Abstract
Based on a selection of 230 papers published during the last 15 years in international journals, the present work aims at evaluating the state of the art of limnological research in the deep southern subalpine lakes (DSL: Garda, Iseo, Como, Lugano and Maggiore). Historically, most of the limnological research was fostered by the need to find solutions to the problems connected with eutrophication and pollution. Many data are available on the thermal structure, algal nutrient concentrations and phytoplankton of the DSL, while other topics still remain more or less constrained to single lakes. Apart from this geographical bias, a number of aspects emerged from this synoptic view. Limnological research is still linked to the concept of scientific monitoring, while experimental studies and modelling are confined to specific niches; the integration of different disciplines is held back by the division of studies on different compartments; integration of studies and synoptic analyses at a macro regional scale have been carried out only for specific research areas. The DSL are increasingly threatened by new pressures (climatic change, excessive proliferation of toxic cyanobacteria, introduction of new species and new micropollutants) and by the interactions among these new and old stressors. In this rapidly changing situation, the paper emphasises the need to define criteria to be used to distinguish research able to produce relevant results and predictive models, which are essential elements for an efficient management of water resources.
1. Introduction
The geographical distribution of lakes in Italy is strongly biased in favour of the Alpine and subalpine regions. The number of lakes characterised by a surface greater than 0.2 km2 is over 500, with an overall volume of water of more than 150×109 m3. Of these, around 80% of the water is concentrated in five lakes located along the southern border of the Alpine chain, namely the lakes Garda, Iseo, Como, Lugano and Maggiore (deep southern subalpine lakes, DSL; ; ). In the catchment areas of these lakes, water is extensively used for the production of hydroelectric power, whereas south of the DSL, in the plain of the River Po, water is intensively used in agriculture and industry, becoming a life-sustaining element for the economy of the most densely populated and productive area in Italy. In addition, these water bodies are one of the key elements for the tourist economy of the Alpine region. Since limnological studies began, the DSL have attracted a large number of scientists, fostering a large number of investigations in different fields (see [1–230] and Citation231).
Figure 1. Geographical location of the deep southern subalpine lakes (DSL) in northern Italy.

Table 1. Morphometric and hydrological characteristics of the deep southern subalpine lakes (DSL) in Northern Italy.
Table 2. Principal limnological studies carried out since 1995 in the deep southern subalpine lakes (DSL). Studies carried out on two or more lakes are reported under ‘DSL’.
The largest lakes south of the Alps share many common characteristics Citation32,Citation232. They are narrow, elongated and delimited laterally by steep sides. Their bottom is generally flat and the thalwegs are roughly north-south oriented, with the exception of the north basin of Lake Lugano, which is oriented east-west. Lake Garda is the largest Italian lake, followed by Lake Maggiore, Lake Como and Lake Iseo (). Lake Lugano stands out for its subdivision into three basins Citation233. The northern basin is the largest of the three, accounting for around 80% of the total water volume. The maximum depths of the DSL range from 251 m (Lake Iseo) to 410 m (Lake Como). The DSL contribute about 40% to the discharge of the River Po, the largest Italian river. Their theoretical renewal times are around 4–5 years (lakes Maggiore, Como and Iseo), 12 years (Lake Lugano, N-basin) and 27 years (Lake Garda). Compared with the other deep subalpine lakes, Lake Garda has a longer theoretical water renewal time (27 years), due to its low catchments area to lake volume ratio and lower annual rainfall.
Large and deep lakes present peculiar ecological characteristics, which justify their inclusion in a separate typology Citation234. The size and form of lakes regulate many transport processes–such as sedimentation and re-suspension, diffusion, mixing, and the burial of organic and inorganic matter–which regulate many abiotic factors, including phosphorus availability, water chemistry and water clarity Citation235. In turn, variations in these fundamental ecosystem characteristics strongly regulate primary and secondary production Citation236. The formation of a strong thermocline during the warmest period contributes to isolating large portions of the upper pelagic zone for many months, suppressing vertical mixing and inhibiting the transport of nutrients towards the trophogenic layers. In contrast to the shallower lakes of the temperate latitudes, which are strictly dimictic or monomictic Citation237, complete vertical mixing of deep lakes can be prevented by warmer winter temperatures or low wind conditions. Therefore the deep hypolimnion may be characterised by several years of gradual warming caused by a downward transport of heat by turbulent diffusion during stratification, terminated by sudden cooling during harsh winters Citation238, with strong implications for the spring vertical replenishment of nutrients from the deeper layers and for primary production Citation15. In this regard, the deep lakes south of the Alps could be classified as warm monomictic lakes. In this category, lakes circulate completely once a year in the winter at or above 4°C, showing stable stratification for the remainder of the year. Nevertheless, owing to their great depth, complete mixing can occur only during cold winters with complete cooling of the water column. This condition–known as oligomixis–is different from that of lakes which circulate freely every year (holomictic lakes) or show a constant stratification (meromictic lakes).
From a biogeochemical perspective, nutrient cycling in large lakes becomes more and more important compared with inputs from the watershed or interactions with the littoral or lake bottom, even though important differences exist due to hydrology and watershed size in relation to lake volume. An important pathway by which nutrients are recycled in the pelagic zone is through the microbial loop, whereas severe limitation of epilimnetic nutrients in summer can be detected in less enriched environments due to settling fluxes of plankton Citation239.
From a synecological perspective, large water bodies tend to operate as large inertial systems, minimizing the effects of external disturbances. Chronological sequences of phytoplankton assemblages generally show ordered cyclical patterns, in contrast with the unpredictable and disorderly development observed in smaller lakes, which are more susceptible to meteorological and hydrological events Citation84. In turn, these properties allow a major degree of predictability and definition of management tools, e.g. Citation103,Citation104.
In large and deep lakes, the contribution of the fauna and flora of the shores in shaping plankton composition and biotic interactions may become less important, even if only apparently for a few species (e.g. see section 3.7). Nevertheless, the naturalness of riparian zones is essential for the proper functionality, sustainability and recreational attractiveness of lake shores. Moreover, from a biodiversity perspective, many large lakes are known for hosting a large number of endemic species; see, e.g. Citation240,Citation241. A part from well known cases (e.g. Salmo trutta carpio in Lake Garda Citation242), the biocenosis of the DSL present many common traits. Many changes due to anthropic pressure occurred, however, during the last decades.
The natural functionality of trophic webs in the DSL underwent alterations due to the introduction of allochtonous species, both in historical time–e.g. fishes, macrophytes, molluscs Citation242–and more recently, e.g. with the introduction of new zooplankton, shrimps, molluscs and fishes (section 3). On the other hand, the principal alterations of the pristine ecological conditions were due to significant changes in the algal nutrient loads, and to the progression of eutrophication since the 1970s. Climate change, occurring on top of all other anthropogenic pressures, is considered a worsening factor for the ecological status of lake ecosystems Citation234, but the quantification of its exact role is still problematic due to the exceeding importance of nutrients compared with the slow changes occurring in the physical properties of the water columns. A further element of degradation was due to the discharge and deposition of toxic contaminants. A substantial pollution from specific contaminants is still present in a few lakes due to historical pollution (e.g. DDT in Lake Maggiore) or to release of paleo-contaminants from the melting of glaciers (e.g. lakes Como and Iseo, DDT).
Trying to go through these topics, this review has two specific objectives: (i) to synthesize the whole set of information published on the limnology of the DSL in peer-reviewed journals since 1995, in order to identify the more recent important topics and research directions in the last 15 years and (ii) to highlight gaps in the research topics, identifying future research directions in relation to the present and future multi-stressor scenarios.
The review of limnological literature was based on a search made on international journals from 1995 to January 2010. Besides consulting the ISI Web of Science (ISI-WoS, Thomson Scientific's Institute for Scientific Information), other limnology serials were consulted, including Memorie dell’Istituto Italiano di Idrobiologia (1995–1998) and its successor, the Journal of Limnology, volumes 58(1)–69(1); Documenta Istituto Italiano di Idrobiologia (1995–1997); Proceedings of the International Society of Limnology (Verhandlungen der internationalen Vereinigung für theoretische und angewandte Limnologie), volumes 26(1)–30(9). Other papers with impact factor not yet included in ISI-WoS but with DOI (Digital Object Identifier) were considered whenever possible. The use of international sources constrains the selection to peer reviewed papers, allowing easy accessibility to the literature by interested readers. However, besides these sources, a lot of information is also included in grey literature, which is often difficult to obtain. Bibliographical information about papers published in local journals or in many national proceedings have been partly summarised in Tartari et al. Citation243, www.limno.eu and www.iii.to.cnr.it. The papers published on the limnology of the DSL were analysed by Correspondence Analysis Citation244, taking into consideration the mutual relationships between lakes and topics (see section 3). Before CA computation, the data were subjected to a square root transformation (Yi = ) to reduce the weight of the topics with the highest number of papers.
Besides a critical description based on published literature, this work will also provide an update of the long-term series of temperature, TP and hypolimnetic dissolved oxygen recorded until 2008. Since serial correlation in the climate time series can influence the estimate of trend, besides the usual Mann–Kendall (MK) test Citation245, the significance of the temperature trends were computed utilizing the trend-free pre-whitening (TFPW) procedure proposed by Yue et al. Citation246. Moreover, the significance of the MK analyses was confirmed with the application of bootstrap tests for trend detection Citation245. The degree of temperature change (slopes) was estimated with the non-parametric Theil-Sen Approach Citation245. Statistical analyses were carried out with the R package Citation247.
2. A brief overview of the studies carried out in the DSL in the 1900s
Deep lakes were considered of primary interest from the earliest limnological studies, in spite of the practical difficulties deriving from their dimensions. The model of scientific study of lakes remained for decades the fundamental work of F.A. Forel and of the staff of scientists he coordinated, resulting in the impressive monograph on Lake Geneva Citation248. Following these activities, Pietro Pavesi started his observations on Lake Maggiore, looking for the pelagic forms of life highlighted in Lake Geneva. In a synthesis of his work, Pavesi Citation249 compared the numerous species he found with those observed in Lake Geneva, and discussed the presence of plankton in lakes in the light of the theory accepted at the time. Two of Pavesi's collaborators developed the hydrobiological studies. Marco De Marchi, from the laboratory he installed in his villa in Pallanza, on the shore of Lake Maggiore, continued with greater regularity the sampling of plankton species, trying to individuate the relationships among the different organisms and the main physical variables (light, water temperature) influencing their distribution in the water column Citation250. More regular and organised was the activity of Rina Monti, who progressively considered waters of subalpine and high altitude lakes. She organised a group of specialists in different types of aquatic organisms, carrying out regular sampling of plankton in many of the lakes of northern Italy. Among her most important results is a monograph on Lake Como Citation251 and the first synoptic work on the DSL Citation252.
With the foundation of the Istituto Italiano di Idrobiologia, studies on the DSL were enhanced, with special attention being paid to Lake Maggiore. Following Monti's tradition, Edgardo Baldi produced a second important synoptic paper on the DSL Citation253. Between the 1950s and the 1970s, further scholars of Monti's, Livia Pirocchi Tonolli and Vittorio Tonolli, coordinated activities dealing with different physical, chemical and biological aspects of Lake Maggiore (for a review see Citation231). Special attention was directed towards the mixing dynamics of the lake and the effects on the biology, the progressive deterioration of lake water because of eutrophication, and the long-term variation of biota as an effect of the modified trophic conditions Citation76,Citation232,Citation254. Richard Vollenweider Citation255,Citation256 made important comparisons between lakes Maggiore, Lugano and Como, considering in detail the mixing processes and their effects on water-column chemistry, with special attention given to the redistribution of nutrients from deep to productive waters.
A comparative study of the deep southern alpine lakes was also made from 1973 to 1982 within the framework of the C.N.R. National Project for the Improvement of Environmental Quality Citation257. This study started a regular monitoring of the chemistry of the DSL, performed in spring, mainly aimed to follow the trophic evolution and the extent of the vertical mixing of waters. Furthermore, the growing interest in eutrophication led to the setting up of a scientific committee within the International Commission for the protection of Italian Swiss Common Waters (CIPAIS), with the specific aim of controlling the eutrophication of lakes Maggiore and Lugano. The first results highlighted the need for homogeneous sampling and analysis techniques, as well as long-term observations to obtain comparable results and reliable information on the temporal trophic development.
Since the 1980s and 1990s, further studies were promoted in the whole group of the DSL by a larger number of research institutions. The collaboration between scientists operating in the different limnological centres has been organised in an informal working group on the limnology of the DSL, which synthesised the chemical and biological analyses recorded in the different institutes and promoted a harmonization of field and laboratory methods, improving the comparability of results Citation258. The studies were also aimed at evaluating the effects of the remedial measures adopted in the 1980s and 1990s on the different lakes, which were intended to reduce the impact of the discharge of pollutants, with special attention to phosphorus, on biotic communities Citation259. Besides these basic limnological studies, further investigations focused on other specialised topics related to urgent environmental concerns (e.g. ecotoxicology) and other emerging environmental and scientific issues (e.g. global warming, microbial ecology).
3. Limnological research at the turn of the twenty-first century
A total of 230 papers were obtained from the exploration of the available literature published in international peer reviewed journals (). A selection of these papers will be commented in the sections 3.1–3.12. The references have been listed for studies carried out in individual lakes or in groups of lakes. Further, the papers were classified into different topics. In a few circumscribed cases, such an attempt met difficulties due to the multidisciplinary nature of investigations, so specific choices may not be considered satisfactory for everybody. Nevertheless, even in these few cases, the choices were based on the prevalent activities, so the proposed classification can be considered representative of the current status of the research in the DSL. The archive obtained this way can be analysed with different bibliometric approaches. However, in this context, we will limit our goal to an analysis of the scientific topics.
Just a brief glance at allows differences in the quantity of works published in the different lakes to be recognised. The largest number of papers reported studies on the limnology of Lake Maggiore (100 contributions), followed by studies concerning lakes Garda and Lugano, as well as the entire group or different group of lakes (DSL, around 30 papers); lakes Como and Iseo had the least number of contributions. Further differences emerge when the analyses take into consideration the main topics covered in the different studies. The results of the Correspondence Analysis (CA) based on clearly indicated the existence of clusters of topics strongly associated with single lakes, or groups of lakes (). The CA did not include topics with less than three cases (MorGeo). The eigenvalues associated with the first two CA axes were 0.23 and 0.19, with explained proportions of 35 and 30%, respectively. The most striking differences in include the association of the studies on microbial ecology, zooplankton, paleolimnology, fishes and ecotoxicology with Lake Maggiore; geochemistry with Lake Lugano; birds with lakes Como and Lugano; and remote sensing with lakes Iseo and Garda. The remaining topics (physical limnology, hydrology, eutrophication, phytoplankton, benthos, aquatic parasites) contributed less to the observed differences among lakes. A striking element which emerges from the analysis is the absence of international studies on the littoral macro- and micro-vegetation (macrophytes and periphytic communities).
Figure 2. Ordination of the DSL and research topics in the correspondence analysis plane defined by the first two axes. Research topics have been coded as in . Gar, Garda; Ise, Iseo; Com, Como; Lug, Lugano, Mag, Maggiore; DSL, deep southern subalpine lakes.
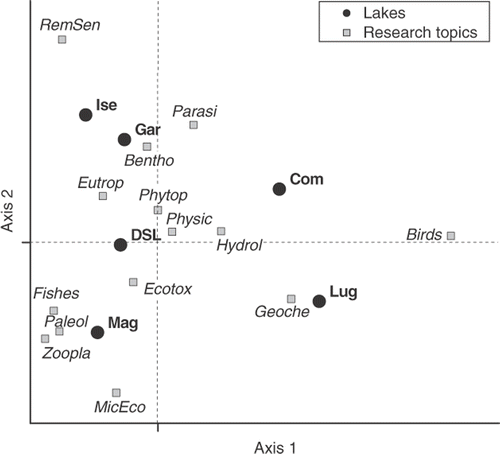
A few differences emerged from a comparison of the works published in the first and second half of the considered period (1995–2002 and 2003–2010). The most significant changes included a decrease of papers dealing with eutrophication, microbial ecology, zooplankton and fish, and an increase of studies regarding phytoplankton and ecotoxicology.
3.1. Physical limnology
Before the 1990s, physical limnology in the DSL was mainly directed towards climatology and hydrology, thermal characteristics and heat exchanges, mechanical energy exchanges and thermal hydrodynamics Citation232. While many of these aspects were still under study during the last 15 years, a major focus was on the effects of global climate change on the physical structure of the water column and, owing to its impact on the biotic communities, deep mixing dynamics Citation25.
