ranges can be delineated if there is a large enough sample of points. However, for very infrequent intervals (e.g., one location per week), TGDE will generate surfaces with too much uncertainty to accurately delineate home ranges, and alternative estimators such as those that use neighborhood graphs to estimate trajectories (Downs and Horner Citation2007) might be necessary. Although GPS tracking technologies permit frequent tracking, a concern is that the current trend in ecology has been to sample less frequently over longer periods of time. This is partially done to extend the battery life of the tracking device but often also to reduce autocorrelation in the data set, which is seen as problematic for some home range estimators such as KDE (Katajisto and Moilanen Citation2006, Fieberg Citation2007). Although TGDE is robust to autocorrelation, extending battery life and collecting long-term data are desirable in many cases. Consequently, an optimal strategy might be to use the widest tracking interval that still produces accurate home range estimates in terms of size and shape. Future work might explore this trade-off for TGDE using detailed animal tracking data collected in the field for which known home ranges can be computed.
Second, also with regard to data collection, the overall sampling scheme is an important consideration. TGDE explicitly handles irregular tracking intervals, because it computes the size of each geoellipse and assigns the corresponding density in proportion to the specified time interval. However, systematic fixed-interval tracking can produce improved results over irregular intervals. This is because home ranges and core areas delineated from variable-interval data can display varying degrees of precision throughout the animal's range if the variance in the tracking interval is unsystematic. This was the case for our sample of sea turtle tracking data for the internesting period, where contours were less precise in the northern part of the animal's range. Although regular intervals are more desirable, irregular tracking is common especially in the marine environment, where locations can only be recorded when the animal surfaces for enough time to fix its location. As such, the TGDE technique may offer the most suitable method for analyzing tracking data sets with unsystematic and irregular sampling intervals, as it realistically factors in the amount of time elapsed between control points when generating home range estimates.
The exploratory analysis also highlights the influence of the choice of geoellipse function on home range estimates. Here, we only considered two possible functions – uniform and linear decay. For our artificial example, both functions produced similar results. However, other distance-weighting functions might produce contrasting results to those observed in this study. For example, a quartic function (Silverman Citation1986) would distribute even higher densities nearer to estimated space–time paths than the linear decay function and might offer improved results for high temporal frequency tracking data. Future work might further evaluate how these and other distance-weighting functions influence home range estimates for a broader array of scenarios or tracking data sets.
Finally, application of the time-geographic technique to all three data sets reveals the large influence of the maximum velocity parameter, which controls the size of the computed geoellipses and accordingly the amount of smoothing. Larger values always produce larger potential path areas and home range estimates. Downs (Citation2010) suggested using either the theoretical or the maximum velocity when applying the technique. A useful procedure for evaluating the appropriateness of the chosen value is to confirm all control points are located within the computed potential path area and that most points fall within the delineated home range; if not, the chosen value is likely too small. Choosing a parameter that is too small excludes too many points from contributing to the density surface and can lead to an underestimation of home range sizes. Selecting a velocity too large can have the opposite effect, generating home ranges that are too large. These findings suggest that care should be exercised in selecting this parameter when applying the technique in practice.
In summary, this research presents TGDE as a new method for estimating animal home ranges. While the technique may provide researchers with an improved approach for analyzing animal locational data, the implications of this research extend more broadly beyond home range estimation. Time geography offers a general framework for analyzing the behavior and interactions of humans in GIS, and these techniques are equally applicable for animals. Adopting the framework of time geography in ecology might enhance our ability to understand the behavior and habitat needs of species, which can in turn aid conservation and management efforts. Future research might explore how time-geographic and other trajectory-based techniques can be used to analyze animal movement and space-use patterns.
Furthermore, the results of this research – particularly in terms of parameter and temporal sampling scheme selection – can be used to guide the application of TGDE to tracking data for other types of mobile objects in GIS. Such objects of interest to GIS scientists include pedestrians, ships, storms, and vehicles, and TGDE might provide an innovative way to explore the movements of these types of agents. In the context of people, TGDE might be used to analyze cumulative activity patterns for groups (Casas et al. Citation2009), and the technique might be extended to visualize collections of trajectories in three dimensions in a manner similar to the traditional kernel approach employed by Demšar and Virrantaus (Citation2010). In the case of vehicles, which do not travel in continuous Euclidean space since they are restricted to roadways, an adaptation of TGDE to the network data model would be warranted. Future work should explore adaptations of TGDE to networks, three-dimensional geovisualization, and other analytic situations.
