
Abstract
Sedentary lifestyle is linked with poor health, most commonly obesity and associated disorders, the corollary being that exercise offers a preventive strategy. However, the scope of exercise biology extends well beyond energy expenditure and has emerged as a great ‘polypill’, which is safe, reliable and cost-effective not only in disease prevention but also treatment. Biological mechanisms by which exercise influences homeostasis are becoming clearer and involve multi-organ systemic adaptations. Most of the elements of a modern lifestyle influence the indigenous microbiota but few studies have explored the effect of increased physical activity. While dietary responses to exercise obscure the influence of exercise alone on gut microbiota, professional athletes operating at the extremes of performance provide informative data. We assessed the relationship between extreme levels of exercise, associated dietary habits and gut microbiota composition, and discuss potential mechanisms by which exercise may exert a direct or indirect influence on gut microbiota.
Introduction
Changes in human behavior have influenced not only the external environment but also the inner microbial environment. This has been particularly evident as societies undergo socio-economic development. The microbiota may be considered as the proximate environmental factor conferring risk or resistance to a range of chronic inflammatory and metabolic disorders that are common in socio-economically developed societies. Underpinning host-environmental interactions is a signaling network among the microbiota, host metabolism and host immunity, with lifestyle factors such as diet influencing each component of this triad. Most of the elements of a modern society have a modifying effect on the indigenous microbiota; but one that has received comparatively little attention is exercise. Exercise is well known for its metabolic and immunologic effects in the host but an impact on the microbiota has been uncertain.
Why is this important? Other than diet, few environmental variables in the modern lifestyle of a host exert such a level of control across a range of physiological events. We have shown that frailty and poor exercise capacity in the elderly is linked with low fecal microbial diversity that correlates with poor dietary diversity.Citation1 The possibility that exercise or combined dietary change might enhance microbial diversity has immediate implications not limited to the elderly. The growing list of reported health benefits from exercise now includes: limitation of age-related cognitive impairment,Citation2 prevention of colon cancerCitation3 and the treatment of diabetes,Citation4 irritable bowel syndromeCitation5 and depression.Citation6 The microbiota and diet-microbe-host immune and metabolic interactions have been implicated in the etiopathogenesis of all of these conditions.
The influence of physical activity on intestinal microbiota has largely been understudied. The majority of work to date pertains to murine models examining the combined effect of exercise and dietary interventions such as low energy or high fat diets.Citation7 However, the anti-inflammatory effect of regular exercise and modulation of infection risk has been the subject of much research.Citation8 Of late, exercise's ability to enable cross talk between skeletal muscle and multiple other organs (e.g. brain, bone, liver, adipose tissue, gut) has become increasingly recognized.Citation9,10
Training-related skeletal muscle plasticity, derives from a complex set of physiological and molecular processes, and is subject to the exercise mode engaged. In particular, peroxisome proliferator-activator receptor-G-coactivator-1α (PGC-1α), abundantly expressed in skeletal muscle tissue, is thought to play a key role in muscular adaptability in response to resistance or weights training.Citation11 PGC-1α, a regulator of mitochondrial biogenesis within skeletal tissue, targets a diverse range of transcription factors in distant tissues, with multiple downstream effects.Citation12
Here, we summarise recent work with professional athletes, operating at the extremes of performance, as a human model to explore diet-exercise-microbe inter-relationships and discuss potential mechanisms by which exercise might influence the microbiota.