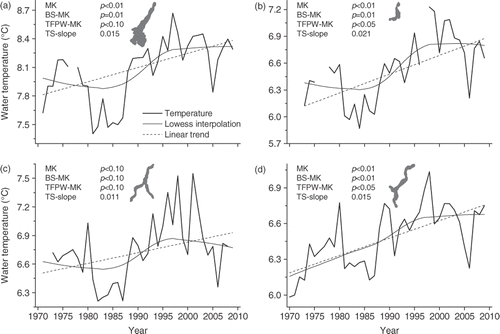
The existence of an increasing and temporally coherent overall trend in the temperature of the DSL was clearly recognised for the first time by Ambrosetti and Barbanti Citation33. They found that the heat content of hypolimnetic waters at the end of the winter months strictly depended on winter climatic conditions (air temperature, solar radiation, wind) and on the quantity of heat reaching the deep layers before and after the onset of thermal stratification. In Lake Maggiore this quantity was around 2–4% of the total heat budget of the lake, which was sufficient to cause a progressive increase in the heat content of deep waters. Similar percentages in the heat transfer were found in the other DSL. It was further noted that the warming of the deep layers of Lake Maggiore occurred in parallel with an increase in temperature of about 1.5°C in its inflows during the 1990s. The conclusion was that the hydrodynamic processes and water temperatures depended on large-scale climatic fluctuations controlled by an ongoing process of climate change. In the Alpine region, temperature changes appear of greater magnitude than the global temperature changes, reflecting an amplification of the global signal in this region Citation260. The increase in temperature in the deep lakes south of the Alps was successively confirmed in a more general study on the trophic development of DSL (see section 3.3; Citation79). The analysis of a long-term data series recorded from the 1970s to 2005 at the time of maximum spring overturn allowed the increase in water temperature to be estimated at around 0.012–0.028°C year−1, which was very close to the warming rate found in other large lakes in Europe and North America, i.e. 0.015–0.030°C year−1 Citation261. An update of these results for lakes Garda, Iseo, Como and Maggiore is reported in . Even considering the occurrence of sharp temperature reductions following full overturn events (e.g. in 1981–1982, 2005–2006), the overall upward trend was still manifest, confirming previous results and temperature slopes (0.011–0.021°C year−1). In the case of Lake Como, however, the overall temperature increase based on the MK, BS-MK and TFPW-MK tests () was at the limit of significance (P < 0.10). Nevertheless, besides the long-term trend, the data were clearly characterised by short-term structures with a periodicity greater than 10 years, highlighting that information relevant to climate change in these types of water bodies need to be based on sufficiently long time series. In this regard, after removing the effect of serial correlation, the upward trend decreased in significance, especially in Lake Garda (P < 0.10).
Figure 3. Volume-weighted mean lake temperature from the beginning of the 1970s to 2009 (updated from Citation79). (a) Garda; (b) Iseo; (c) Como; (d) Maggiore. Measurements were made during spring, at the time of maximum overturn. Data are smoothed using the LOWESS procedure. Dashed lines show the Theil-Sen slopes of the linear trends. For every temperature series, various statistics have been computed: MK, Mann–Kendall test Citation245; BS-MK, Bootstrap-based MK test with 10000 replications Citation245; TFPW-MK, trend-free pre-whitening procedure (TFPW) and MK test Citation246; TS-slope, Theil-Sen's slope estimates (°C year−1) Citation245.
Numerous studies in this period addressed the problem of the factors controlling mixing and deep-water dynamics. A first line of research studied the meromixis of lakes Iseo and Lugano. The decrease in the intensity of deep mixing in Lake Iseo was observed since the mid 1980s, whereas meromixis took place in the mid 1990s. Studying the evolution in time of stability in relation to water temperature, solute and dissolved oxygen concentrations, Ambrosetti and Barbanti Citation17 showed that the increase in total water stability (thermal and chemical) was determined by an increase in the thickness of the surface water layer involved in the annual heat exchange and by an increase of solutes present in the deep layers. The solutes were partially related to the sinking of calcite produced in the photic zone and to their dissolution in the tropholitic layers. A partial deep-water renewal of the lake was documented in 2005 Citation79 and 2006, during severe winters. The absence of complete overturn was even more marked in the northern basin of Lake Lugano, which showed a permanent stratification over the last 40 years because of an increased dissolution of mineral particles and increased water stability in response to strong eutrophication Citation22. After the 1960s, the meromixis determined the complete disappearance of oxygen in the deeper layers, and an increase in the concentrations of reducing chemical compounds such as ammonium, hydrogen sulphide and methane Citation70. During this period, the regular increase in the deep water temperature approached a threshold above which the cooling of the surface water could force overturn, which, nevertheless, was prevented by mild winter temperatures Citation21. The use of environmental tracers allowed detailed documentation of the interruption of meromixis after two consecutive cold and windy winters (2004–2005 and 2005–2006), which destabilised the water column, leading to two strong mixing events Citation22. However, the process was associated with a significant reduction in the trophic level of the lake, due to treatment of sewage in the watershed. In this regard, the meromixis should be considered as favoured by eutrophication, which regulates the fluxes of calcite from the productive layer to the hypolimnion and the CO2 concentrations in the whole water column.
A second line of research in deep-water dynamics considered the transport mechanisms determining the extent of the overturn of hypolimnetic waters. Analysing a long-term data set of physical and chemical data measured in Lake Maggiore from 1951 to 2008, Ambrosetti et al. Citation27 showed that–besides convective mixing–at least other three mechanisms favoured the mixing and oxygenation of deeper waters: i.e. conveyor belt currents, the inflow of colder and denser fluvial water that consequently slide down to the lake bottom, and differential cooling of littoral waters sinking down the lake flanks. While the water mass undergoing convective motion increases in thickness during the winter months, these three mechanisms can contribute significantly to the penetration and mixing of surface waters at depth. As for the inflows, the role played by flood-induced turbidity currents in favouring the circulation of Lake Lugano was studied by De Cesare et al. Citation20. The overall conclusion reached by Ambrosetti et al. Citation27 was that deep water dynamics in the DSL can be driven by a variety of mechanisms depending on the dynamics of atmospheric forcing. From a slight different point of view, Salmaso Citation15 analysed the impact of the winter climatic fluctuations on the extent of the spring vertical water mixing and phosphorus fertilization in Lake Garda. A strong causal relationship was found between the severity of winter and the spring temperature of the upper hypolimnion. In turn, the cooling of the water column strongly controlled the vertical extent of spring overturn and the degree of phosphorus fertilization and phytoplankton growth of the trophogenic layers Citation65,Citation66,Citation86. An attempt was also made to relate the winter climate and mixing dynamics to large scale climatic phenomena Citation15. Nevertheless, the relationship between the winter climate and the North Atlantic Oscillation (NAO) was uncertain, probably because of the location of the lake between different centres of action of the NAO (the Mediterranean region and central and northern Europe).
Apart from a contribution by Buzzi et al. Citation18, who analysed the oscillation periods of surface seiches in Lake Como, only a few numerical applications analysing the thermal regime and water movements in the DSL have appeared very recently. Castellano et al. Citation28 described a numerical study for estimating the spatial distribution of the hydraulic residence time in Lake Maggiore. The model was used to simulate the velocity distributions, the paths of different massless markers and their residence times within the lake. The estimated residence times were in accordance with the mechanisms of mixing and deep-water oxygenation described by Ambrosetti et al. Citation27. A different logic animated the work of Morillo et al. Citation19, which originated within the framework of a pilot experiment aimed at removing polluted waters from a coastal margin of Lake Como by using a downward-pointing impeller. A field study conducted using rhodamine dye introduced into the impeller was used to validate a three-dimensional numerical hydrodynamic model. Successive simulations showed that a reduction in the amount of pollutants adjacent to the City of Como by half could be reached within 2 weeks with the use of six impellers.
A large number of papers addressed the question of relating heat content, thermal structures and the depth of the thermocline of a large number of Italian lakes (including the DSL) to their morphometric characteristics Citation29–31,Citation34,Citation35. Lake flood episodes and the climate of Lake Maggiore watershed were analysed in other specific papers Citation23,Citation24,Citation26.
3.2. Geochemistry
Geochemistry was the object of many investigations, particularly in Lake Lugano. An important research line regarded isotopic geochemistry, with the quantification of different isotopes (Cesium, Boron, Berillium, and Uranium) in the pelagic zone and sediments Citation37,Citation42,Citation46,Citation47,Citation50,Citation61,Citation62. In this field, specific studies were made to assess the risk of radioactivity on human health. Rusconi et al. Citation37 evaluated the possible impact of some unused conventional bombs which had been dropped into Lake Garda in 1999, during the Kosovo conflict. They found 238U concentrations typical of natural samples, ranging from 10 to 20 mBq l−1. Moreover, 234U/238U ratios were never significantly less than 1, as would have been the case in samples heavily contaminated by depleted uranium.
In the fields of biogeochemistry and organic geochemistry, many studies analyzed the processes involved in the mineralization and isotopic composition of carbon, nitrogen and oxygen in settling organic matter Citation41,Citation51–56. In particular, analysing the evolution of δ
13C and δ
15N of sedimenting particulate organic matter (SPOM) of Lake Lugano from 1985 to 1998, Lehmann et al. Citation55 demonstrated a parallel decrease of δ
13C and phosphorus loading, suggesting that δ
13C in SPOM was a good indicator of P availability. This was explained considering the control of available on algal biomass, and thus on CO2 reduction, and with the relationship existing between the intensity of aqueous CO2 depletion and δ
13C values of autochthonous organic matter. Conversely, changes in total phytoplankton biomass did not seem to affect δ
15N values.
Due to their importance in the co-precipitation processes involving phosphorus and in the determination of density profiles, a few studies analyzed the processes controlling the formation and dissolution of calcite particles in the epilimnion and hypolimnion Citation36,Citation45. In Lake Garda, Salmaso and Decet Citation36 found that the reduction of Ca2+ in the whole lake could be estimated as between 23,800 and 17,500 t year−1, which corresponds to a precipitation rate between 50 and 65 g Ca2+ m−2 year−1. These values are lower than those found in other two deep eutrophic lakes (Lake Lugano and Sempachersee; about 170 g Ca2+ m−2 year−1 Citation45), confirming the dependence of Ca precipitation on phytoplankton productivity (see section 3.1).
In Lake Lugano, other studies considered the chemistry and distribution of reducing compounds Citation40,Citation57, the transport of suspended matter in the hypolimnion Citation39,Citation43, and the dissolved silica budget in the North basin Citation49.
3.3. Chemistry and eutrophication: different responses due to different management strategies
The studies on water chemistry carried out on the DSL were directed mainly at evaluating lake trophic status, as eutrophication has always been the main cause of deterioration for these water bodies. Nevertheless, other important uses of the chemical data are in evaluating the extent of the spring vertical mixing of the waters, and in investigating the interactions between biological processes and effects on water chemistry, such as the formation of calcite in the productive layers and its subsequent partial dissolution during sedimentation (see section 3.2). As biological production in the DSL is essentially P limited, P concentrations are a reliable indicator of lake trophic status. Therefore all the processes related to P (input from the watershed, consumption in the epilimnion, sedimentation, release in the hypolimnion and storage in the sediments) have been of major interest. The concentrations of P in water, both in the reactive and total forms (SRP and TP), are a reliable indicator of the excess of P loading from the watershed reaching the lake; on the other hand, the increase or decrease of P concentrations indicate the effects of remedial measures. The long-term series of P concentrations documented an increase in the trophic level of Lake Lugano and of the remaining lakes since the 1960s and 1970s, respectively Citation76. At the end of the 1980s eutrophication ranged between the oligo-mesotrophy of Lake Garda to the eutrophy-hypereutrophy of Lake Lugano, whereas Lake Maggiore and lakes Como and Iseo showed meso-eutrophic and eutrophic conditions, respectively (). The remedial measures produced positive effects for lakes Maggiore, Como and Lugano since the early 1990s, but not for lakes Iseo and Garda. These modifications were documented and commented upon in several papers, e.g. Citation63,Citation64,Citation68,Citation71,Citation75–79.
Figure 4. Volume-weighted average concentrations of (a) total phosphorus in the whole lake and (b) dissolved oxygen in the layer below 200 m from the beginning of the 1970s to 2008. The series are based on measurements carried out at the time of maximum overturn during spring (updated from Refs Citation75,Citation76,Citation79).
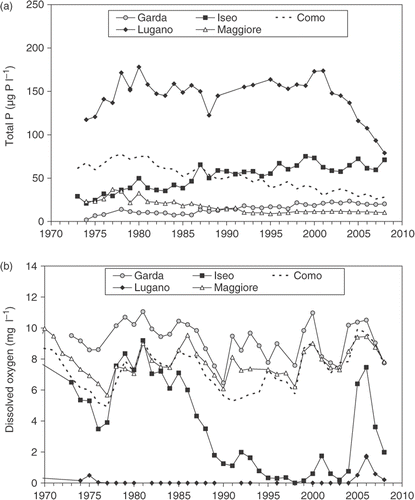
Among the other chemical variables, of particular interest were nitrate and dissolved oxygen. Nitrate is the most important N species present in lake waters and for several years it was associated with P as a cause of eutrophication. Nitrate showed a relevant increase since the 1960s in lakes Maggiore, Como and Iseo, while remained almost stable in Lake Garda. In Lake Lugano nitrate showed a more complex dynamic in relation to the anoxic conditions of deep waters Citation70,Citation79. Other specific studies, based on a multi-year analysis of the sources of nitrogen to the lakes, clearly showed the importance of the atmospheric loads reaching the lake and watershed surface in precipitation, in addition to the N load produced by human activities in the watershed Citation73. The relative contribution to the total N load was estimated as being between 65 and 75%, depending on the interannual variability of the amount of precipitation Citation74. The increase in nitrate in the DSL could be related to the atmospheric cycle of ammonium, deriving from zootechnical activities and manure, and nitrogen oxides, resulting from car exhaust emissions and, more generally, from high temperature combustion Citation262. The absence of a positive trend in Lake Garda can be related to the lower atmospheric load and to the higher water retention time, which could allow a relatively higher incidence of biological processes consuming nitrogen. On the other hand, due to reduction processes, nitrate concentrations strongly depend on the oxygen concentrations in the deeper waters, as clearly shown in Lake Lugano Citation70 and, less manifestly, Lake Iseo Citation67.
As for nitrates, it is interesting to observe that in lakes Garda and Lugano, the epilimnetic nitrate concentrations during the summer months could reach far lower values than in the other lakes (<30 µg N l−1), suggesting that a limitation of phytoplankton growth in the top few metres of the water column in these two lakes during the periods of maximum nutrient consumption may be determined by N as well as P Citation82,Citation103.
The oxygen profiles over the water column were always used, together with temperature and a few other variables (e.g. pH, conductivity, nitrates, silica), as an indicator of mixing depth at the time of maximum late winter and early spring overturn, see, e.g. Citation15,Citation17,Citation36,Citation72,Citation76,Citation79. An update of the long-term concentrations of O2 considered in previous works is reported in . The use of this variable still represents a useful and synthetic way to follow the mixing dynamics in deep lakes. In particular, the long-term trends clearly showed the partial and almost complete oxygenation of the deeper layers of lakes Lugano and Iseo in 2005–2006.
Other aspects related to the chemical studies of the DSL are the chemical budget of nutrients and main ions originating from the watershed, and atmospheric concentrations, mainly focusing on P loads as the main cause of eutrophication. Continuing studies of these budgets are available since 1978 for Lake Maggiore and Lake Lugano. Studies lasting 1 or 2 years were performed on lakes Como and Iseo, while no definite budget studies are available in the case of Lake Garda. Results showed a regular decrease in the P load in lakes Maggiore and Lugano, in agreement with the decrease in P in lake water and the decrease in lake water production Citation75. As for lakes Iseo Citation69 and Como, the short period of measurement only allowed evaluation of the agreement of the relationship between annual P load and lake P concentrations, and of its agreement with the OECD Citation263 models.