Acknowledgments
Portions of this research were funded by grants made to the authors from the National Science Foundation (BCS-1062947 (Downs); BCS-1062924 (Horner)). The contents of this article are the responsibility of the authors and do not reflect the views of the NSF. The loggerhead sea turtle tracking data were a subset collected through projects funded with major support from the Sea Turtle Grants Program (# 07-024R, #08-019R, #09-001R (Tucker)) and other support provided by Disney Worldwide Conservation Fund, MOTE Scientific Foundation, Norcross Wildlife Foundation, New Canaan County School/Jeniam Foundation, Wooster School, the Samek family, Morrison Family Foundation, V. Miller, Sarasota County Environmental Services, NASA Signals of Spring, Longboat Key Turtle Watch, Coastal Wildlife Club, Curtis School, Community Foundation of Greater Lakeland, Comerica, Suntrust Bank, Seaturtle.org, and West Marine of Sarasota. Animal handling was in accordance with IACUC permit 07-04-AT1 and Marine Turtle Permit #126 from the Florida Fish and Wildlife Conservation Commission.
Related Research Data
References
- Bath , S.K. 2006 . Characterization of home range using point peeling algorithms . Journal of Wildlife Management , 70 ( 2 ) : 422 – 434 .
- Blundell , G.M. 2001 . Linear home ranges: effects of smoothing, sample size. and auto correlation on kernel estimates . Ecological Monographs , 71 ( 3 ) : 469 – 489 .
- Burt , W.H. 1943 . Territoriality and home range as applied to mammals . Journal of Mammalogy , 24 : 346 – 352 .
- Casas , I. , Horner , M.W. and Weber , J. 2009 . A comparison of three methods for identifying transport-based exclusion: a case study of children's access to urban opportunities in Erie and Niagara counties . International Journal of Sustainable Transportation , 3 ( 4 ) : 227 – 245 .
- Coyne , M.S. and Godley , B.J. 2005 . Satellite tracking and analysis tool (STAT): an integrated system for archiving, analyzing and mapping animal tracking data . Marine Ecology Progress Services , 301 : 1 – 7 .
- Demšar , U. and Virrantaus , K. 2010 . Space–time density of trajectories: exploring spatio-temporal patterns in movement data . International Journal of Geographical Information Science , 24 ( 10 ) : 1527 – 1542 .
- Downs , J.A. 2010 . Time-geographic density estimation for moving point objects . Lecture Notes in Computer Science , 6292 : 16 – 26 .
- Downs , J.A. and Horner , M.W. Network-based home range analysis using Delaunay triangulation . The 4th international symposium on voronoi diagrams in science and engineering . July 9–12 2007 . Edited by: Gold , C.M. pp. 255 – 259 . Pontypridd, Wales, , UK : IEEE . University of Glamorgan. doi: 10.1109/ISVD.2007.31
- Downs , J.A. and Horner , M.W. 2008 . Effects of point pattern shape on home-range estimates . Journal of Wildlife Management , 72 ( 8 ) : 1813 – 1818 .
- Downs , J.A. and Horner , M.W. 2009 . A characteristic-hull based method for home range estimation . Transactions in GIS , 13 : 527 – 537 .
- Duckham , M. 2008 . Efficient generation of simple polygons for characterizing the shape of a set of points in the plane . Pattern Recognition , 41 : 3224 – 3236 .
- Fieberg , J. 2007 . Kernel density estimators of home range: smoothing and the autocorrelation red herring . Ecology , 88 ( 4 ) : 1059 – 1066 .
- Getz , W.M. and Wilmers , C.C. 2004 . A local nearest-neighbor convex-hull construction of home ranges and utilization distributions . Ecography , 27 ( 4 ) : 489 – 505 .
- Girard , C. , Tucker , A.D. and Calmettes , B. 2009 . Post-nesting migrations of loggerhead sea turtles in the Gulf of Mexico: dispersal in highly dynamic conditions . Marine Biology , 156 ( 9 ) : 1827 – 1839 .
- Gitzen , R.A. , Millspaugh , J.J. and Kernohan , B.J. 2006 . Bandwidth selection for fixed-kernel analysis of animal utilization distributions . Journal of Wildlife Management , 70 : 1334 – 1344 .
- Godley , B.J. 2008 . Satellite tracking of sea turtles: Where have we been and where do we go next? . Endangered Species Research , 4 ( 1–2 ) : 3 – 22 .