A study of the extreme
To begin to explore the relationship among exercise, associated dietary adjustments, and the microbiota in humans, we reasoned that professional athletes operating at the extremes of fitness and performance would be an optimal study group in which to detect biologically significant deviations from controls. We studied the Irish International rugby football team during a pre-World Cup training camp in which dietary intake and physical activity were monitored.Citation13,14 A comprehensive range of observational and laboratory assessments were performed including: pattern and content of dietary intake, metagenomic fecal microbiota compositional analysis, body composition (by DEXA scanning), pro- and anti-inflammatory cytokines, and creatine kinase. It is noteworthy that many professional rugby athletes might be classified as overweight or obese, due to their large muscle mass, if assessed by body mass index (BMI) alone. Body size of rugby players is, in part, related to their specialist position within the team formation; forwards have high BMI and back players tend to have a lower or normal BMI. In anticipation of the possibility that BMI might directly or indirectly have a confounding influence on the interpretation of results, we recruited 2 control groups of healthy non-professional athletes with varying levels of physical activity, i.e. a low BMI group (BMI < 25) and a high BMI group (BMI > 28). As expected, the elite professional athletes had a higher lean body mass, lower body fat percentage and waist:hip ratios compared to controls. The rugby professionals grazed throughout the day whereas controls ate at conventional meal times. The professional athletes also consumed significantly higher quantities of protein, fat, carbohydrates, sugar, cholesterol and saturated fat per day than both control groups and consumed significantly higher quantities of starch, fiber, mono-unsaturated fat and poly-unsaturated fat than the high BMI control group (details of diet were extrapolated from food frequency questionnaires). Remarkably, despite extremes of exercise as evidenced by extraordinarily high levels of creatine kinase in serum (serving as a proxy for degree of exercise) compared to the control groups, the professional athletes had lower levels of inflammatory cytokines than the controls. In addition, they had increased fecal microbial diversity. From a taxonomic perspective, this increased diversity was reflected by the presence of representatives of 22 phyla of bacteria in the athlete microbiota, in contrast to 11 and 9 phyla in the low and high BMI controls, respectively. Notably, among the taxa that were enriched in the athlete and low BMI populations relative to their high BMI counterparts was the lean phenotype-associated, Akkermansia muciniphila.Citation15,16 The increased gut microbial diversity in athletes correlated with both protein intake and creatine kinase levels ().
Table 1. Correlations between exercise intensity (creatine kinase), diet (protein intake) and diversity (estimated over 5 metrics; chao1, Simpson, Shannon, phylogenetic diversity and observed species). Table displays Pearson correlation coefficients followed by p-value in all incidences. P >0.05 was taken to be significant
Limitations and prospects
While we were careful in our interpretation of the data not to transmute correlation into causation it may be tempting to view increased microbial diversity as a benefit of exercise. However, this remains to be demonstrated by a prospective rather than cross-sectional study of volunteers undergoing a structured exercise program which attempts to control for diet. Such a study is underway and will focus on healthy controls rather than the elite professional athlete. Furthermore, in addition to compositional analysis, whole genome sequencing for functional genomic analysis along with metabolomic profiling will be included to ascertain which, if any, functional pathways are altered during exercise training. Diet-exercise may represent an inseparable interaction, the contributions of each being impossible to assess in isolation. It is noteworthy that exercise as a weight reduction strategy is often poorly efficacious unless combined with dietary restriction,Citation17 as an increase in physical activity by exercise-naïve individuals may result in unanticipated dietary change, driven by physiological responses to exercise and a risk of compensatory over-eating post-exercise.Citation18 Appetite response to exercise is complex and appears to be highly variable between individuals.Citation19 It is dependent on multiple factors including mental attitude toward exercise,Citation20 the intensity of the activity involved Citation21 and prior reliance on substrate availability for energy expenditure.Citation22 Moderately intense physical activity is followed by a transient surge in serum ghrelin levels in the post-exercise period. This rise is not apparent however following high intensity exercise.Citation21 The close relationship between diet and exercise obscures the study of exercise and microbiota in humans. Detailed analysis of baseline and subsequent dietary changes are essential.
Exercise biology and the microbiota – potential mechanism of interaction
Although a direct causal relationship between exercise and gut microbial composition or function has not been established and is inextricably linked with dietary adjustments, several potential mechanisms by which physical activity and fitness might modify the microbiota can be entertained. A schematic representation of some of the points of interaction is shown in .
Figure 1. Schematic overview of potential sites of interaction between the biological adaptations to exercise and the microbiota. This is intended to be representative not comprehensive. Exercise is linked with a diversity of biological responses including a modifying influence on the brain-gut-microbe axis, diet-microbe-host metabolic interactions, neuro-endocrine and neuro-immune interactions. For example, exercise is long known to increase vagal tone - the hard wiring of the gut - which is anti-inflammatory and immune-modulatory. The latter might represent an indirect means by which exercise conditions gut microbiota composition.