3.4. Phytoplankton
Besides physical and chemical limnology, phytoplankton and zooplankton (see section 3.5) represent another two important topics in historical development of limnology in Italy. A review of the main studies on planktonic communities before the 1990s was carried out by Manca et al. Citation254 and Ambrosetti et al. Citation264. During the last 15 years, the main studies on phytoplankton were directed toward understanding the effects of eutrophication and oligotrophication processes on the composition, structure and dynamics of algal assemblages, and on the temporal dynamics of algal communities. Only in very recent years have studies also been directed towards understanding the role of climatic fluctuations and global change in shaping phytoplankton assemblages.
The main changes in phytoplankton assemblages during the eutrophication and oligotrophication phases of Lake Maggiore during the last 30 years were the object of many studies (see Citation96), the first of which was conducted by Ruggiu et al. Citation93. A number of synthetic variables expressing variations in abundance and morphological phytoplankton features allowed two phases in the long-term process of oligotrophication to be identified. Despite the reduction of lake phosphorus concentrations (see 3.3; Citation99) between 1978 and 1987, the phytoplankton structure showed no significant changes. In the following years, between 1988 and 1995, the phytoplankton underwent noteworthy changes, including an increase in biodiversity, a reduction in biovolume and a decrease in average community cell size. The decrease in average cell volume and its close association with the oligotrophication of the lake was further confirmed by successive analyses Citation102. Modifications in these synthetic features were followed by a strong reorganization of the community Citation93,Citation102. Many persistent large taxa tolerating a wide spectrum of trophic conditions showed a consistent biovolume reduction (e.g. Asterionella, Fragilaria, Aulacoseira, Cyclotella, Planktothrix, and Ceratium). Other alterations were observed in species more closely related to the trophic changes. Synedra, Melosira, Stephanodiscus and Microcystis–all important during the first phase–were followed by the gradual development of Aphanothece, Chrysochromulina, Snowella and thin Oscillatoriales.
Similarly, the slow process of eutrophication in Lake Garda was paralleled by many qualitative and quantitative changes Citation83. Compared with the pristine oligotrophic conditions, other new, important species appeared in the lake during the 1970s, such as Planktothrix and Mougeotia, which are now among the dominant taxa in the community. During the 1980s, new cyanobacteria (Planktolyngbya and Anabaena) also appeared. Changes in the phytoplankton on a smaller temporal scale (1993–2007) were analysed by Salmaso Citation86. Following the slight and continuous increase in phosphorus concentrations, after a first long period of dominance by conjugatophytes (Mougeotia) and diatoms (Fragilaria), the recent phytoplankton biomass in Lake Garda was mainly sustained by cyanobacteria (Planktothrix). Superimposed over this pattern, other modifications were directly or indirectly induced by fluctuations in the winter climate, mediated by the deep mixing dynamics at spring overturn (see also section 3.1) and by the thermal structure of the water column during the growing season.
A further line of research concentrated on understanding the processes and mechanisms controlling the selection of phytoplankton assemblages along temporal gradients. In particular, a number of studies highlighted the higher regularity and predictability that characterise the annual cycles of phytoplankton development in the largest DSL compared with smaller and shallower ecosystems, e.g. Citation80,Citation82,Citation84,Citation86,Citation88,Citation97,Citation100. The regular annual cycles in the DSL are favoured by a complex of causes which include the seasonal variability of the climate, the vertical mixing of the water column from late autumn to early spring, the formation of extended and stable epilimnetic layers, and the high inertial characteristics of large lakes, which minimise the effects of local and stochastic perturbations (meteorological events, hydrological inputs) Citation84.
An important aspect investigated in Lake Maggiore was primary production (PP). In line with the decrease in algal biomass after the recovery of the lake, the mean annual PP showed a clear decrease from 300 g C m−2 year−1 during the 1970s to 190 g C m−2 year−1 in the period 1994–1995 Citation94. More recent measurements made in 2002 confirmed the recent oligotrophic state of the lake (150 g C m−2 year−1; Citation101).
Other studies in individual lakes addressed specific ecological topics, which included the definition and use of morpho-functional phytoplankton groups in the comparison of communities across lakes Citation85; autoecological studies of diatoms Citation90,Citation91,Citation95; and the assessment of carbon assimilation and phytoplankton growth rates in Lake Maggiore and other water bodies of contrasting trophic status Citation98.
In recent years, the phytoplankton composition and structure across the deep southern subalpine lakes were the object of two synoptic studies Citation103,Citation104 based on a group of concurrent investigations made on the individual water bodies by Buzzi Citation89, Morabito et al. Citation96, Salmaso Citation83; Garibaldi et al. Citation87 and Simona Citation92. With a few exceptions, the lists of the more abundant species were highly comparable. The major differences were between the eastern lakes (Garda and Iseo) and the western ones (Lugano and, partly, Maggiore), with intermediate characteristics in Lake Como. Nevertheless, the most apparent differences were due to quantitative changes in the dominance relationships among the constituent taxa and only secondarily to compositional changes. At the community level, it was possible to recognize common patterns in the replacement of the seasonal assemblages, which were mainly represented by the development of large diatoms during spring, a more diversified community in summer (mostly Zygnemales, Peridiniales and Ochromonadales), and cyanobacteria and Chlorococcales in summer and autumn. Of the cyanobacteria, the most representative species was Planktothrix rubescens, which was present with the highest biovolumes in lakes Iseo and Como. Of the remaining cyanobacteria, it was stressed that a consolidated difference was the strong development of Aphanizomenon flos-aquae in Lake Lugano and its presence in Lake Maggiore, and the occurrence of oligotrophic blooms of Anabaena lemmermannii in lakes Garda and Iseo Citation82,Citation103. It is interesting to observe that in very recent years the blooms of Anabaena have rapidly extended also to lakes Maggiore (2005) and Como (2006). At the same time, a spreading of blooms of Microcystis aeruginosa was observed in lakes Como, Iseo and Lugano. The results obtained within the framework of these investigations were used to define new ecological indices based on phytoplankton (PTIs indices) as suggested by the European Union Water Framework Directive Citation104.
3.5. Zooplankton
Excluding two sporadic contributions on Lake Garda, in the last 15 years zooplankton was the object of intensive studies exclusively in Lake Maggiore. Paralleling what was extensively done for phytoplankton, a few studies analysed the consequences of the long-term lake oligotrophication process on the zooplankton community Citation107,Citation112,Citation116. de Bernardi and Canale Citation107 described in detail the modifications of the zooplanktonic biocenosis from 1948 to the beginning of the 1990s, demonstrating important differences due to a dominance of cladocerans and rotifers in the period of maximum eutrophication, and a returning of the community to conditions typical of the 1950s–with a dominance of copepods–after the first phases of the oligotrophication process. As for the possible impact of allochtonous species on the aquatic ecosystems, it is worth observing that the disappearance of Sida cristallina and Heterocope saliens at the beginning of the 1960s was correlated with a significant development of an allochtonous coregonid introduced into Lake Maggiore in the middle of the 1950s. By contrast, new species of Daphnia (D. galeata and D. cucullata) made their appearance in the lake since the end of the 1980s. These changes were followed by modifications in the food webs. Manca and Ruggiu Citation112 and Manca et al. Citation116 showed that Daphnia population dynamics in Lake Maggiore in the middle of the 1990s appeared to be mainly controlled by Bythotrephes predation in June, which constituted a quite different situation from that documented at the beginning of the 1970s, when Leptodora was the prevalent predatory cladoceran. In two subsequent works, Manca et al. Citation120 and Manca and DeMott Citation124 related the 10-fold increase in the abundance of Bythotrephes from the middle of the 1980s to the beginning of the 1990s to a rise in lake temperature, which could have affected Bythotrephes reproduction and population growth. In the following years, Bythotrephes further increased as water temperature continued to increase. The alteration of the thermal stratification regime of the lake could also have caused an increase in the depth and duration of a dark refuge of relatively warm water, which might be able to support growth and development while minimizing the risk of predation by visually orienting fish predators. The results provided evidence of the complex modifications in the population density and phenology of key planktonic species and functioning of the pelagic food webs triggered by climatic changes.
Testing the hypothesis of an overall eutrophication-like effect of climate warming on zooplankton, Visconti et al. Citation123 compared the structure of the zooplankton community in the warmest years of the recent oligotrophic phase with that observed during the non-warm years of oligotrophy, and during years of the mesotrophic phase. They observed an abrupt increase in zooplankton abundance during 2003, one of the warmest years, particularly due to a larger development of Daphnia, which reached values typical of the mesotrophic phase. These abundances strongly differed from those observed during cooler years since re-oligotrophication. The results supported the hypothesis that warming may result in an eutrophication-like signal.
From a community perspective, one research line was concerned with the impact of predation on zooplankton, with specific studies considering the trophic relationships between Daphnia and ciliates Citation119, the role played by zooplanktivorous fish and invertebrate predators on the population dynamics of Daphnia Citation122, and the relationship between the feeding basket of Leptodora and the size of the prey that can be captured Citation108. Along the same line of research, studies were also carried out on the first (1992) identification of exotopic protrusions and ellobiopsid infection in copepods Citation118.
The recent colonization of Lake Maggiore by Eudiaptomus gracilis and its consequences for the indigenous and abundant E. padanus populations were discussed by Riccardi and Rossetti Citation121. It was postulated that, provided E. gracilis were permanently established, Lake Maggiore could become an invasion hub, facilitating further spreading owing both to natural and human-mediated connections to other water bodies in the region.
In Lake Maggiore, other specific studies regarded the role of zooplankton in biogeochemical cycles and in the transport of potential energy and chemical elements Citation111,Citation113,Citation114,Citation117, and population dynamics in rotifers Citation110.
As for Lake Garda, a comparison of the historical results obtained before and in the middle of the 1990s did not seem to demonstrate substantial shifts in the composition of the dominant zooplankters Citation105. The only differences were due to the disappearance of Sida cristallina since the 1950s, and the appearance of new rotifers since the 1970s and 1980s. Of particular interest was the appearance of ‘black spots’ spreading throughout the lake between July and early September 2004 Citation106. The phenomenon was due to a strong development of Stentor amethystinus, a ciliate, and its symbiotic alga, Chlorella sp.
3.6. Microbial ecology
Similarly to zooplankton, the study of the smaller constituents of the aquatic food webs was carried out almost exclusively in Lake Maggiore. The rationale of the studies carried out before the 1990s was synthesized by Bertoni and Callieri Citation265. In line with the discoveries made during the previous 15 years Citation266, this research line started with the consideration that most of the organic carbon was present in the smaller planktonic particles and in the dissolved fraction Citation141.
A number of papers dealing with the constituents of the microbial loop focused on heterotrophic (HPP) and autotrophic (APP) picoplankton, e.g. Citation126,Citation127,Citation142. Using a trophic gradient approach Citation133, it was possible to examine the influence of resource availability on picocyanobacteria growth and abundance. As for Lake Maggiore, the decrease of phytoplankton biomass in the recent oligotrophication phases (see section 3.4) was followed by an increase in the abundance and production of picocyanobacteria. More generally, it was argued that the success of picocyanobacteria in oligotrophic lakes could be achieved only if additional conditions different from low ambient concentrations of nutrients were met, including low grazing and competition, and favourable light climate. Therefore, the success of these organisms in oligotrophic lakes should not be considered as a certainty, but a potentiality.
Other studies considered the relationships between nutrient enrichment and bacterioplankton Citation140, as well as the size distribution of POC Citation131, the implications of the formation of aggregates and microcolonies in chroococcoid cyanobacteria Citation134, and the horizontal distribution of organic carbon and picoplankton Citation137.
A number of investigations considered the functional and physiological characteristics of picoplanktonic organisms. The study of the photosynthetic characteristics and genetic diversity of Synechococcus populations under various light and mixing conditions showed that samples from different depths and with different nutrient concentrations hosted different strains characterised by different genetic fingerprints and photosynthetic properties Citation139. As for the production of organic matter accounted for by the smaller photosynthetic organisms, it was estimated that the annual average productions of autotrophic picoplankton in 1994 and 1998 were 16 and 41 g C m−2 year−1, which represented a total phototrophic production of between 10 and 20%. However, in the seasonal periods of high P limitation, APP carbon fixation (13 mg C m−3 h−1) in the upper epilimnion (3 m) exceeded that due to larger photosynthetic fractions Citation135. Furthermore, it was shown that, under natural UVB irradiance, the activity of picoplankton (both HPP and APP) and of the greater autotrophic fraction (>2 µm) was not significantly inhibited Citation130.
With a more general approach, further studies carried out in Lake Maggiore Citation129 showed that the total amount of carbon fixed per unit of chlorophyll-a (Chl-a) increased from 1981 (meso-eutrophic phase) to 1994 (oligotrophication period). These changes were reflected in a change in the qualitative characteristics of the organic matter, with an increase in the POC/Chl-a ratio, suggesting a more efficient utilization of energy and nutrients during the oligotrophic phase of the lake.
A specific research line studied the implications of grazing on picoplankton Citation132. In particular, the size of picocyanobacteria was inversely correlated with the clearance rate of heterotrophic micrograzers, indicating a less efficient cell uptake by protozoa when picocyanobacteria are composed of larger cells. A preliminary annual energy balance demonstrated that 80% of the carbon produced by picocyanobacteria could be taken up by protozoa Citation136.
3.7. Benthos
In the only available review of the macrobenthos of the deep lakes, Bonacina et al. Citation267 limited their analysis to a specific period (1963–1973), highlighting the fact that many other data collected sparsely after the 1970s were still unpublished. After the 1990s, the benthos was still among the more poorly studied communities in the DSL. At least two reasons may have favoured this negative bias. Many investigations were directed towards one of the most critical invasive species, Dreissena polymorpha (zebra mussel), which was detected in Lake Garda for the first time in the late 1960s. A second reason is that many efforts were directed towards the effects of micropollutants on specific target species (mainly D. polymorpha; see below, section 3.9). Overall, these applied issues contributed to shifting attention towards population ecology and ecotoxicology, at the expense of community ecology.
Investigations covering diverse autoecological characteristics of Dreissena have been carried out in different DSL Citation147–150. More specifically, the role in nutrient cycling played by Dreissena was investigated in the western basin of Lake Como Citation148. The entire population was able to filter epilimnetic waters 2.1 times per year, transferring 2.9×104 t year−1 of pseudofaeces to the sediments. In a recent study aimed at tracking human-mediated dispersal pathways, the genetic structure of three populations from Lake Garda was compared with that of two populations from Lake Constance Citation145. In order to define phylogeographic relationships, the results of the five populations were compared with all the available sequences available from GenBank (the National Institutes of Health genetic sequence database). All the Italian samples joined with both German and mid-European samples, suggesting a mixed origin of the Italian Dreissena. Further studies concerned other larger molluscs, such as the populations of Unio pictorum mancus and Viviparus ater in Lake Maggiore Citation151,Citation152.
A few studies have focused on recent new introductions of shrimps and molluscs in Lake Garda (Dikerogammarus villosus Citation144; Corbicula fluminalis and C. fluminea Citation146), and on the identification of new species of Gastrotrichs (Lake Garda Citation143).
3.8. Fishes
Studies on fishes have been carried out almost exclusively in Lake Maggiore. A few investigations concerned the management of fish resources Citation156,Citation157, ecology Citation155, physiology Citation158,Citation161, the implications of the introduction or invasion of new species Citation159,Citation160, and population genetics Citation153,Citation154. Implications of the contamination of fishes by persistent organic pollutants (POPs) will be considered in the next section. As for the contribution of genetic studies, evidence was proposed to support the hypothesis that Salmo carpio (a species endemic to Lake Garda) issued from a recent hybridization of Salmo marmoratus and S. trutta fario Citation153. Further, significant differences were found between the natural populations of Salmo of the Po basin and three of the major Italian fish-farm strains, which were of Atlantic origin. The ‘natural’ populations showed introgression rates ranging from 0 to 70% due to contamination by stocking.