- Hägerstrand , T. 1970 . What about people in regional science? . Papers of the Regional Science Association , 24 : 7 – 21 .
- Hays , G.C. 2001 . The implications of location accuracy for the interpretation of satellite-tracking data . Animal Behaviour , 61 : 1035 – 1040 .
- Hemson , G. 2005 . Are kernels the mustard? Data from global positioning system (GPS) collars suggests problems for kernel home-range analyses with least-squares cross-validation . Journal of Animal Ecology , 74 ( 3 ) : 455 – 463 .
- Horne , J.S. 2007 . Analyzing animal movements using Brownian bridges . Ecology , 88 ( 9 ) : 2354 – 2363 .
- Katajisto , J. and Moilanen , A. 2006 . Kernel-based home range method for data with irregular sampling intervals . Ecological Modelling , 194 ( 4 ) : 405 – 413 .
- Kernohan , B.J. , Gitzen , R.A. and Millspaugh , J.J. 2001 . “ Analysis of animal space use and movements ” . In Radio tracking animals and populations , Edited by: Millspaugh , J.J. and Marzluff , J.M. 125 – 166 . San Diego, CA : Academic Press .
- Kwan , M.P. 2000 . Interactive geovisualization of activity-travel patterns using three-dimensional geographical information systems: a methodological exploration with a large data set . Transportation Research Part C – Emerging Technologies , 8 ( 1–6 ) : 185 – 203 .
- Kwan , M.P. and Lee , J. 2003 . “ Geovisualization of human activity patterns using 3D GIS: a time-geographic approach ” . In Spatially integrated social science: examples in best practice, chapter 3 , Edited by: Goodchild , M.F. and Janelle , D.G. 48 – 56 . Oxford University Press .
- Laube , P. , Imfeld , S. and Weibel , R. 2005 . Discovering relative motion patterns in groups of moving point objects . International Journal of Geographical Information Science , 19 ( 6 ) : 639 – 668 .
- Laver , P.N. and Kelly , M.J. 2008 . A critical review of home range studies . Journal of Wildlife Management , 72 ( 1 ) : 290 – 298 .
- Miller , H.J. 2005a . A measurement theory for time geography . Geographical Analysis , 37 ( 1 ) : 17 – 45 .
- Miller , H.J. 2005b . Necessary space-time conditions for human interaction . Environment and Planning B: Planning & Design , 32 ( 3 ) : 381 – 401 .
- Miller , H.J. and Bridwell , S.A. 2009 . A field-based theory for time geography . Annals of the Association of American Geographers , 99 ( 1 ) : 49 – 75 .
- Mitchell , M.S. and Powell , R.A. 2008 . Estimated home ranges can misrepresent habitat relationships on patchy landscapes . Ecological Modelling , 216 ( 3–4 ) : 409 – 414 .
- Mohr , C.O. 1947 . Table of equivalent populations of North American small mammals . American Midland Naturalist , 37 : 223 – 249 .
- Neutens , T. 2007a . Human interaction spaces under uncertainty . Transportation Research Record , 2021 : 28 – 35 .
- Neutens , T. 2007b . Space-time opportunities for multiple agents: a constraint-based approach . International Journal of Geographical Information Science , 21 ( 10 ) : 1061 – 1076 .
- Row , J.R. and Blouin-Demers , G. 2006 . Kernels are not accurate estimates of home-range size for herpetofauna . Copeia , 4 : 797 – 802 .
- Silverman , B.W. 1986 . Density estimation for statistics and data analysis , London : Chapman and Hall .
- Steury , T.D. 2010 . Evaluation of root-n bandwidth selectors for kernel density estimation . Journal of Wildlife Management , 74 : 539 – 548 .
- Trajcevski , G. 2004 . Managing uncertainty in moving objects databases . ACM Transactions on Database Systems , 29 ( 3 ) : 463 – 507 .
- Tucker , A.D. 2010 . Nest site fidelity and clutch frequency of loggerhead turtles are better elucidated by satellite telemetry than by nocturnal tagging efforts: implications for stock estimation . Journal of Experimental Marine Biology and Ecology , 383 ( 1 ) : 48 – 55 .
- White , G.C. and Garrott , R.A. 1990 . Analysis of wildlife radio-tracking data , San Diego, CA : Academic Press .
- Worton , B.J. 1989 . Kernel methods for estimating the utilization distribution in home-range studies . Ecology , 70 ( 1 ) : 164 – 168 .