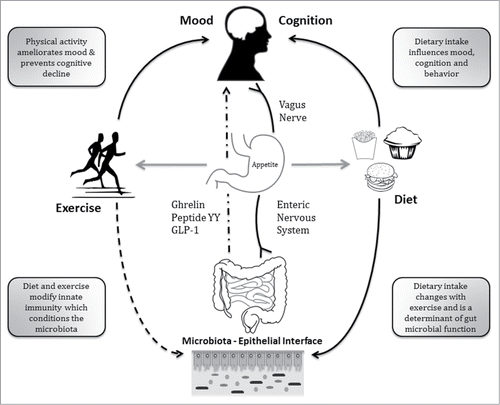
Abrupt exercise involves the production of multiple metabolites and inflammatory mediators,Citation23 but habitual exercise and fitness leads to suppression of basal pro-inflammatory cytokines (as found in our study)Citation13 indicating a regulatory loop between exercise biology and host immunity. Regular physical activity is a recognized treatment for multiple inflammatory-centered conditions. The anti-inflammatory effect of regular exercise results in improved immunological profile in type 2 diabetes mellitus,Citation24 coronary artery disease,Citation25 peripheral arterial diseaseCitation26 and obesity.Citation27 In particular, combined aerobic and resistance training in type 2 diabetics leads to a decrease in pro-inflammatory cytokines IL-1β, IL-6, TNF-α, IFN-γ as well as an increase in anti-inflammatory modulators IL-4 and IL-10.Citation24 Among elderly subjects, aerobic and resistance training individually attenuate inflammatory tone but may take up to 6-months to take effect.Citation28
Whether intestinal immunological responses to exercise mirror those of the systemic immune compartment is unclear but seems likely. Strenuous aerobic activity in healthy mice leads to an immediate increase in TNF-α expression in intestinal lymphocytes,Citation29 but appears to be suppressed below pre-exercise levels at 24-hours post-exercise.Citation30 Anti-inflammatory IL-10 expression in intestinal lymphocytes is also increased immediately post-exercise.Citation30,31 Furthermore, regular low to moderate intensity exercise reduces colonic oxidative insult in a rat model of colitis.Citation32
Of course, prolonged excessive exercise has a deleterious influence on intestinal function. Intense exercise redistributes blood from the splanchnic circulation to actively respiring tissues.Citation33 Prolonged intestinal hypo-perfusion impairs mucosal homeostasis and causes enterocyte injury.Citation34 Intestinal ischemia may result, particularly in the setting of dehydration, manifesting as abdominal cramps, diarrhea or occasionally bloody diarrhea.Citation35 This is particularly true in endurance sports.Citation36 As a result, increased intestinal permeability ensues, thought to be driven by the phosphorylation of several tight junction proteins.Citation37 These events render the gut mucosa susceptible to endotoxin translocation.Citation38 By contrast, regular moderate exercise mitigates the effects of stress-induced intestinal barrier dysfunction. Moderate exercise has been associated with a lesser degree of intestinal permeability, preservation of mucous thickness and lower rates of bacterial translocation along with up-regulation of anti-microbial protein production and gene expression in small intestinal tissue (α-defensin, β-defensin, Reg IIIb and Reg IIIc).Citation39 In a mouse model of colitis, forced treadmill exercise training aggravated inflammatory indices and outcomes, while voluntary wheel training displayed a protective effect.Citation40 This suggests a biphasic and dose-response effect of exercise on gut health that has yet to be fully explored in humans.
The brain-gut axis is an established bi-directional avenue of signaling between 2 organs both of which are influenced by exercise. Several lines of evidence have extended this to include the microbiota.Citation41 Information exchange along the brain-gut-microbe axis is hard wired by the vagus nerve, neuroendocrine mediators including the hypothalamic-pituitary-adrenal axis and a variety of neurotransmitter and local hormones, some of which are also produced by the microbiota.Citation41,42 This regulates several homeostatic processes including appetite, satiety and digestion. The vagus also regulates gastro-intestinal inflammatory tone, direct visualization of which has been shown with a pro-inflammatory response to vagotomy in mice.Citation43 The presence of a vagally driven anti-inflammatory pathway is now establishedCitation44 and cholinergic innervation to intestinal sub-mucosa may exert influence on mucosal innate immunity.Citation45 Evidence suggests that this parasympathetic neuro-immune reflex is dependent on vagal afferent neurons for the local release of intestinal inflammatory mediators in response to pathogenic gut bacteria.Citation46 Therefore, elevated vagal tone and parasympathetic influence, as seen in the resting state of athletes, may foster a preferential anti-inflammatory milieu at the intestinal-luminal interface, with an attendant conditioning influence on microbial composition.