3.9. Ecotoxicology
The qualitative and quantitative determination of chemical pollutants and their impact on the DSL ecosystems have been the object of a large number of investigations, particularly in Lake Maggiore (). A lot of studies examined the presence and distribution of many POPs, such as dichlorodiphenyltrichloroethane (DDT), e.g. Citation186,Citation192,Citation196; polychlorinated biphenyls (PCBs), e.g. Citation203,Citation208,Citation211; polycyclic aromatic hydrocarbons (PAHs), e.g. Citation193,Citation197,Citation198; polybrominated dyphenil ethers (PBDEs) Citation203,Citation204,Citation206,Citation207; polychlorinated dibenzodioxins (PCDDs, dioxins), e.g. Citation203,Citation205; perfluorooctanesulfonic acid (PFOS), perfluorooctanoic acid (PFOA) and nonylphenol Citation201. Of particular concern is still the pollution by DDT in Lake Maggiore due to its production in a chemical plant located near the main influent of the Baveno Bay until 1996. Even although the DDT pollution did not compromise the use of the lake water for drinking or bathing purposes, health authorities still prohibited fishing and the eating of many fish species Citation72,Citation191. After a heavy flood in autumn 2000, DDT concentrations increased due to the transport of insecticide from contaminated sediments still present in the area of the closed chemical plant. This was paralleled by a sharp increase in PCB pollution not due to the flood, but probably to improper release of contaminated sediments from numerous dams located in the watershed. The high concentrations of POPs represented a danger for the water community, and a possible risk to human health due to fish consumption Citation191. In Lake Maggiore, the existence of high concentrations of DDT and its metabolites in fishes was proved in several studies. Bettinetti et al. Citation194 showed that concentrations of DDT were very close to, and sometimes exceeded, the Italian limit for human consumption (0.05 and 0.1 mg kg−1 w.w. of total DDT for fishes with lipid content of >5% and within the range of 5–20%, respectively). They concluded that the environmental quality standard of 0.2 ng l−1 in water could not guarantee the suitability of fish for human consumption. Volta et al. Citation210 showed that DDT and PCB concentrations in Coregonus macrophthalmus were dependent on season and fish age. They described a 2-fold increase in the lipid-normalised concentrations of DDTs and PCBs across season and also across age, resulting in an overall 4-fold increase. Seasonal variations in POPs content were related to the ecophysiological cycle of fishes, while the effect of the fish age was explained taking into account biomagnification mechanisms (see also Citation173). At a higher trophic level, DDT and PCB monitoring of birds’ eggs in Lake Maggiore revealed a situation of high contamination in comparison with other aquatic environments Citation185. Successive studies considered the birds’ eggs contamination by these two POPs in lakes Maggiore and Garda Citation217, and carbofuran (another pesticide) poisoning in mallard ducks in Lake Lugano Citation180. The evaluation of risk for human health of PCBs and PAHs due to fish from Lake Iseo was studied by Binelli and Provini Citation174.
The long-term risks posed by POPs were highlighted also by Citation216. This study related the sharp increase of DDT and its metabolites observed in lakes Como and Iseo in 2005 to the meltwater rich in pollutants released by retreating glaciers. In particular, specimens of Alosa fallax lacustris collected from Lake Como showed DDT levels of 0.12 mg kg−1 (w.w.), which exceeded the Italian limit for human consumption.
The use of zebra mussel for lake biomonitoring was the object of different studies Citation199–200,Citation214,Citation218. Based on the linkage between bioaccumulation and biomarker data, Binelli et al. Citation215 defined site-specific environmental quality indices for man-made chemicals. The application of these tools allowed three different degrees of xenobiotic contamination of the DSL to be distinguished. Lake Lugano had negligible pollutant levels and no effects on enzyme activities, while lakes Garda, Iseo and Como showed homogeneous poor water quality. The ecological status of Lake Maggiore was considered at risk due to the presence of some xenobiotic point-sources and the heavy DDT contamination revealed since 1996. The endocrine disrupting properties of DDT were confirmed in specific studies carried out on Dreissena populations in the lakes Maggiore Citation187,Citation190 and Iseo Citation175. In particular, histopathological analyses performed on mussels collected from Lake Iseo during the sharp increase of POPs (2005–2006) showed a disrupting action of DDT on the mechanisms involved in sperm release, as well as disturbances in the gametogenic phases of the ovary Citation175.
Further investigations were carried out on diverse ecotoxicological aspects. Among the others we can cite: metal and trace element concentrations in the environment and in aquatic biota, mainly molluscs: e.g. Citation181,Citation188,Citation189,Citation202,Citation209,Citation213; POPs in sediments Citation182; the mutagenic activity of lake water used as drinking-water supply Citation212; the use of advanced technologies for organic micropollutant removal from lake water Citation178; and the development of models for the estimation of temporal emissions of DDT to Lake Maggiore Citation195.
3.10. Extending the temporal perspective: paleolimnology
The most complete paleolimnological studies were carried out in Lake Maggiore. In this lake, Marchetto et al. Citation221 demonstrated a good level of coherence between diatom assemblages in sediment cores and documentary data on phytoplankton and hydrochemistry. In particular, they clearly documented the displacement of ‘small Cyclotella species’, typical of the oligotrophic phase, by more meso-eutrophic species (e.g. Tabellaria flocculosa). Similarly good was the prediction of total algal biomass from the profile of a few paleopigments during a certain period of the lake's development. Interpretation of the results obtained with chironomids was less straightforward. Lake eutrophication did not appear to have a clear and immediate impact on the composition of this group. Most of the sharp changes in the littoral chironomid assemblages appeared to be linked to major fluctuations in lake level. In the same study Citation221, the results obtained in Lake Maggiore were compared with a preliminary investigation of fossil diatom assemblages recorded in short cores collected in lakes Garda, Iseo and Como at the beginning of the 1990s. During oligotrophic phases, Cyclotella spp., which were the dominant taxa in the oldest sediments, were successively replaced by more eutrophic and mesotrophic taxa, such as Stephanodiscus spp., Aulacoseira spp., Fragilaria crotonensis, Asterionella formosa and T. flocculosa. The authors concluded that the trophic histories of these lakes, as reconstructed from the diatom remains, were basically the same as in Lake Maggiore. Along the same line of research, sedimentary diatoms were used to reconstruct the long-term development of phosphorus Citation220.
More recently, Manca et al. Citation219 reconstructed the trophic dynamics of Lake Maggiore using subfossil cladoceran data from the beginning of the 1940s to 2002. The changes in the total abundance and the proportion of planktonic cladocerans clearly tracked the major changes that had occurred in the trophic characteristics of the lake, and the changes that had occurred in the composition and abundance of the fish populations due to the introduction of allochtonous species and to the ban on fishing because of the DDT pollution.
3.11. Extending the spatial perspective: remote sensing
The areal extension of the DSL may render difficult the evaluation of the representativeness of data collected in the pelagic zone or along the shoreline. Remote sensing has proved to be useful in large lakes research Citation268. In the DSL, remote sensing was used to investigate selected water quality variables in lakes Garda and Iseo, and macrophytic vegetation in Lake Garda. A specific research line was directed at defining algorithms for the determination of optical properties, algal biomass concentrations and tripton using satellite data (Landsat, Earth Observing-1, Envisat) Citation222–224,Citation226,Citation228–230. In the most recent study, Giardino et al. Citation226 compared the chlorophyll-a and tripton concentrations derived from Hyperion (hyperspectral sensor) records with concurrent in-situ data collected in Lake Garda both at fixed stations and by continuous flow-through systems. The correlation coefficient between the concentrations obtained at the fixed stations and the Hyperion derived values was 0.77. For tripton, the validation was problematic due to resuspended material; however, if the sampling data from the resuspension zones were excluded, the correlation coefficient between the fixed stations and Hyperion-derived concentrations increased from 0.48 to 0.75.
A second line of research was concerned with the monitoring of macrophytes, i.e. submerged vegetation Citation225 and reeds Citation227. A study of the distribution of submerged vegetation in the littoral zone of the Sirmione Peninsula (in the southern basin of Lake Garda) documented a marked decrease in macrophyte cover from 72% (1997) to 52% (2005), with a concomitant increase in sandy substrates Citation225.
3.12. Other topics
Other important limnological topics considered in these last 15 years have included lake morphometry and geology; hydrology and hydrochemistry in the watershed areas; and birds and aquatic parasites (for complete references, see ).
Among the hydrological investigations, a study of the influence of the inflowing River Sarca in the northernmost section of Lake Garda by isotopic measurements in water samples showed no detectable effects and no vertical stratification of isotopes Citation4. Strictly linked to ecological economics, a few studies Citation5,Citation7,Citation8 addressed the important problem of water availability and the allocation of water resources among competitors in the Adda River basin, especially during situations of water scarcity (such as in 2003, Citation5). At the other extreme of the hydrological gradient, Luino et al. Citation12 presented the results of a model aimed at evaluating economic losses from the impact of floodwaters on exposed elements; the model was tested on a small watercourse flowing into Lake Maggiore, which was recently affected by a flash flood (the Boesio Stream).
From a purely ecological perspective, microparasites and macroparasites are considered crucial elements in the control of host populations Citation269. However, the studies carried out in the DSL mostly addressed important problems linked to human health. These included the quantification of populations and the identification of mechanisms responsible for the persistence of Enterococcus faecalis in water bodies Citation167,Citation168, and the study of Diphyllobothrium latum, associated with the reappearance of Human Diphyllobothriasis in localised areas of the DSL due to the increasing consumption of raw or undercooked fish because of the increasing influence of oriental cuisine Citation169–172.
4. Research gaps, new directions, perspectives
Synoptic analysis of the available peer-reviewed limnological literature has allowed us to acquire an overall picture of the most advanced research carried out during the last 15 years in the deep southern subalpine lakes. A few conclusions can be drawn. (i) Limnological research in the DSL is strongly linked to the concept of scientific monitoring, while experimental studies and modelling are still confined to specific niches; as a consequence, present limnological research is strongly biased towards specific research areas, while other research areas require urgent improvement. (ii) Integration of different disciplines is hampered by the fragmentation of studies on different compartments and by a general, reductionist approach. (iii) Integration of studies and synoptic analyses on a macro-regional scale have been carried out only for specific research areas. (iv) Huge differences exist both in the number and diversity of investigations carried out in the different DSL.
(i) There is no doubt that most of the limnological research conducted during the last 30 years has been fostered by an urgent need to find effective solutions to the problems connected with eutrophication and pollution, and by monitoring programmes. As in other countries, this more applied research has undoubtedly contributed a great deal to fundamental limnology Citation270. In the case of long-term monitoring programmes, these activities have represented a triggering nucleus for the establishment of long-term ecological research projects. The legacy of this recent past has been responsible for promoting the investigations conducted on a few DSL (Garda, Maggiore) within the framework of the Long Term Ecological Research (LTER) network. The establishment of long-term monitoring programmes provides researchers with a more solid foundation for future environmental decisions Citation271. The challenges of LTER are to achieve a predictive understanding of temporal and spatial patterns and variability Citation272,Citation273. Besides data collection, the scientific monitoring includes interpretation, modelling and experimental manipulation of an ample array of variables in order to identify the environmental stressors typical of different ecosystems on different temporal scales, and the degree and quality of interactions among these stressors. In this regard, the physical, chemical and biological (phytoplankton and, to a lesser extent, zooplankton) data collected in the DSL since the beginning of modern limnological analyses have allowed the effects of human impact and different eutrophication phases to be sufficiently determined. Nutrient availability and climate change–along with organic pollutants–still represent the main ecosystem stressors linked to human activity in the DSL. The effects of the interactions, covariations and synergies between nutrients, climate and other stressors remain less well understood. An important role in remedying this lack of understanding may be played by lake models. Their use in the DSL began only very recently, and only on specific ecosystem compartments, essentially in the field of physical limnology. Both the integration of simple mechanistic models and the use of complex modelling represent a way to improve predictive ability and, at the same time, to formulate and test hypotheses.
Rapid adaptation of research is required in dealing with new environmental emergencies. The causes of the rapid proliferation of substantial cyanobacterial blooms in the whole group of large lakes south of the Alps are still poorly understood, and the fraction of toxic strains present in the populations as well as the diversity, quantity, distribution and seasonality of cyanotoxins remain largely unidentified. A thorough knowledge about the nature and quantity of algal toxins and their fate in the environment and trophic webs is a necessary prerequisite for effective risk assessment and management of water resources. Similarly, the impact of the appearance of new invasive species on the DSL has been overlooked. It is worth noting that, apart from a few studies, no experimental research has been conducted so far on the effects of successful invaders such as D. polymorpha and other smaller molluscs, or new exotic shrimps (D. villosus). Besides the use of classical population and community ecology concepts, the adoption of recent rapid improvements in analytical methods in the field of genetics and metabolomics could add new insights and interpretative elements Citation274, allowing the study of organisms to include the intimate mechanisms controlling adaptation and–ultimately–the evolutionary features of populations and communities. These viewpoints, which encompass evolution, biogeography, phylogeography and systematics, are important for the study of biodiversity and the assessment of losses in biodiversity Citation237.
(ii) Twenty years ago, Peters Citation272 observed that the search for the components and the mechanisms of an ecosystem has fragmented limnology into expert, specialised fields, with limited interaction and negligible competition. It is difficult to disagree with this view, even if its judgment may be softened by taking into account the present greater specialization which is coupled–in many cases–with a necessary knowledge of advanced methodologies, both analytical and computational. The idea behind this view, i.e. that the different component properties may be put together to understand the whole system from them, has been largely disputed. Due to the result of interactions and feedback, ecosystem properties may differ appreciably from the sum of its elements, and therefore the properties of complex systems cannot be understood by looking solely at the single constituent parts Citation270,Citation275. It seems commonplace to bring up these observations yet again in this review, but it is worth underlining that, whereas a certain number of studies in the DSL have addressed the relationships between nutrients and planktonic communities, only a very few have considered the interactions between planktonic organisms, and between nekton, benthos and plankton. It seems that this situation arose not only from the idiosyncratic character of each individual discipline and the general consensus about the importance of resources availability, but also from an intrinsic weakness in defining quantitatively competition and predation processes from exclusive field observations. Therefore, the general view is that of a limnological science strongly biased towards the importance of bottom-up interactions rather than food-web interactions and top-down control. However, a less restricted vision should be adopted in studies of bottom-up regulation. The very low epilimnetic concentrations of nutrients documented during the summer months in a few DSL must force research to adopt a different point of view, taking into account potential limitation by P and N (and by other potential limiting factors, e.g. Fe) over shorter, but still ecologically meaningful, time scales relevant for the control of biodiversity patterns Citation276. When the microbial food web accounts for the main part of pelagic carbon flow and most nutrients are regenerated within the system Citation277, a better understanding of resource limitation at short time scales could be obtained by integrating purely chemical determinations with the knowledge of biological structures and activities at the microbial level. In addition, the implications of these aspects for the development of nitrogen-fixing cyanobacteria remain to be investigated. Even more intricate could appear the integration of other environmental stressors such as POPs in the classical bottom-up and top-down models. Nevertheless, taking into account the strong impact that POPs can have on the trophic webs, the impact of these pollutants should not be disregarded in the study of population interactions, community dynamics and risk assessment.
(iii) The difficulty of combining the different bricks to build the cathedral (sensu Peters Citation272), is worsened by the difficulty of integrating the available observations on a supra-regional level. One of the most important tasks of LTER (in different organisational systems, e.g. LTER networks, CIPAIS) is to compare the data obtained in various ecosystems at different temporal and spatial scales, favouring a greater level of integration and a better capacity to generalise and understand the mechanisms acting on continental scales. Nevertheless, the strong disequilibrium existing both in the number and diversity of investigations in the DSL does not allow a complete overall picture to be obtained. The only quantified generalities were defined considering physical, chemical and phytoplankton data collected–or re-calculated from seasonal data–on an annual scale, disregarding the intrinsic complexity existing at shorter time scales. Despite sufficient availability of data from the different lakes, the definition of general patterns in the development of phytoplankton communities on the seasonal scale begun only very recently. Similarly, a number of comparative studies have been made on the distribution of specific and selected POPs in the DSL. However, in this case, the adoption of a regionally integrated approach may be hindered by the large number of micropollutants, which are associated with specific analytical techniques.
(iv) The huge differences in the number of investigations carried out in the DSL has a number of historical reasons linked to the existence of limnological centres or research institutions working in specific lakes, and to the necessity to cope with environmental emergencies. The most common information available in the DSL is that collected within the framework of monitoring programs for the evaluation of the trophic status of water bodies, i.e. the basic limnological information (thermal structure, nutrients and phytoplankton). Complete series of these data exist since at least the 1970s and 1980s for lakes Maggiore and Lugano, and since the beginning of the 1990s for Lake Garda and–though with more scattered surveys–for lakes Iseo and Como. Similarly, the pollution of Lake Maggiore with DDT attracted a number of researchers, with the creation of schools dedicated to the study of different aspects of ecotoxicology, from the evaluation of the diversity and distribution of POPs and metals in the ecosystem compartments to the study of the magnification processes, fate and impact of specific pollutants. At the other extreme, other topics have been completely or insufficiently considered, such as macrophytes and benthonic communities, or have been completely disregarded in the majority of lakes, e.g. paleolimnology and microbial ecology. These topics should not be covered just for the sake of completeness, in the spirit of the multidisciplinary nature of limnology–quite the opposite in fact. Rather than fostering a more ample spectrum of field studies–just because information is lacking–we need to reflect seriously on the criteria that should be used to distinguish relevant scientific research from that lacking concrete aims. Inland waters are more and more threatened by new pressures–from climatic change to the excessive proliferation of toxic microalgae, from the introduction of new species and new micropollutants to the interactions among these new and old stressors. In this changing world, limnology has a great responsibility to produce high quality research and predictive models, which are essential elements for the efficient management of water resources.