Finally, exercise prescription is increasingly recognized as a safe and effective treatment for disorders ranging from depressionCitation6 and irritable bowel syndromeCitation5 to inflammatory bowel diseaseCitation47 adding further support for the potential influence of exercise on the brain-gut-microbiome axis. By contrast, withdrawal of exercise from individuals accustomed to regular activity leads to deterioration in mood in as little as 2 weeks post-exercise.Citation48 Furthermore, plasma kynurenine metabolite levels, which are strongly correlated with depression,Citation49 are subject to influence by exercise-related PGC-1α1 expression. Increased expression of muscle-specific PGC-1α1 in transgenic mice is associated with greater resilience to chronic stress and stress-induced CNS inflammation.Citation50 Thus, there are several points at which exercise biology and accompanying dietary adjustments may interact directly or indirectly with host-microbe interactions in the gut ().
Conclusion
Whether the gut microbiota of sedentary subjects can be “trained” to resemble that of an elite athlete through exercise, with increased protein intake or a combination of both is an intriguing prospect. It may be impossible to truly separate the physical act of exercise from the accompanying dietary adjustments that volunteers make, often unwittingly, during an exercise program. In addition, it is presently unclear if there is a dose-response between exercise levels and beneficial alterations in microbial composition or function. Regardless, determination of optimal diet-exercise combination programmes which might give the elite athlete ‘an edge’ over competitors is worthy of careful pursuit including in sub-elite non-professional volunteers and may ultimately inform our advice regarding such activity in patients with a variety of disorders which could benefit from a personalised training program.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
The authors are supported in part by research grants from Science Foundation Ireland including a center grant (Alimentary Pharmabiotic Center; Grant Numbers SFI/12/RC/2273 and 12/RC/2273). Dr. Orla O’Sullivan is funded by a Starting Investigator Research Grant from Science Foundation Ireland (Grant number 13/SIRG/2160). Dr. Paul Cotter is funded by a Principal Investigator Award from Science Foundation Ireland P.D.C are supported by a SFI PI award (Grant number 11/PtdIns/1137).
References
- Claesson MJ, Jeffery IB, Conde S, Power SE, O'Connor EM, Cusack S, Harris HM, Coakley M, Lakshminarayanan B, O'Sullivan O, et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012; 488:178-84; PMID:22797518; http://dx.doi.org/10.1038/nature11319
- Bherer L, Erickson KI, Liu-Ambrose T. A review of the effects of physical activity and exercise on cognitive and brain functions in older adults. J Aging Res 2013; 2013: Article ID 657508
- Robsahm TE, Aagnes B, Hjartaker A, Langseth H, Bray FI, Larsen IK. Body mass index, physical activity, and colorectal cancer by anatomical subsites: a systematic review and meta-analysis of cohort studies. Eur J Cancer Prev 2013; 22:492-505; PMID:23591454; http://dx.doi.org/10.1097/CEJ.0b013e328360f434
- Schwingshackl L, Missbach B, Dias S, Konig J, Hoffmann G. Impact of different training modalities on glycaemic control and blood lipids in patients with type 2 diabetes: a systematic review and network meta-analysis. Diabetologia 2014; 57:1789-97; PMID:24996616; http://dx.doi.org/10.1007/s00125-014-3303-z
- Johannesson E, Simren M, Strid H, Bajor A, Sadik R. Physical activity improves symptoms in irritable bowel syndrome: a randomized controlled trial. Am J Gastroenterol 2011; 106:915-22; PMID:21206488; http://dx.doi.org/10.1038/ajg.2010.480
- Harkin A. Muscling In on Depression. N Engl J Med 2014; 371:2333-4; PMID:25494274; http://dx.doi.org/10.1056/NEJMcibr1411568
- Evans CC, LePard KJ, Kwak JW, Stancukas MC, Laskowski S, Dougherty J, Moulton L, Glawe A, Wang Y, Leone V, et al. Exercise prevents weight gain and alters the gut microbiota in a mouse model of high fat diet-induced obesity. PLoS One 2014; 9:e92193; PMID:24670791; http://dx.doi.org/10.1371/journal.pone.0092193
- Pedersen BK, Akerstrom TC, Nielsen AR, Fischer CP. Role of myokines in exercise and metabolism. J Appl Physiol (1985) 2007; 103:1093-8; PMID:17347387; http://dx.doi.org/10.1152/japplphysiol.00080.2007
- Handschin C, Spiegelman BM. The role of exercise and PGC1alpha in inflammation and chronic disease. Nature 2008; 454:463-9; PMID:18650917; http://dx.doi.org/10.1038/nature07206
- Hawley JA, Hargreaves M, Joyner MJ, Zierath JR. Integrative Biology of Exercise. Cell 2014; 159:738-49; PMID:25417152; http://dx.doi.org/10.1016/j.cell.2014.10.029
- Ruas JL, White JP, Rao RR, Kleiner S, Brannan KT, Harrison BC, Greene NP, Wu J, Estall JL, Irving BA, et al. A PGC-1alpha isoform induced by resistance training regulates skeletal muscle hypertrophy. Cell 2012; 151:1319-31; PMID:23217713; http://dx.doi.org/10.1016/j.cell.2012.10.050
- Lin J, Handschin C, Spiegelman BM. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab 2005; 1:361-70; PMID:16054085; http://dx.doi.org/10.1016/j.cmet.2005.05.004
- Clarke SF, Murphy EF, O'Sullivan O, Lucey AJ, Humphreys M, Hogan A, Hayes P, O'Reilly M, Jeffery IB, Wood-Martin R, et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014; 63:1913-20; PMID:25021423; http://dx.doi.org/10.1136/gutjnl-2013-306541
- Hold GL. The gut microbiota, dietary extremes and exercise. Gut 2014; 63:1838-9; PMID:25021422; http://dx.doi.org/10.1136/gutjnl-2014-307305
- Walters WA, Xu Z, Knight R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Letters 2014; 588:4223-33; PMID:25307765; http://dx.doi.org/10.1016/j.febslet.2014.09.039
- Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, Guiot Y, Derrien M, Muccioli GG, Delzenne NM, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci U S A 2013; 110:9066-71; PMID:23671105; http://dx.doi.org/10.1073/pnas.1219451110
- Johns DJ, Hartmann-Boyce J, Jebb SA, Aveyard P. Diet or Exercise Interventions vs Combined Behavioral Weight Management Programs: A Systematic Review and Meta-Analysis of Direct Comparisons. J Acad Nutr Diet 2014; 114:1557-68; PMID:25257365; http://dx.doi.org/10.1016/j.jand.2014.07.005
- Thomas DM, Bouchard C, Church T, Slentz C, Kraus WE, Redman LM, Martin CK, Silva AM, Vossen M, Westerterp K, et al. Why do individuals not lose more weight from an exercise intervention at a defined dose? An energy balance analysis. Obes Rev 2012; 13:835-47; PMID:22681398; http://dx.doi.org/10.1111/j.1467-789X.2012.01012.x
- Hopkins M, Blundell JE, King NA. Individual variability in compensatory eating following acute exercise in overweight and obese women. Br J Sports Med 2014; 48:1472-6; PMID:23666018; http://dx.doi.org/10.1136/bjsports-2012-091721
- Werle CO, Wansink B, Payne CR. Just thinking about exercise makes me serve more food. Physical activity and calorie compensation. Appetite 2011; 56:332-5; PMID:21185895; http://dx.doi.org/10.1016/j.appet.2010.12.016
- Bilski J, Manko G, Brzozowski T, Pokorski J, Nitecki J, Nitecka E, Wilk-Franczuk M, Ziolkowski A, Jaszczur-Nowicki J, Kruczkowski D, et al. Effects of exercise of different intensity on gut peptides, energy intake and appetite in young males. Ann Agric Environ Med 2013; 20:787-93; PMID:24364454