Acknowledgements
We are grateful to two anonymous reviewers for their helpful comments and constructive suggestions.
References
- Bini , A , Corbari , D , Falletti , P , Fassina , M , Perotti , CR and Piccin , A . 2007 . Morphology and geological setting of Iseo Lake (Lombardy) through multibeam bathymetry and high-resolution seismic profiles . Swiss J. Geosc. , 100 : 23 – 40 .
- Fanetti , D and Vezzoli , L . 2007 . Sediment input and evolution of lacustrine deltas: the Breggia and Greggio rivers case study (Lake Como, Italy) . Quatern. Internat. , 173 : 113 – 124 .
- Marchetti , F , Fravezzi , L and Büsing , N . 2000 . Nutrient loading from the Sarca River into Lake Garda, Italy . Verh. Int. Verein. Limnol. , 27 : 3129 – 3131 .
- Longinelli , A , Stenni , B , Genoni , L , Flora , O , De Francesco , C and Pellegrini , G . 2008 . A stable isotope study of the Garda lake, Northern Italy: its hydrological balance . J. Hydrol. , 360 : 103 – 116 .
- Giacomelli , P , Rossetti , A and Brambilla , M . 2008 . Adapting water allocation management to drought scenarios . Nat. Hazards Earth Syst. Sci. , 8 : 293 – 302 .
- Giussani , B , Monticelli , D , Gambillara , R , Pozzi , A and Dossi , C . 2008 . Three-way principal component analysis of chemical data from Lake Como watershed . Microchem. J. , 88 : 160 – 166 .
- Galelli , S , Gandolfi , C , Soncini-Sessa , R and Agostani , D . 2010 . Building a metamodel of an irrigation district distributed-parameter model . Agr. Water Manage. , 97 : 187 – 200 .
- Galelli , S and Soncini-Sessa , R . 2010 . Combining metamodelling and stochastic dynamic programming for the design of reservoir release policies . Environ. Modell. Softw. , 25 : 209 – 222 .
- Hilger , S , Sigg , L and Barbieri , A . 1999 . Size fractionation of phosphorus (dissolved, colloidal and particulate) in two tributaries to Lake Lugano . Aquat. Sci. , 61 : 337 – 353 .
- Burlando , P , Montanari , A and Rosso , R . 1996 . Modelling hydrological data with and without long memory . Meccanica , 31 : 87 – 101 .
- Montanari , A , Rosso , R and Taqqu , MS . 1997 . Fractionally differenced ARIMA models applied to hydrologic time series: identification, estimation, and simulation . Water Resour. Res. , 33 : 1035 – 1044 .
- Luino , F , Cirio , CG , Biddoccu , M , Agangi , A , Giulietto , W , Godone , F and Nigrelli , G . 2009 . Application of a model to the evaluation of flood damage . Geoinformatica , 13 : 339 – 353 .
- Ranzi , R , Grossi , G and Bacchi , B . 1999 . Ten years of monitoring areal snowpack in the Southern Alps using NOAA-AVHRR imagery, ground measurements and hydrological data . Hydrol. Process. , 13 : 2079 – 2095 .
- Gilebbi , S , Cavolo , F , Salmaso , N and Cordella , P . 2000 . Underwater light measurements and phytoplankton content in Lake Garda . Verh. Int. Verein. Limnol. , 27 : 2908 – 2912 .
- Salmaso , N . 2005 . Effects of climatic fluctuations and vertical mixing on the interannual trophic variability of Lake Garda, Italy . Limnol. Oceanogr. , 50 : 553 – 565 .
- Maria , L , Biotto , C , Decoene , A and Bonaventura , L . 2009 . A coupled ecological-hydrodynamic model for the spatial distribution of sessile aquatic species in thermally forced basins . Ecol. Model. , 220 : 2310 – 2324 .
- Ambrosetti , W and Barbanti , L . 2005 . Evolution towards meromixis of Lake Iseo (Northern Italy) as revealed by its stability trend . J. Limnol. , 64 : 1 – 11 .
- Buzzi , F , Gerosa , G and Saldavè , G . 1997 . Descrizione e analisi di alcuni aspetti limnologici e idrodinamici del Lago di Como . Docum. Ist. ital. Idrobiol. , 61 : 93 – 115 .
- Morillo , S , Imberger , J , Antenucci , JP and Copetti , D . 2009 . Using impellers to distribute local nutrient loadings in a stratified lake: Lake Como, Italy . J. Hydraul. Eng-ASCE , 135 : 564 – 574 .
- De Cesare , G , Boillat , JL and Schleiss , AJ . 2006 . Circulation in stratified lakes due to flood-induced turbidity currents . J. Environ. Eng.-ASCE , 132 : 1508 – 1517 .
- Aeschbach-Hertig , W , Holzner , CP , Hofer , M , Simona , M , Barbieri , A and Kipfer , R . 2007 . A time series of environmental tracer data from deep, meromictic Lake Lugano, Switzerland . Limnol. Oceanogr. , 52 : 257 – 273 .
- Holzner , CP , Aeschbach-Hertig , W , Simona , M , Veronesi , M , Imboden , DM and Kipfer , R . 2009 . Exceptional mixing events in meromictic Lake Lugano (Switzerland/Italy), studied using environmental tracers . Limnol. Oceanogr. , 54 : 1113 – 1124 .
- Carollo , A . 1995 . La piena del Settembre ed Ottobre 1993 nel bacino idrografico del Lago Maggiore: quadro generale ed analisi dell’episodio del 22–25 Settembre . Docum. Ist. ital. Idrobiol. , 51 : 1 – 33 .
- Barbanti , L , Pompilio , L and Ambrosetti , W . 1995 . La piena dell’autunno 1993 nel Lago Maggiore. Ripercussioni sulle sue caratteristiche fisiche . Docum. Ist. ital. Idrobiol. , 50 : 47
- Ambrosetti , W , Barbanti , L and Sala , N . 2003 . Residence time and physical processes in lakes . J. Limnol. , 62 ( suppl. 1 ) : 1 – 15 .
- Ambrosetti , W , Barbanti , L and Rolla , A . 2006 . Il Clima dell’areale del Lago Maggiore durante gli ultimi cinquant’anni . J. Limnol. , 65 ( suppl. 1 ) : 1 – 62 .
- Ambrosetti , W , Barbanti , L and Carrara , EA . 2010 . Mechanisms of hypolimnion erosion in a deep lake (Lago Maggiore . N. Italy), J. Limnol. , 69 : 3 – 14 .
- Castellano , L , Ambrosetti , W , Barbanti , L and Rolla , A . 2010 . The residence time of the water in Lago Maggiore (N. Italy): first results from an Eulerian-Lagrangian approach . J. Limnol. , 69 : 15 – 28 .
- Ambrosetti , W , Barbanti , L and Pompilio , L . 1996 . Morphometry and thermal stratification in Italian lakes. 3. Dynamics of the deepening of thermocline . Mem Ist. ital. Idrobiol. , 54 : 43 – 50 .
- Barbanti , L , Pompilio , L and Ambrosetti , W . 1996 . Morphometry and thermal stratification in Italian lakes. 2. The ‘Depth Ratio’ as a predictive index of thermal structures . Mem. Ist. ital. Idrobiol. , 54 : 31 – 42 .
- Pompilio , L , Ambrosetti , W and Barbanti , L . 1996 . Morphometry and thermal stratification in Italian lakes. 1. Predictive models . Mem. Ist. ital. Idrobiol. , 54 : 1 – 29 .
- Ambrosetti , W and Barbanti , L . 1997 . Alcune problematiche fisiche dei grandi laghi sudalpini . Docum. Ist. ital. Idrobiol. , 61 : 3 – 18 .
- Ambrosetti , W and Barbanti , L . 1999 . Deep water warming in lakes: an indicator of climatic change . J. Limnol. , 58 : 1 – 9 .
- Ambrosetti , W and Barbanti , L . 2002a . Physical limnology of Italian lakes. 1. Relationship between morphometry and heat content . J. Limnol. , 61 : 147 – 157 .
- Ambrosetti , W and Barbanti , L . 2002b . Physical limnology of Italian lakes. 2. Relationships between morphometric parameters, stability and Birgean work . J. Limnol. , 61 : 159 – 167 .
- Salmaso , N and Decet , F . 1998 . Interactions of physical, chemical and biological processes affecting the seasonality of mineral composition and nutrient cycling in the water column of a deep subalpine lake (Lake Garda, Northern Italy) . Arch. Hydrobiol. , 142 : 385 – 414 .
- Rusconi , R , Azzellino , A , Bellinzona , S , Forte , M , Gallini , R and Sgorbati , G . 2004 . Assessment of drinking water radioactivity content by liquid scintillation counting: set up of high sensitivity and emergency procedures . Anal. Bioanal. Chem. , 379 : 247 – 253 .
- Giussani , B , Dossi , C , Monticelli , D , Pozzi , A and Recchia , S . 2006 . A chemometric approach to the investigation of major and minor ion chemistry in Lake Como (Lombardia, Northern Italy) . Ann. Chim. , 96 : 339 – 346 .
- Hofmann , A and Dominik , J . 1995 . Turbidity and mass concentration of suspended matter in lake water–a comparison of 2 calibration methods . Aquat. Sci. , 57 : 54 – 69 .
- Liu , RM , Hofmann , A , Gulacar , FO , Favarger , PY and Dominik , J . 1996 . Methane concentration profiles in a lake with a permanently anoxic hypolimnion (Lake Lugano, Switzerland-Italy) . Chem. Geol. , 133 : 201 – 209 .
- Bernasconi , M , Barbieri , A and Simona , M . 1997 . Carbon and nitrogen isotope variations in sedimenting organic matter in Lake Lugano . Limnol. Oceanogr. , 42 : 1755 – 1765 .
- Drissner , J , Klemt , E , Klenk , T , Miller , R , Zibold , G , Burger , M and Jakob , A . 1999 . Investigations on the caesium-137 household of Lake Lugano, Switzerland . Czech. J. Phys. , 49 : 133 – 139 .
- Hofmann , A and Filella , M . 1999 . Transport of suspended matter in the hypolimnion of Lake Lugano: a comparison of field observations and model predictions . J. Great Lakes Res. , 25 : 865 – 882 .
- Lienemann , CP , Monnerat , M , Dominik , J and Perret , D . 1999 . Identification of stoichiometric iron-phosphorus colloids produced in a eutrophic lake . Aquat. Sci. , 61 : 133 – 149 .
- Ramisch , F , Dittrich , M , Mattenberger , C , Wehrli , B and Wuest , A . 1999 . Calcite dissolution in two deep eutrophic lakes . Geochim. Cosmochim. Ac. , 63 : 3349 – 3356 .
- Steinmann , P , Billen , T , Loizeau , JL and Dominik , J . 1999 . Beryllium-7 as a tracer to study mechanisms and rates of metal scavenging from lake surface waters . Geochim. Cosmochim. Ac. , 63 : 1621 – 1633 .
- Vengosh , A , Barth , S , Heumann , KG and Eisenhut , S . 1999 . Boron isotopic composition of freshwater lakes from central Europe and possible contamination sources . Acta Hydroch. Hydrob. , 27 : 416 – 421 .
- Perret , D , Gaillard , JF , Dominik , J and Atteia , O . 2000 . The diversity of natural hydrous iron oxides . Environ. Sci. Technol. , 34 : 3540 – 3546 .
- Hofmann , A , Roussy , D and Filella , M . 2002 . Dissolved silica budget in the North basin of Lake Lugano . Chem. Geol. , 182 : 35 – 55 .
- Konoplev , A , Kaminski , S , Klemt , E , Konopleva , I , Miller , R and Zibold , G . 2002 . Comparative study of Cs-137 partitioning between solid and liquid phases in Lakes Constance, Lugano and Vorsee . J. Environ. Radioactiv. , 58 : 1 – 11 .
- Lehmann , MF , Bernasconi , SM , Barbieri , A and McKenzie , JA . 2002 . Preservation of organic matter and alteration of its carbon and nitrogen isotope composition during simulated and in situ early sedimentary diagenesis . Geochim. Cosmochim. Ac. , 66 : 3573 – 3584 .
- Veronesi , ML , Barbieri , A and Hanselmann , KW . 2002 . Phosphorus, carbon and nitrogen enrichment during sedimentation in a seasonally anoxic lake (Lake Lugano, Switzerland) . J. Limnol. , 61 : 215 – 223 .
- Lehmann , MF , Reichert , P , Bernasconi , SM , Barbieri , A and McKenzie , JA . 2003 . Modelling nitrogen and oxygen isotope fractionation during denitrification in a lacustrine redox-transition zone . Geochim. Cosmochim. Ac. , 67 : 2529 – 2542 .
- Lehmann , MF , Bernasconi , SM , Barbieri , A , Simona , M and McKenzie , JA . 2004a . Interannual variation of the isotopic composition of sedimenting organic carbon and nitrogen in Lake Lugano: a long-term sediment trap study . Limnol. Oceanogr. , 49 : 839 – 849 .
- Lehmann , MF , Bernasconi , SM , McKenzie , JA , Barbieri , A , Simona , M and Veronesi , M . 2004b . Seasonal variation of the delta C-13 and delta N-15 of particulate and dissolved carbon and nitrogen in Lake Lugano: constraints on biogeochemical cycling in a eutrophic lake . Limnol. Oceanogr. , 49 : 415 – 429 .
- Bechtel , A and Schubert , CJ . 2009 . Biogeochemistry of particulate organic matter from lakes of different trophic levels in Switzerland . Org. Geochem. , 40 : 441 – 454 .
- Niemann , H , Hitz , C , Blees , J , Schubert , CJ , Veronesi , M , Simona , M and Lehmann , MF . 2009 . Biogeochemical signatures of the anaerobic methane oxidation in a south alpine lake (Lake Lugano) . Geochim. Cosmochim. Ac. , 73 : A942 – A942 .
- Callieri , C . 1997 . Sedimentation and aggregate dynamics in Lake Maggiore, a large, deep lake in Northern Italy . Mem. Ist. ital. Idrobiol. , 56 : 37 – 50 .
- Gröne , T . 1997 . Volatile organic sulfur species in three North Italian lakes: seasonality, possible sources and flux to the atmosphere . Mem. Ist. ital. Idrobiol. , 56 : 77 – 94 .
- Baudo , R , Cenci , RM , Sena , F , Dabergami , D , Leva , F , Trincherini , PR , Beltrami , M , Barbero , P , Corace , C and Murray , CN . 2002 . Chemical composition of Lake Maggiore sediments . Fresen. Environ. Bull. , 11 : 675 – 680 .
- Putyrskaya , V and Klemt , E . 2007 . Modeling Cs-137 migration processes in lake sediments . J. Environ. Radioactiv. , 96 : 54 – 62 .
- Putyrskaya , V , Klemt , E and Röllin , S . 2009 . Migration of Cs-137 in tributaries, lake water and sediment of Lago Maggiore (Italy, Switzerland)–analysis and comparison with Lago di Lugano and other lakes . J. Environ. Radioactiv. , 100 : 35 – 48 .
- Salmaso , N , Decet , F and Mosello , R . 1997a . Chemical characteristics and trophic evolution of the deep subalpine Lake Garda (Northern Italy) . Mem. Ist. ital. Idrobiol. , 56 : 51 – 76 .
- Salmaso , N , Decet , F , Manfrin , M and Cordella , P . 1997b . Ricerche limnologiche sul Lago di Garda (1991–1996) . Docum. Ist. ital. Idrobiol. , 61 : 173 – 199 .
- Salmaso , N , Decet , F and Cordella , P . 1999 . Understanding deep oligotrophic subalpine lakes for efficient management . Hydrobiologia, , 395 : 253 – 263 .