- Pannacciulli N, Salbe AD, Ortega E, Venti CA, Bogardus C, Krakoff J. The 24-h carbohydrate oxidation rate in a human respiratory chamber predicts ad libitum food intake. Am J Clin Nutr 2007; 86:625-32; PMID:17823426
- Pedersen BK. Special feature for the Olympics: effects of exercise on the immune system: exercise and cytokines. Immunol Cell Biol 2000; 78:532-5; PMID:11050536; http://dx.doi.org/10.1111/j.1440-1711.2000.t01-11-.x
- Balducci S, Zanuso S, Nicolucci A, Fernando F, Cavallo S, Cardelli P, Fallucca S, Alessi E, Letizia C, Jimenez A, et al. Anti-inflammatory effect of exercise training in subjects with type 2 diabetes and the metabolic syndrome is dependent on exercise modalities and independent of weight loss. Nutr, Metab Cardiovasc Dis 2010; 20:608-17; PMID:19695853; http://dx.doi.org/10.1016/j.numecd.2009.04.015
- Lara Fernandes J, Serrano CV, Jr., Toledo F, Hunziker MF, Zamperini A, Teo FH, Oliveira RT, Blotta MH, Rondon MU, Negrao CE. Acute and chronic effects of exercise on inflammatory markers and B-type natriuretic peptide in patients with coronary artery disease. Clin Res Cardiol 2011; 100:77-84; PMID:20835904; http://dx.doi.org/10.1007/s00392-010-0215-x
- Tisi PV, Hulse M, Chulakadabba A, Gosling P, Shearman CP. Exercise training for intermittent claudication: does it adversely affect biochemical markers of the exercise-induced inflammatory response? Eur J Vasc Endovasc Surg 1997; 14:344-50; PMID:9413374; http://dx.doi.org/10.1016/S1078-5884(97)80283-3
- Ho SS, Dhaliwal SS, Hills AP, Pal S. Effects of chronic exercise training on inflammatory markers in Australian overweight and obese individuals in a randomized controlled trial. Inflammation 2013; 36:625-32; PMID:23250821; http://dx.doi.org/10.1007/s10753-012-9584-9
- Martins RA, Neves AP, Coelho-Silva MJ, Verissimo MT, Teixeira AM. The effect of aerobic versus strength-based training on high-sensitivity C-reactive protein in older adults. Eur J Appl Physiol 2010; 110:161-9; PMID:20437055; http://dx.doi.org/10.1007/s00421-010-1488-5
- Pervaiz N, Hoffman-Goetz L. Immune cell inflammatory cytokine responses differ between central and systemic compartments in response to acute exercise in mice. Exerc Immunol Rev 2012; 18:142-57; PMID:22876726
- Hoffman-Goetz L, Spagnuolo PA, Guan J. Repeated exercise in mice alters expression of IL-10 and TNF-alpha in intestinal lymphocytes. Brain, Behav Immun 2008; 22:195-9; PMID:17720362; http://dx.doi.org/10.1016/j.bbi.2007.07.002
- Hoffman-Goetz L, Pervaiz N, Packer N, Guan J. Freewheel training decreases pro- and increases anti-inflammatory cytokine expression in mouse intestinal lymphocytes. Brain, Behav Immun 2010; 24:1105-15; PMID:20510350; http://dx.doi.org/10.1016/j.bbi.2010.05.001
- Kasimay O, Guzel E, Gemici A, Abdyli A, Sulovari A, Ercan F, Yegen BC. Colitis-induced oxidative damage of the colon and skeletal muscle is ameliorated by regular exercise in rats: the anxiolytic role of exercise. Exp Physiol 2006; 91:897-906; PMID:16763006; http://dx.doi.org/10.1113/expphysiol.2006.034439
- Qamar MI, Read AE. Effects of exercise on mesenteric blood flow in man. Gut 1987; 28:583-7; PMID:3596339; http://dx.doi.org/10.1136/gut.28.5.583
- van Wijck K, Lenaerts K, van Loon LJ, Peters WH, Buurman WA, Dejong CH. Exercise-induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS One 2011; 6:e22366; PMID:21811592; http://dx.doi.org/10.1371/journal.pone.0022366
- de Oliveira EP, Burini RC. The impact of physical exercise on the gastrointestinal tract. Curr Opin Clin Nutr Metab Care 2009; 12:533-8; PMID:19535976; http://dx.doi.org/10.1097/MCO.0b013e32832e6776
- Oktedalen O, Lunde OC, Opstad PK, Aabakken L, Kvernebo K. Changes in the gastrointestinal mucosa after long-distance running. Scand J Gastroenterol 1992; 27:270-4; PMID:1589703; http://dx.doi.org/10.3109/00365529209000073
- Zuhl M, Schneider S, Lanphere K, Conn C, Dokladny K, Moseley P. Exercise regulation of intestinal tight junction proteins. Br J Sports Med 2014; 48:980-6; PMID:23134759; http://dx.doi.org/10.1136/bjsports-2012-091585
- Brock-Utne JG, Gaffin SL, Wells MT, Gathiram P, Sohar E, James MF, Morrell DF, Norman RJ. Endotoxaemia in exhausted runners after a long-distance race. S Afr Med J 1988; 73:533-6; PMID:3375945
- Luo B, Xiang D, Nieman DC, Chen P. The effects of moderate exercise on chronic stress-induced intestinal barrier dysfunction and antimicrobial defense. Brain, Behav Immun 2014; 39:99-106; PMID:24291325; http://dx.doi.org/10.1016/j.bbi.2013.11.013
- Cook MD, Martin SA, Williams C, Whitlock K, Wallig MA, Pence BD, Woods JA. Forced treadmill exercise training exacerbates inflammation and causes mortality while voluntary wheel training is protective in a mouse model of colitis. Brain, Behav Immun 2013; 33:46-56; PMID:23707215; http://dx.doi.org/10.1016/j.bbi.2013.05.005
- Grenham S, Clarke G, Cryan JF, Dinan TG. Brain-gut-microbe communication in health and disease. Fron Physiol 2011; 2:94 eCollection 2011; PMID:22162969; http://dx.doi.org/10.3389/fphys.2011.00094
- Stilling RM, Bordenstein SR, Dinan TG, Cryan JF. Friends with social benefits: host-microbe interactions as a driver of brain evolution and development? Front Cell Infect Microbiol 2014; 4:147; PMID:25401092; http://dx.doi.org/10.3389/fcimb.2014.00147
- O'Mahony C, van der Kleij H, Bienenstock J, Shanahan F, O'Mahony L. Loss of vagal anti-inflammatory effect: in vivo visualization and adoptive transfer. Am J Physiol Regul, Integr Comp Physiol 2009; 297:R1118-26; PMID:19675277; http://dx.doi.org/10.1152/ajpregu.90904.2008
- Tracey KJ. The inflammatory reflex. Nature 2002; 420:853-9; PMID:12490958; http://dx.doi.org/10.1038/nature01321
- de Jonge WJ. The Gut's Little Brain in Control of Intestinal Immunity. ISRN Gastroenterol 2013; 2013:630159; PMID:23691339; http://dx.doi.org/10.1155/2013/630159
- Riley TP, Neal-McKinney JM, Buelow DR, Konkel ME, Simasko SM. Capsaicin-sensitive vagal afferent neurons contribute to the detection of pathogenic bacterial colonization in the gut. J Neuroimmunol 2013; 257:36-45; PMID:23481698; http://dx.doi.org/10.1016/j.jneuroim.2013.01.009
- Khalili H, Ananthakrishnan AN, Konijeti GG, Liao X, Higuchi LM, Fuchs CS, Spiegelman D, Richter JM, Korzenik JR, Chan AT. Physical activity and risk of inflammatory bowel disease: prospective study from the Nurses' Health Study cohorts. Bmj 2013; 347:f6633; PMID:24231178; http://dx.doi.org/10.1136/bmj.f6633
- Kop WJ, Weinstein AA, Deuster PA, Whittaker KS, Tracy RP. Inflammatory markers and negative mood symptoms following exercise withdrawal. Brain, Behav Immun 2008; 22:1190-6; PMID:18619772; http://dx.doi.org/10.1016/j.bbi.2008.05.011
- Claes S, Myint AM, Domschke K, Del-Favero J, Entrich K, Engelborghs S, De Deyn P, Mueller N, Baune B, Rothermundt M. The kynurenine pathway in major depression: haplotype analysis of three related functional candidate genes. Psychiatry Res 2011; 188:355-60; PMID:21492941; http://dx.doi.org/10.1016/j.psychres.2011.03.012
- Agudelo LZ, Femenia T, Orhan F, Porsmyr-Palmertz M, Goiny M, Martinez-Redondo V, Correia JC, Izadi M, Bhat M, Schuppe-Koistinen I, et al. Skeletal muscle PGC-1alpha1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 2014; 159:33-45; PMID:25259918; http://dx.doi.org/10.1016/j.cell.2014.07.051