- Salmaso , N , Decet , F and Cordella , P . 2002 . Spring mixing depth affects the interannual variations in phytoplankton abundance and composition in deep lakes. A case study from Lake Garda (Northern Italy) . Verh. Int. Verein. Limnol. , 28 : 1486 – 1489 .
- Garibaldi , L , Brizzio , MC , Mezzanotte , V , Varallo , A and Mosello , R . 1995 . The continuing evolution of Lago Iseo (N. Italy): the appearance of anoxia . Mem. Ist. ital. Idrobiol. , 53 : 191 – 212 .
- Garibaldi , L , Brizzio , MC , Mezzanotte , V , Varallo , A and Mosello , R . 1997 . Evoluzione idrochimica e trofica del Lago d’Iseo . Docum. Ist. ital. Idrobiol. , 61 : 135 – 151 .
- Garibaldi , L , Mezzanotte , V , Brizzio , MC , Rogora , M and Mosello , R . 1999 . The trophic evolution of Lake Iseo as related to its holomixis . J. Limnol. , 58 : 10 – 19 .
- Barbieri , A and Simona , M . 1997 . Evoluzione trofica recente del Lago di Lugano in relazione agli interventi di risanamento . Docum. Ist. ital. Idrobiol. , 61 : 73 – 91 .
- Calderoni , A and Mosello , R . 1996 . L’eutrofizzazione del Lago Maggiore e il suo risanamento . Docum. Ist. ital. Idrobiol. , 56 : 5 – 20 .
- Calderoni , A , Mosello , R and de Bernardi , R . 1997 . Interpretazioni problematiche di alcuni processi nella fase di oligotrofizzazione del Lago Maggiore nel decennio 1988–1997 . Docum. Ist. ital. Idrobiol. , 61 : 33 – 53 .
- Mosello , R , Barbieri , A , Brizzio , MC , Calderoni , A , Marchetto , A , Passera , S , Rogora , M and Tartari , G . 2001 . Nitrogen budget of Lago Maggiore: the relative importance of atmospheric deposition and catchment sources . J. Limnol. , 60 : 27 – 40 .
- Rogora , M , Mosello , R , Calderoni , A and Barbieri , A . 2006 . Nitrogen budget of a subalpine lake in northwestern Italy: the role of atmospheric input in the upward trend of nitrogen concentrations . Verh. Int. Verein. Limnol. , 29 : 2027 – 2030 .
- de Bernardi , R , Calderoni , A and Mosello , R . 1996 . Environmental problems in Italian lakes, and lakes Maggiore and Orta as successful examples of correct management leading to restoration . Verh. Int. Verein. Limnol. , 26 : 123 – 138 .
- Mosello , R , Calderoni , A and de Bernardi , R . 1997 . Le indagini sulla evoluzione dei laghi profondi sudalpini svolte dal C.N.R. Istituto Italiano di Idrobiologia . Docum. Ist. ital. Idrobiol. , 61 : 19 – 32 .
- Tartari , G , Copetti , D and Marchetto , A . 2002 . Northern Italian lakes: regionalization of limnological features and pressure factors relationships . Verh. Int. Verein. Limnol. , 28 : 223 – 227 .
- Salmaso , N , Mosello , R , Garibaldi , L , Decet , F , Brizzio , MC and Cordella , P . 2003a . Vertical mixing as a determinant of trophic status in deep lakes: a case study from two lakes south of the Alps (Lake Garda and Lake Iseo) . J. Limnol. , 62 : 33 – 41 .
- Salmaso , N , Morabito , G , Garibaldi , L and Mosello , R . 2007 . Trophic development of the deep lakes south of the Alps: a comparative analysis . Fund. Appl. Limnol. , 170 : 177 – 196 .
- Salmaso , N . 1996 . Seasonal variation in the composition and rate of change of the phytoplankton community in a deep subalpine lake (Lake Garda, northern Italy). An application of nonmetric multidimensional scaling and cluster analysis . Hydrobiologia , 337 : 49 – 68 .
- Büsing , N . 1998 . Seasonality of phytoplankton as an indicator of trophic status of the large perialpine Lago di Garda . Hydrobiologia , 370 : 153 – 162 .
- Salmaso , N . 2000 . Factors affecting the seasonality and distribution of cyanobacteria and chlorophytes: a case study from the large lakes south of the Alps, with special reference to Lake Garda . Hydrobiologia, , 438 : 43 – 63 .
- Salmaso , N . 2002 . Ecological patterns of phytoplankton assemblages in Lake Garda: seasonal, spatial and historical features . J. Limnol. , 61 : 95 – 115 .
- Salmaso , N . 2003 . Life strategies, dominance patterns and mechanisms promoting species coexistence in phytoplankton communities along complex environmental gradients . Hydrobiologia , 502 : 13 – 36 .
- Salmaso , N and Padisák , J . 2007 . Morpho-functional groups and phytoplankton development in two deep lakes (Lake Garda, Italy and Lake Stechlin, Germany) . Hydrobiologia , 578 : 97 – 112 .
- Salmaso , N . Long-term phytoplankton community changes in a deep subalpine lake: responses to nutrient availability and climatic fluctuations, Freshwater Biol. 55 (2010), pp. 825–846
- Garibaldi , L , Anzani , A , Marieni , A , Leoni , B and Mosello , R . 2003 . Studies on the phytoplankton of the deep subalpine Lake Iseo . J. Limnol. , 62 : 177 – 189 .
- Bettinetti , R , Morabito , G and Provini , A . 2000 . Phytoplankton assemblage structure and dynamics as indicator of the recent trophic and biological evolution of the western basin of Lake Como (N. Italy) . Hydrobiologia , 435 : 177 – 190 .
- Buzzi , F . 2002 . Phytoplankton assemblages in two sub-basins of Lake Como . J. Limnol. , 61 : 117 – 128 .
- Scheffler , W and Morabito , G . 2003 . Topical observations on centric diatoms (Bacillariophyceae, Centrales) of Lake Como (N. Italy) . J. Limnol. , 62 : 47 – 60 .
- Scheffler , W , Nicklisch , A and Schonfelder , I . 2005 . Articles on the morphology, ecology and ontogenesis of the planktonic diatom Cyclotella comensis (Grunow). Studies on the historic and recently sampled material . Diatom Res. , 20 : 171 – 200 .
- Simona , M . 2003 . Winter and spring mixing depths affect the trophic status and composition of phytoplankton in the northern meromictic basin of Lake Lugano . J. Limnol. , 62 : 190 – 206 .
- Ruggiu , D , Morabito , G , Panzani , P and Pugnetti , A . 1998 . Trends and relations among basic phytoplankton characteristics in the course of the long-term oligotrophication of Lake Maggiore (Italy) . Hydrobiologia , 370 : 243 – 257 .
- Morabito , G and Pugnetti , A . 2000 . Primary productivity and related variables in the course of the trophic evolution of Lake Maggiore . Verh. Int. Verein. Limnol. , 27 : 2934 – 2937 .
- Marchetto , A and Musazzi , S . 2001 . Comparison between sedimentary and living diatoms in Lago Maggiore (N. Italy): implications of using transfer functions . J. Limnol. , 60 : 19 – 26 .
- Morabito , G , Ruggiu , D and Panzani , P . 2002 . Recent dynamics (1995–1999) of the phytoplankton assemblages in Lago Maggiore as a basic tool for defining association patterns in the Italian deep lakes . J. Limnol. , 61 : 129 – 145 .
- Morabito , G , Oggioni , A and Panzani , P . 2003 . Phytoplankton assemblage at equilibrium in large and deep subalpine lakes: a case study from Lago Maggiore (N. Italy) . Hydrobiologia , 502 : 37 – 48 .
- Morabito , G , Hamza , W and Ruggiu , D . 2004 . Carbon assimilation and phytoplankton growth rates across the trophic spectrum: an application of the chlorophyll labelling technique . J. Limnol. , 63 : 33 – 43 .
- Morabito , G , Oggioni , A and Caravati , E . 2005 . Decadal trends of pelagic algal biomass capacities in Lago Maggiore (N. Italy) . Verh. Int. Verein. Limnol. , 29 : 231 – 234 .
- Callieri , C , Caravati , E , Morabito , G and Oggioni , A . 2006 . The unicellular freshwater cyanobacterium Synechococcus and mixotrophic flagellates: evidence for a functional association in an oligotrophic, subalpine lake . Freshwater Biol. , 51 : 263 – 273 .
- Pugnetti , A , Camatti , E , Mangoni , O , Morabito , G , Oggioni , A and Saggiomo , V . 2006 . Phytoplankton production in Italian freshwater and marine ecosystems: state of the art and perspectives . Chem. Ecol. , 22 : S49 – S69 .
- Kamenir , Y and Morabito , G . 2009 . Lago Maggiore oligotrophication as seen from the long-term evolution of its phytoplankton taxonomic size structure . J. Limnol. , 68 : 146 – 161 .
- Salmaso , N , Morabito , G , Mosello , R , Garibaldi , L , Simona , M , Buzzi , F and Ruggiu , D . 2003b . A synoptic study of phytoplankton in the deep lakes south of the Alps (lakes Garda, Iseo, Como, Lugano and Maggiore . J. Limnol. , 62 : 207 – 227 .
- Salmaso , N , Morabito , G , Buzzi , F , Garibaldi , L , Simona , M and Mosello , R . 2006 . Phytoplankton as an indicator of the water quality of the deep lakes south of the Alps . Hydrobiologia , 563 : 167 – 187 .
- Salmaso , N and Naselli-Flores , L . 1999 . Studies on the zooplankton of the deep subalpine Lake Garda . J. Limnol. , 58 : 66 – 76 .
- Pucciarelli , S , Buonanno , F , Pellegrini , G , Pozzi , S , Ballarini , P and Miceli , C . 2008 . Biomonitoring of Lake Garda: identification of ciliate species and symbiotic algae responsible for the ‘black-spot’ bloom during the summer of 2004 . Environ. Res. , 107 : 194 – 200 .
- de Bernardi , R and Canale , C . 1995 . Ricerche pluriennali (1948–1992) sull’ecologia dello zooplancton del Lago Maggiore . Docum. Ist. ital. Idrobiol. , 55 : 68
- Manca , M and Comoli , P . 1995 . Seasonal-changes in size of the feeding basket of Leptodora kindtii (Focke) in Lago Maggiore as related to variations in prey size selection . Limnol. Oceanogr. , 40 : 834 – 838 .
- Manca , M , Beltrami , M and Sonvico , D . 1996 . On the appearance of epibionts on the crustacean zooplankton of a large subalpine lake undergoing oligotrophication (L. Maggiore . Italy), Mem. Ist. ital. Idrobiol. , 54 : 161 – 171 .
- Manca , M and Sonvico , D . 1996 . Seasonal variations in population density and size structure of Conochilus in Lago Maggiore: a biannual study . Mem. Ist. ital. Idrobiol. , 54 : 97 – 108 .
- Manca , M , Comoli , P and Spagnuolo , T . 1997 . Length-specific carbon content of the Daphnia population in a large subalpine lake, Lago Maggiore (Northern Italy): the importance of seasonality . Aquat. Sci. , 59 : 48 – 56 .
- Manca , M and Ruggiu , D . 1998 . Consequences of pelagic food-web changes during a long-term lake oligotrophication process . Limnol. Oceanogr. , 43 : 1368 – 1373 .
- Riccardi , N and Mangoni , M . 1999 . Considerations on the biochemical composition of some freshwater zooplankton species . J. Limnol. , 58 : 58 – 65 .
- Manca , M and Comoli , P . 2000 . Biomass estimates of freshwater zooplankton from length-carbon regression equations . J. Limnol. , 59 : 15 – 18 .
- Manca , M , Cavicchioni , N and Morabito , G . 2000a . First observations on the effect of a complete, exceptional overturn of Lake Maggiore on plankton and primary productivity . Int. Rev. Hydrobiol. , 85 : 209 – 222 .
- Manca , M , Ramoni , C and Comoli , P . 2000b . The decline of Daphnia hyalina galeata in Lago Maggiore: a comparison of the population dynamics before and after oligotrophication . Aquat. Sci. , 62 : 142 – 153 .
- Riccardi , N . 2000 . Comparison of different stoichiometric methods for the estimation of proximate biochemical composition of crustacean zooplankton and some considerations on energy transfer to planktophagous fish . J. Limnol. , 59 : 179 – 185 .
- Manca , M , Carnovale , A and Alemani , P . 2004 . Exotopic protrusions and ellobiopsid infection in zooplanktonic copepods of a large, deep subalpine lake, Lago Maggiore, in northern Italy . J. Plankton Res. , 26 : 1257 – 1263 .
- Manca , M , Callieri , C and Cattaneo , A . 2007a . Daphnia and ciliates: who is the prey? . J. Limnol. , 66 : 170 – 173 .
- Manca , M , Portogallo , M and Brown , ME . 2007b . Shifts in phenology of Bythotrephes longimanus and its modern success in Lake Maggiore as a result of changes in climate and trophy . J. Plankton Res. , 29 : 515 – 525 .
- Riccardi , N and Rossetti , G . 2007 . Eudiaptomus gracilis in Italy: how, where and why . J. Limnol. , 66 : 64 – 69 .
- Manca , M , Vijverberg , J , Polishchuk , LV and Voronov , DA . 2008 . Daphnia body size and population dynamics under predation by invertebrate and fish predators in Lago Maggiore: an approach based on contribution analysis . J. Limnol. , 67 : 15 – 21 .
- Visconti , A , Manca , M and de Bernardi , R . 2008 . Eutrophication-like response to climate warming: an analysis of Lago Maggiore (N. Italy) zooplankton in contrasting years . J. Limnol. , 67 : 87 – 92 .
- Manca , M and DeMott , WR . 2009 . Response of the invertebrate predator Bythotrephes to a climate-linked increase in the duration of a refuge from fish predation . Limnol. Oceanogr. , 54 : 2506 – 2512 .
- Lazzaretti-Ulmer , MA and Hanselmann , KW . 1999 . Seasonal variation of the microbially regulated buffering capacity at sediment-water interfaces in a freshwater lake . Aquat. Sci. , 61 : 59 – 74 .
- Callieri , C , Bertoni , R , Amicucci , E and Pinolini , ML . 1995 . Picoplankton composition, size frequency distribution and carbon content in Lago Maggiore, Italy . Mem. Ist. ital. Idrobiol. , 53 : 177 – 179 .
- Callieri , C and Pinolini , ML . 1995 . Picoplankton In Lake Maggiore, Italy . Int. Rev. Ges. Hydrobio. , 80 : 491 – 501 .
- Callieri , C , Amicucci , E , Bertoni , R and Voros , L . 1996 . Fluorometric characterization of two picocyanobacteria strains from lakes of different underwater light quality . Int. Rev. Ges. Hydrobio. , 81 : 13 – 23 .
- Bertoni , R , Callieri , C , Morabito , G , Pinolini , ML and Pugnetti , A . 1997 . Quali-quantitative changes in organic carbon production during the oligotrophication of Lake Maggiore . Italy, Verh. Int. Verein. Limnol. , 26 : 300 – 304 .
- Bertoni , R and Callieri , C . 1999 . Effects of UVB radiation on freshwater autotrophic and heterotrophic picoplankton in a subalpine lake . J. Plankton Res. , 21 : 1373 – 1388 .
- Bertoni , R . 2000 . Size distribution of particulate organic carbon in Lago Maggiore . Verh. Int. Verein. Limnol. , 27 : 2836 – 2839 .
- Callieri , C . 2000 . Carbon flux from picocyanobacteria to ciliates in a subalpine lake . Verh. Int. Verein. Limnol. , 27 : 2840 – 2844 .
- Callieri , C and Stockner , J . 2000 . Picocyanobacteria success in oligotrophic lakes: fact or fiction? . J. Limnol. , 59 : 72 – 76 .
- Passoni , S and Callieri , C . 2000 . Picocyanobacteria single forms, aggregates and microcolonies: survival strategy or species succession? . Verh. Int. Verein. Limnol. , 27 : 1879 – 1883 .
- Callieri , C and Piscia , R . 2002 . Photosynthetic efficiency and seasonality of autotrophic picoplankton in Lago Maggiore after its recovery . Freshwater Biol. , 47 : 941 – 956 .
- Callieri , C , Karjalainen , SM and Passoni , S . 2002 . Grazing by ciliates and heterotrophic nanoflagellates on picocyanobacteria in Lago Maggiore . Italy, J. Plankton Res. , 24 : 785 – 796 .
- Bertoni , R , Piscia , R and Callieri , C . 2004 . Horizontal heterogeneity of seston, organic carbon and picoplankton in the photic zone of Lago Maggiore, Northern Italy . J. Limnol. , 63 : 244 – 249 .
- Dittrich , M and Sibler , S . 2005 . Cell surface groups of two picocyanobacteria strains studied by zeta potential investigations, potentiometric titration, and infrared spectroscopy . J. Colloid Interf. Sci. , 286 : 487 – 495 .
- Callieri , C , Corno , G , Caravati , E , Galafassi , S , Bottinell , M and Bertoni , R . 2007 . Photosynthetic characteristics and diversity of freshwater Synechococcus at two depths during different mixing conditions in a deep oligotrophic lake . J. Limnol. , 66 : 81 – 89 .
- Bertoni , R , Callieri , C , Balseiro , E and Modenutti , B . 2008 . Susceptibility of bacterioplankton to nutrient enrichment of oligotrophic and ultraoligotrophic lake waters . J. Limnol. , 67 : 120 – 127 .
- Bertoni , R and Callieri , C . 1997 . Il microbial loop e il carbonio organico nei grandi laghi sudalpini . Docum. Ist. ital. Idrobiol. , 61 : 201 – 224 .
- Callieri , C and Heinimaa , S . 1997 . Microbial loop in the large subalpine lakes . Mem. Ist. ital. Idrobiol. , 56 : 143 – 156 .
- Balsamo , M and Fregni , E . 1995 . Gastrotrichs from interstitial fresh-water, with a description of 4 new species . Hydrobiologia , 302 : 163 – 175 .
- Casellato , S , La Piana , G , Latella , L and Ruffo , S . 2006 . Dikerogammarus villosus (Sowinsky, 1894) (Crustacea, Amphipoda, Gammaridae) for the first time in Italy . Ital. J. Zool. , 73 : 97 – 104 .
- Quaglia , F , Lattuada , L , Mantecca , P and Bacchetta , R . 2008 . Zebra mussels in Italy: where do they come from? . Biol. Invasions , 10 : 555 – 560 .
- Ciutti , F and Cappelletti , C . 2009 . First record of Corbicula fluminalis (Muller, 1774) in Lake Garda (Italy), living in sympatry with Corbicula fluminea (Muller, 1774) . J. Limnol. , 68 : 162 – 165 .
- Mantecca , P , Vailati , G , Garibaldi , L and Bacchetta , R . 2003a . Depth effects on zebra mussel reproduction . Malacologia , 45 : 109 – 120 .
- Binelli , A , Provini , A and Galassi , S . 1997 . Modifications in Lake Como (N. Italy) caused by the zebra mussel (Dreissena polymorpha) . Water Air Soil Poll. , 99 : 633 – 640 .
- Bacchetta , R , Mantecca , P and Vailati , G . 2001 . Reproductive behavior of the freshwater mussel Dreissena polymorpha in Italy: a comparison between two populations . Arch. Hydrobiol. , 151 : 247 – 262 .
- Vailati , G , Bacchetta , R and Mantecca , P . 2001 . Sexual maturity in young Dreissena polymorpha from Lake Como (N. Italy) . Invertebr. Reprod. Dev. , 40 : 181 – 192 .
- Campanella , L , Gatta , T and Ravera , O . 2005 . Relationship between anti-oxidant capacity and manganese accumulation in the soft tissues of two freshwater molluscs: Unio pictorum mancus (Lamellibranchia, Unionidae) and Viviparus ater (Gastropoda, Prosobranchia) . J. Limnol. , 64 : 153 – 158 .
- Ravera , O , Frediani , A and Riccardi , N . 2007 . Seasonal variations in population dynamics and biomass of two Unio pictorum mancus (Mollusca, Unionidae) populations from two lakes of different trophic state . J. Limnol. , 66 : 15 – 27 .
- Giuffra , E , Guyomard , R and Forneris , G . 1996 . Phylogenetic relationships and introgression patterns between incipient parapatric species of Italian brown trout (Salmo trutta L. complex) . Mol. Ecol. , 5 : 207 – 220 .
- Heldstab , H and Katoh , M . 1995 . Low genetic-variation in perch (Perca fluviatilis L.) from 3 major european drainage systems in switzerland . Aquat. Sci. , 57 : 14 – 19 .
- Tatrai , I , Giussani , G , Manca , M and de Bernardi , R . 1995 . An experimental study of Lago Maggiore zooplankton consumption by bleak (Alburnus alburnus alborella) in different simulated habitats . Mem. Ist. ital. Idrobiol. , 53 : 74 – 84 .
- Calderoni , A . 1996 . Attività della Commissione per la pesca nelle acque italo-svizzere per la gestione della fauna ittica del Lago Maggiore . Docum. Ist. ital. Idrobiol. , 56 : 65 – 87 .
- Grimaldi , E . 1996 . Realtà e potenzialità delle attività di pesca nel Lago Maggiore . Docum. Ist. ital. Idrobiol. , 56 : 21 – 30 .
- Serrini , G , Luzzana , U , Grimaldi , P , Paleari , MA and Valfre , F . 1996 . Fatty acid composition of some coregonid species (Coregonus spp.) from different European lakes . J. Appl. Ichthyol. , 12 : 57 – 59 .
- Douglas , MR and Brunner , PC . 2002 . Biodiversity of Central Alpine Coregonus (Salmoniformes): impact of one-hundred years of management . Ecol. Appl. , 12 : 154 – 172 .
- Volta , P and Jepsen , N . 2008 . The recent invasion of Rutilus rutilus (L.) (Pisces: Cyprinidae) in a large South-Alpine lake: Lago Maggiore . J. Limnol. , 67 : 163 – 170 .
- Luzzana , U , Serrini , G , Moretti , VM , Grimaldi , P , Paleari , MA and Valfre , F . 1996 . Seasonal variations in fat content and fatty acid composition of male and female coregonid ‘bondella’ from Lake Maggiore and landlocked shad from Lake Come (Northern Italy) . J. Fish Biol. , 48 : 352 – 366 .
- Gagliardi , A , Martinoli , A , Wauters , L and Tosi , G . 2003 . A floating platform: a solution to collecting pellets when cormorants roost over water . Waterbirds , 26 : 54 – 55 .
- Martinoli , A , Gagliardi , A , Preatoni , DG , di Martino , S , Wauters , LA and Tosi , G . 2003 . The extent of Great Crested Grebe predation on bleak in Lake Como, Italy . Waterbirds , 26 : 201 – 208 .
- Gagliardi , A , Martinoli , A , Preatoni , D , Wauters , LA and Tosi , G . 2006 . Behavioral responses of wintering Great Crested Grebes to dissuasion experiments: implications for management . Waterbirds , 29 : 105 – 114 .
- Sergio , F and Boto , A . 1999 . Nest dispersion, diet, and breeding success of Black Kites (Milvus migrans) in the Italian pre-Alps . J. Raptor Res. , 33 : 207 – 217 .
- Sergio , F , Pedrini , P and Marchesi , L . 2003 . Spatio-temporal shifts in gradients of habitat quality for an opportunistic avian predator . Ecography , 26 : 243 – 255 .
- Lleo , MD , Tafi , MC , Signoretto , C , Dal Cero , C and Canepari , P . 1999 . Competitive polymerase chain reaction for quantification of nonculturable Enterococcus faecalis cells in lake water . FEMS Microbiol. Ecol. , 30 : 345 – 353 .
- Signoretto , C , Burlacchini , G , Lleo , MD , Pruzzo , C , Zampini , M , Pane , L , Franzini , G and Canepari , P . 2004 . Adhesion of Enterococcus faecalis in the nonculturable state to plankton is the main mechanism responsible for persistence of this bacterium in both lake and seawater . Appl. Environ. Microb. , 70 : 6892 – 6896 .
- Terramocci , R , Pagani , L , Brunati , P , Gatti , S , Bernuzzi , AM and Scaglia , M . 2001 . Reappearance of human diphyllobothriasis in a limited area of Lake Como, Italy . Infection , 29 : 93 – 95 .
- Wicht , B , Gustinelli , A , Fioravanti , ML , Invernizzi , S and Peduzzi , R . 2009a . Prevalence of the broad tapeworm Diphyllobothrium latum in perch (Perca fluviatilis) and analysis of abiotic factors influencing its occurrence in Lake Lario (Como, Italy) . Bull. Eur. Assn. Fish P. , 29 : 58 – 65 .
- Peduzzi , R and Boucher-Rodoni , R . 2001 . Resurgence of human bothriocephalosis (Diphyllobothrium latum) in the subalpine lake region . J. Limnol. , 60 : 41 – 44 .
- Wicht , B , Limoni , C , Peduzzi , R and Petrini , O . 2009b . Diphyllobothrium latum (Cestoda: Diphyllobothriidea) in perch (Perca fluviatilis) in three sub-alpine lakes: influence of biotic and abiotic factors on prevalence . J. Limnol. , 68 : 167 – 173 .
- Binelli , A and Provini , A . 2003a . The PCB pollution of Lake Iseo (N. Italy) and the role of biomagnification in the pelagic food web . Chemosphere , 53 : 143 – 151 .
- Binelli , A and Provini , A . 2004 . Risk for human health of some POPs due to fish from Lake Iseo . Ecotox. Environ. Safe. , 58 : 139 – 145 .
- Bacchetta , R and Mantecca , P . 2009 . DDT polluted meltwater affects reproduction in the mussel Dreissena polimorpha . Chemosphere , 76 : 1380 – 1385 .
- Provini , A , Binelli , A and Galassi , S . 1997 . Ruolo di Dreissena polymorpha nel ciclo dei nutrienti e dei PCB nel bacino occidentale del Lago di Como . Docum. Ist. ital. Idrobiol. , 61 : 117 – 133 .
- Binelli , A , Galassi , S and Provini , A . 2001a . Factors affecting the use of Dreissena polymorpha as a bioindicator: the PCB pollution in Lake Como (N. Italy) . Water Air Soil Poll. , 125 : 19 – 32 .
- Guzzella , L , Feretti , D and Monarca , S . 2002 . Advanced oxidation and adsorption technologies for organic micropollutant removal from lake water used as drinking-water supply . Water Res. , 36 : 4307 – 4318 .
- Ciceri , E , Recchia , S , Dossi , C , Yang , L and Sturgeon , RR . 2008 . Validation of an isotope dilution, ICP-MS method based on internal mass bias correction for the determination of trace concentrations of Hg in sediment cores . Talanta , 74 : 642 – 647 .
- Kupper , J , Baurngartner , M , Bacciarini , LN , Hoop , R , Kupferschmidt , H and Naegeli , H . 2007 . Carbofuran poisoning in mallard ducks . Schweiz. Arch. Tierh. , 149 : 517 – 520 .
- Cenci , R , Baudo , R , Bianchi , M and Muntau , H . 1996 . Mercury deposition and redistribution patterns (southern Lake Maggiore . Italy), Environ. Technol. , 17 : 547 – 551 .
- Guzzella , L , Patrolecco , L , Pagnotta , R , Langone , L and Guilizzoni , P . 1998 . DDT and other organochlorine compounds in the Lake Maggiore sediments: a recent point source of contamination . Fresen. Environ. Bull. , 7 : 79 – 89 .
- Binelli , A , Bacchetta , R , Vailati , G , Galassi , S and Provini , A . 2001b . DDT contamination in Lake Maggiore (N. Italy) and effects on zebra mussel spawning . Chemosphere , 45 : 409 – 415 .
- Silvana , G and Roberto , C . 2001 . Key species for monitoring persistent and bioaccumulable pesticides . Fresen. Environ. Bull. , 10 : 451 – 454 .
- Galassi , S , Saino , N , Melone , G and Croce , V . 2002 . DDT homologues and PCBs in eggs of great crested grebe (Podiceps cristatus) and mallard (Anas platyrhynchos) from Lake Maggiore (Italy) . Ecotox. Environ. Safe. , 53 : 163 – 169 .
- Binelli , A and Provini , A . 2003b . DDT is still a problem in developed countries: the heavy pollution of Lake Maggiore . Chemosphere , 52 : 717 – 723 .
- Mantecca , P , Vailati , G and Bacchetta , R . 2003b . Histological studies on the zebra mussel Dreissena polymorpha reproduction from a DDT contaminated area in Lake Maggiore (N. Italy) . Arch. Hydrobiol. , 158 : 233 – 248 .
- Ravera , O , Beone , GM , Cenci , R and Lodigiani , P . 2003a . Metal concentrations in Unio pictorum mancus (Mollusca, Lamellibranchia) from of 12 Northern Italian lakes in relation to their trophic level . J. Limnol. , 62 : 121 – 138 .
- Ravera , O , Cenci , R , Beone , GM , Dantas , M and Lodigiani , P . 2003b . Trace element concentrations in freshwater mussels and macrophytes as related to those in their environment . J. Limnol. , 62 : 61 – 70 .
- Binelli , A , Bacchetta , R , Mantecca , P , Ricciardi , F , Provini , A and Vailati , G . 2004a . DDT in zebra mussels from Lake Maggiore (N. Italy): level of contamination and endocrine disruptions . Aquat. Toxicol. , 69 : 175 – 188 .
- Binelli , A , Ricciardi , F and Provini , A . 2004b . Present status of POP contamination in Lake Maggiore (Italy) . Chemosphere , 57 : 27 – 34 .
- Bettinetti , R , Croce , V and Galassi , S . 2005 . Ecological risk assessment for the recent case of DDT pollution in Lake Maggiore (Northern Italy) . Water Air Soil Poll. , 162 : 385 – 399 .
- Olivella , MA . 2005 . Trace analysis of polycyclic aromatic hydrocarbons in suspended particulate matter by accelerated solvent extraction followed by gas chromatography-mass spectrometry . Anal. Bioanal. Chem. , 383 : 107 – 114 .
- Bettinetti , R , Croce , V , Galassi , S and Volta , P . 2006 . pp’DDT and pp’DDE accumulation in a food chain of Lake Maggiore (Northern Italy): testing steady-state condition . Environ. Sci. Pollut. R. , 13 : 59 – 66 .
- Di Guardo , A , Ferrari , C and Infantino , A . 2006 . Development of a dynamic aquatic model (DynA Model): estimating temporal emissions of DDT to Lake Maggiore (N. Italy) . Environ. Sci. Pollut. R. , 13 : 50 – 58 .
- Galassi , S , Volta , P , Calderoni , A and Guzzella , L . 2006 . Cycling pp’DDT and pp’DDE at a watershed scale: the case of Lago Maggiore (Italy) . J. Limnol. , 65 : 100 – 106 .
- Olivella , MA . 2006a . Polycyclic aromatic hydrocarbons in rainwater and surface waters of Lake Maggiore, a subalpine lake in Northern Italy . Chemosphere , 63 : 116 – 131 .
- Olivella , MA . 2006b . Isolation and analysis of polycyclic aromatic hydrocarbons from natural water using accelerated solvent extraction followed by gas chromatography-mass spectrometry . Talanta , 69 : 267 – 275 .
- Ricciardi , F , Binelli , A and Provini , A . 2006 . Use of two biomarkers (CYP450 and acetylcholinesterase) in zebra mussel for the biomonitoring of Lake Maggiore (northern Italy) . Ecotox. Environ. Safe. , 63 : 406 – 412 .
- Binelli , A , Riva , C and Provini , A . 2007 . Biomarkers in Zebra mussel for monitoring and quality assessment of Lake Maggiore (Italy) . Biomarkers , 12 : 349 – 368 .
- Loos , R , Wollgast , J , Huber , T and Hanke , G . 2007 . Polar herbicides, pharmaceutical products, perfluorooctanesulfonate (PFOS), perfluorooctanoate (PFOA), and nonylphenol and its carboxylates and ethoxylates in surface and tap waters around Lake Maggiore in Northern Italy . Anal. Bioanal. Chem. , 387 : 1469 – 1478 .
- Ravera , O , Beone , GM , Trincherini , PR and Riccardi , N . 2007 . Seasonal variations in metal content of two Unio pictorum mancus (Mollusca, Unionidae) populations from two lakes of different trophic state . J. Limnol. , 66 : 28 – 39 .
- Vives , I , Canuti , E , Castro-Jimenez , J , Christoph , EH , Eisenreich , SJ , Hanke , G , Huber , T , Mariani , G , Mueller , A , Skejo , H , Umlauf , G and Wollgast , J . 2007 . Occurrence of polychlorinated dibenzo-p-dioxins and dibenzofurans (PCDD/Fs), polychlorinated biphenyls (PCBs) and polybrominated diphenyl ethers (PBDEs) in Lake Maggiore (Italy and Switzerland) . J. Environ. Monitor. , 9 : 589 – 598 .
- Binelli , A , Guzzella , L and Roscioli , C . 2008 . Levels and congener profiles of polybrominated diphenyl ethers (PBDEs) in Zebra mussels (D. polymorpha) from Lake Maggiore (Italy) . Environ. Pollut. , 153 : 610 – 617 .
- Castro-Jimenez , J , Mariani , G , Eisenreich , SJ , Christoph , EH , Hanke , G , Canuti , E , Skejo , H and Umlauf , G . 2008 . Atmospheric input of POPs into Lake Maggiore (Northern Italy): PCDD/F and dioxin-like PCB profiles and fluxes in the atmosphere and aquatic system . Chemosphere , 73 : S122 – S130 .
- Guzzella , L , Roscioli , C and Binelli , A . 2008 . Contamination by polybrominated diphenyl ethers of sediments from the Lake Maggiore basin (Italy and Switzerland) . Chemosphere , 73 : 1684 – 1691 .
- Mariani , G , Canuti , E , Castro-Jimenez , J , Christoph , EH , Eisenreich , SJ , Hanke , G , Skejo , H and Umlauf , G . 2008 . Atmospheric input of POPs into Lake Maggiore (Northern Italy): PBDE concentrations and profile in air, precipitation, settling material and sediments . Chemosphere , 73 : S114 – S121 .
- Dueri , S , Castro-Jimenez , J and Zaldivar , JM . 2009 . Modelling the influence of thermal stratification and complete mixing on the distribution and fluxes of polychlorinated biphenyls in the water column of Ispra Bay (Lake Maggiore) . Chemosphere , 75 : 1266 – 1272 .
- Ravera , O , Beone , GM , Fontanella , MC , Riccardi , N and Cattani , I . 2009 . Comparison between the mercury contamination in populations of Unio pictorum mancus (Mollusca, Bivalvia) from two lakes of different trophic state: the oligo-mesotrophic Lake Maggiore and the eutrophic Lake Candia . J. Limnol. , 68 : 359 – 367 .
- Volta , P , Tremolada , P , Neri , MC , Giussani , G and Galassi , S . 2009 . Age-Dependent Bioaccumulation of Organochlorine Compounds in Fish and their Selective Biotransformation in Top Predators from Lake Maggiore (Italy) . Water Air Soil Poll. , 197 : 193 – 209 .
- Provini , A , Galassi , S , Guzzella , L and Valli , G . 1995 . PCB profiles in sediments of lakes Maggiore, Como and Garda (Italy) . Mar. Freshwater Res. , 46 : 129 – 136 .
- Guzzella , L and Sora , S . 1998 . Mutagenic activity of lake water samples used as drinking water resources in Northern Italy . Water Res. , 32 : 1733 – 1742 .
- Camusso , M , Balestrini , R and Binelli , A . 2001 . Use of zebra mussel (Dreissena polymorpha) to assess trace metal contamination in the largest Italian subalpine lakes . Chemosphere , 44 : 263 – 270 .
- Binelli , A , Ricciardi , F , Riva , C and Provini , A . 2005 . Screening of POP pollution by AChE and EROD activities in Zebra mussels from the Italian Great Lakes . Chemosphere , 61 : 1074 – 1082 .
- Binelli , A , Ricciardi , F , Riva , C and Provini , A . 2006 . Integrated use of biomarkers and bioaccumulation data in Zebra mussel (Dreissena polymorpha) for site-specific quality assessment . Biomarkers , 11 : 428 – 448 .
- Bettinetti , R , Quadroni , S , Galassi , S , Bacchetta , R , Bonardi , L and Vailati , G . 2008 . Is meltwater from Alpine glaciers a secondary DDT source for lakes? . Chemosphere , 73 : 1027 – 1031 .
- Cortinovis , S , Galassi , S , Melone , G , Saino , N , Porte , C and Bettinetti , R . 2008 . Organochlorine contamination in the Great Crested Grebe (Podiceps cristatus): effects on eggshell thickness and egg steroid levels . Chemosphere , 73 : 320 – 325 .
- Riva , C , Binelli , A and Provini , A . 2008 . Evaluation of several priority pollutants in zebra mussels (Dreissena polymorpha) in the largest Italian subalpine lakes . Environ. Pollut. , 151 : 652 – 662 .
- Manca , M , Torretta , B , Comoli , P , Amsinck , SL and Jeppesen , E . 2007c . Major changes in trophic dynamics in large, deep sub-alpine Lake Maggiore from 1940s to 2002: a high resolution comparative palaeo-neolimnological study . Freshwater Biol. , 52 : 2256 – 2269 .
- Marchetto , A and Bettinetti , R . 1995 . Reconstruction of the phosphorus history of two deep, subalpine Italian lakes from sedimentary diatoms, compared with long-term chemical measurements . Mem. Ist. ital. Idrobiol. , 53 : 27 – 38 .
- Marchetto , A , Lami , A , Musazzi , S , Massaferro , J , Langone , L and Guilizzoni , P . 2004 . Lake Maggiore (N. Italy) trophic history: fossil diatom, plant pigments, and chironomids, and comparison with long-term limnological data . Quatern. Int. , 113 : 97 – 110 .
- Zilioli , E and Brivio , PA . 1997 . The satellite derived optical information for the comparative assessment of lacustrine water quality . Sci. Total Environ. , 196 : 229 – 245 .
- Brivio , PA , Giardino , C and Zilioli , E . 2001a . Determination of chlorophyll concentration changes in Lake Garda using an image-based radiative transfer code for Landsat TM images . Int. J. Remote Sens. , 22 : 487 – 502 .
- Giardino , C , Candiani , G and Zilioli , E . 2005 . Detecting chlorophyll-a in Lake Garda using TOA MERIS radiances . Photogramm. Eng. Rem. S. , 71 : 1045 – 1051 .
- Giardino , C , Bartoli , M , Candiani , G , Bresciani , M and Pellegrini , L . Recent changes in macrophyte colonisation patterns: an imaging spectrometry-based evaluation of southern Lake Garda (northern Italy), J. Appl. Remote Sens. 1, 011509 (2007a), 17 pp., DOI:10.1117/1.2834807
- Giardino , C , Brando , VE , Dekker , AG , Strombeck , N and Candiani , G . 2007b . Assessment of water quality in Lake Garda (Italy) using Hyperion . Remote Sens. Environ. , 109 : 183 – 195 .
- Bresciani , M , Stroppiana , D , Fila , G , Montagna , M and Giardino , C . 2009 . Monitoring reed vegetation in environmentally sensitive areas in Italy . Riv. Ital. Telerilevam. , 41 : 125 – 137 .
- Giardino , C , Pepe , M , Brivio , PA , Ghezzi , P and Zilioli , E . 2001 . Detecting chlorophyll, Secchi disk depth and surface temperature in a sub-alpine lake using Landsat imagery . Sci. Total Environ. , 268 : 19 – 29 .
- Pepe , M , Giardino , C , Borsani , G , Cardoso , AC , Chiaudani , G , Premazzi , G , Rodari , E and Zilioli , E . 2001 . Relationship between apparent optical properties and photosynthetic pigments in the sub-alpine Lake Iseo . Sci. Total Environ. , 268 : 31 – 45 .
- Brivio , PA , Giardino , C and Zilioli , E . 2001b . Validation of satellite data for quality assurance in lake monitoring applications . Sci. Total Environ. , 268 : 3 – 18 .
- de Bernardi , R . 1992 . Outline of limnological research in Italy . Mem. Ist. Ital. Idrobiol. , 50 : 1 – 17 .
- Ambrosetti , W and Barbanti , L . 1992 . Physical limnology in Italy: an historical review . Mem. Ist. Ital. Idrobiol. , 50 : 37 – 59 .
- Barbieri , A and Polli , B (eds.) . Limnological aspects and management of Lago di Lugano, Aquat. Sci. 54, 1992, pp. 179–390
- Nõges , P , Kangur , K , Nõges , T , Reinart , A , Simola , H and Viljanen , M . 2008 . Highlights of large lake research and management in Europe . Hydrobiologia , 599 : 259 – 276 .
- Håkanson , L . 2005a . The importance of lake morphometry and catchment characteristics in limnology–ranking based on statistical analyses . Hydrobiologia , 541 : 117 – 137 .
- Håkanson , L . 2005b . The importance of lake morphometry for the structure and function of lakes . Int. Rev. Hydrobiol. , 90 : 433 – 461 .
- Wetzel , RG . Limnology. Lake and River Ecosystems, 3rd ed., Academic Press, San Diego, CA, 2001
- Livingstone , DM . 1997 . An example of simultaneous occurrence of climate-driven ‘sawtooth’ deep-water warming/cooling episodes in several Swiss lakes . Verh. Int. Verein. Limnol. , 26 : 822 – 828 .
- Tilzer , MM . 1990 . Environmental and physiological control of phytoplankton productivity in large lakes, in Large Lakes, Ecological Structure and function, M.M. Tilzer and C. Serruya eds. , 339 – 367 . Berlin, Heidelberg : Springer-Verlag .
- Slugina , ZV . 2006 . Endemic Bivalvia in ancient lakes . Hydrobiologia , 568 : 213 – 217 .
- Stoma , I , Klyuchevskayaa , AA , Stomb , AD and Balayan , AE . 2009 . Experimental study of some Siberian and Baikal endemic species of hydrobionts: comparative analysis of oxyphility and frigophility . Contemp. Probl. Ecol. , 2 : 51 – 54 .
- IRSA (Istituto di Ricerca sulle Acque), Indagini sul Lago di Garda, IRSA Quaderni 18, Roma, 1974
- Tartari , G , Marchetto , A and Copetti , D . Qualità delle Acque Lacustri della Lombardia alle Soglie del 2000, Fondazione Lombardia per l’Ambiente (44), Milano, 2000
- Legendre , P and Legendre , L . Numerical Ecology, Elsevier Science BV, Amsterdam, 1998
- Yue , S and Pilon , P . 2004 . A comparison of the power of the t test, Mann-Kendall and bootstrap tests for trend detection . Hydrolog. Sci. J. , 49 : 21 – 37 .
- Yue , S , Pilon , P , Phinney , B and Cavadias , G . 2002 . The influence of autocorrelation on the ability to detect trend in hydrological series . Hydrol. Process. , 16 : 1807 – 1829 .
- R Development Core Team . R: a language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria (2009), http://www.R-project.org
- Forel , FA . Le Léman–Monographie Limnologique, Lousanne, 1892–1904
- Pavesi , P . 1883 . Altra serie di ricerche e studi sulla fauna pelagica dei laghi italiani . Atti Soc. Ven.-Tren. Sci. Nat. , 8 : 340 – 403 .
- De Marchi , M . 1912 . Appunti limnologici sul Verbano . Rend. R. Ist. Lomb. Sc. Lett. , 45 : 166 – 170 .
- Monti , R . La limnologia del Lario in relazione al ripopolamento delle acque e alla pesca, Luzzati, Roma, 1925
- Monti , R . 1929 . Limnologia comparata dei laghi insubrici . Verh. Int. Verein. Limnol. , 4 : 462 – 497 .
- Baldi , E . 1949 . Alcuni caratteri generali dei laghi marginali sudalpini . Verh. Int. Verein. Limnol. , 10 : 50 – 69 .
- Manca , M , Calderoni , A and Mosello , R . 1992 . Limnological reserch in Lago Maggiore: studies on hydrochemistry and plankton . Mem. Ist. Ital. Idrobiol. , 50 : 171 – 200 .
- Vollenweider , RA . 1964 . Ueber oligomiktische Verhaltnisse des Lago Maggiore und einiger anderer insubrischer Seen . Mem. Ist. ital. Idrobiol. , 17 : 191 – 206 .
- Vollenweider , RA . 1965 . Materiali ed idee per una idrochimica delle acque insubriche . Mem. Ist. Ital. Idrobiol. , 19 : 213 – 286 .
- Ambrosetti , W , Barbanti , L , Mosello , R and Ruggiu , D (eds.) . Mescolamento, Caratteristiche Chimiche, Fitoplancton e Situazione Trofica nei Laghi Profondi Subalpini, C.N.R. Progetto Finalizzato ‘Promozione della qualità dell’ambiente’, AQ/2/20, 1983, pp. 1–151
- Mosello , R and Salmaso , N . 2000 . Cooperazione per lo studio della qualità delle acque dei laghi profondi sudalpini (QuAlps): obiettivi e primi risultati, Atti della Giornata di Studio: I Laghi come Risorsa per lo Sviluppo Milano, 16 ottobre 1998, Politecnico di Milano, Consiglio Regionale della Lombardia, CNR-III Pallanza e Associazione ex Consiglieri Regionali della Lombardia 39 – 50 .
- Mosello , R and Giussani , G (eds.) . Evoluzione recente della qualità delle acque dei laghi profondi sudalpini, Docum. Ist. Ital. Idrobiol. 61 (1997), pp. 1–228
- Beniston , M , Rebetez , M , Giorgi , F and Marinucci , MR . 1994 . An analysis of regional climate change in Switzerland . Theor. Appl. Climatol. , 49 : 135 – 159 .
- Coats , RN , Perez-Losada , J , Schladow , G , Richards , R and Goldman , CR . 2006 . The warming of Lake Tahoe . Climatic Change , 76 : 121 – 148 .
- Rogora , M and Mosello , R . 2007 . Climate as a confounding factor in the response of surface water to nitrogen deposition in an area South of the Alps . Appl. Geochem. , 22 : 1122 – 1128 .
- OECD, Eutrophication of Waters. Monitoring, Assessment and Control, OECD, Paris, 1982
- Ambrosetti , W , Barbanti , L , Mosello , R and Pugnetti , A . 1992 . Limnological studies on the deep southern Alpine lakes Maggiore, Lugano, Como, Iseo and Garda . Mem. Ist. Ital. Idrobiol. , 50 : 117 – 146 .
- Bertoni , R and Callieri , C . 1992 . Research trends in the study of microbial loop . Mem. Ist. Ital. Idrobiol. , 50 : 107 – 116 .
- Azam , F , Fenchel , T , Field , JG , Gray , JS , Meyer-Reil , LA and Thingstad , F . 1983 . The ecological role of water-column microbes in the sea . Mar. Ecol.-Prog. Ser. , 10 : 257 – 263 .
- Bonacina , C , Bonomi , G and Pasteris , A . 1992 . Some remarks on the macrobenthos community of the profundal zone of the large Italian lakes . Mem. Ist. Ital. Idrobiol. , 50 : 79 – 106 .
- Alikas , K and Reinart , A . 2008 . Validation of the MERIS products on large european lakes: Peipsi, Vanern and Vattern . Hydrobiologia, , 599 : 161 – 168 .
- Begon , M , Harper , JL and Townsend , CR . Ecology: individuals, populations and communities, Blackwell Scientific Publications, Oxford, 1990
- Kalff , J . Limnology, Prentice Hall, Upper Saddle River, NJ, 2002
- Stow , CA , Carpenter , SR , Webster , KE and Frost , TM . 1998 . Long-Term environmental monitoring: some perspectives from lakes . Ecol. Appl. , 8 : 269 – 276 .
- Peters , RH . 1990 . Pathologies in limnology . Mem. Ist. Ital. Idrobiol. , 47 : 181 – 217 .
- Magnuson , JJ , Kratz , T and Benson , BJ . Long-Term Dynamics of Lakes in the Landscape–Long-Term Ecological Research on North Temperate Lakes, Oxford University Press, Oxford, UK, 2005
- Peñuelas , J and Sardans , J . 2009 . Ecological metabolomics . Chem. Ecol. , 25 : 305 – 309 .
- Harris , G . 1999 . This is not the end of limnology (or of science): the world may well be a lot simpler than we think . Freshwater Biol. , 42 : 689 – 706 .
- Sterner , RW . 2008 . On the phosphorus limitation paradigm for lakes . Int. Rev. Hydrobiol. , 93 : 433 – 445 .
- Weisse , T . 2004 . Pelagic Microbes–Protozoa and the Microbial Food Web in The Lakes Handbook Volume 1–Limnology and Limnetic Ecology P.E. O'sullivan and C.S. Reynolds, eds. 417 – 460 . USA : Blackwell Science Ltd, .