
ABSTRACT
Akkermansia muciniphila is a common member of the intestinal microbiota of healthy human individuals. Its abundance is negatively associated with inflammatory bowel disease and metabolic disorders and the oral administration of A. muciniphila improves the symptoms of metabolic disease in mice. Therefore, A. muciniphila is a promising candidate for the treatment of type-2 diabetes and obesity. However, some studies using animal models of intestinal inflammation reported that A. muciniphila may exacerbate gut inflammation. Because of these contradictory reports the present study aimed to clarify the role of A. muciniphila in the development of intestinal inflammation and the conditions promoting it. For this purpose, the short-term colitogenic potential of A. muciniphila strain ATCC BAA-835 was investigated in colitis-prone, gnotobiotic IL-10-deficient (Il10-/-) mice. Il10-/- mice mono-associated with A. muciniphila showed no signs of intestinal inflammation based on body-weight change, histopathological scoring and inflammatory markers. Additional association of the mice with the colitogenic Escherichia coli strain NC101 led to cecal but not colonic inflammation. However, the severity of the inflammation did not exceed that observed in mice mono-associated with E. coli NC101. Il10-/- mice colonized with a simplified human intestinal microbiota showed increased histopathology, but no increase in inflammatory markers. Furthermore, co-colonization with A. muciniphila did not modify histopathology. The turnover of intestinal mucus was similar in all groups despite the mucus-degrading property of A. muciniphila. Overall, the data do not support a short-term pro-inflammatory effect of A. muciniphila strain ATCC BAA-835 in the Il10-/- mouse model for inflammatory bowel disease.
Introduction
The human digestive tract is colonized by a complex microbial community which influences host health in many ways. The intestinal microbiota degrades undigested food components such as dietary fiber, catalyzes the bio-activation of non-nutritive food constituents such as polyphenols, impacts on the development of gut tissue, and promotes the maturation and maintenance of the gut-associated immune system.Citation1 Akkermansia muciniphila, a member of the Verrucomicrobia, is an abundant and common inhabitant of the healthy human gut.Citation2,Citation3 This bacterium was first isolated from human feces because of its ability to grow on mucin.Citation4 A. muciniphila also colonizes the intestinal tract of various animals including rodents.Citation5 The abundance of A. muciniphila is reduced in patients suffering from Crohn’s disease (CD) and ulcerative colitis.Citation6 Moreover, the severity of appendicitis correlates negatively with the fecal abundance of A. muciniphila.Citation7 Reduced fecal cell numbers of A. muciniphila are also linked to obesity and metabolic disorders in both mice and humans.Citation8–Citation12 The direct administration of A. muciniphila or of dietary factors, such as polyphenols, that promote the growth of endogenous A. muciniphila improves the metabolic profile of diet-induced obese mice.Citation12–Citation14 Therefore, the oral administration of A. muciniphila has been proposed as a potential treatment of metabolic disorders.Citation12,Citation15,Citation16 In conflict with these reports about beneficial effects of the organism, several studies suggest that A. muciniphila may compromise gut health. For example, A. muciniphila was reported to exacerbate the inflammatory response in the murine Salmonella enterica Typhimurium infection model of acute intestinal inflammation and in dextran sodium sulfate (DSS)-induced acute colitis.Citation17,Citation18 Moreover, stool samples from colorectal cancer patients displayed four-fold higher cell counts of A. muciniphila compared with healthy controls.Citation19 These inconsistent effects of A. muciniphila require clarification.
Interleukin-10-deficient (Il10-/-) mice develop chronic intestinal inflammation starting before three weeks of age when in contact with normal bacterial antigens and thereby mimic some features of CD.Citation20,Citation21 Onset, severity and site of the colitis are dependent on the genetic background of the mice and on the composition of the gut microbiota.Citation22–Citation24 Mono-association of Il10-/- mice with Helicobacter hepaticus, Pseudomonas fluorescens, viridans group streptococci, Clostridium sordellii, and some Lactobacillus, Bifidobacterium or Bacillus species do not induce a chronic inflammatory response.Citation25–Citation28 In contrast, Enterococcus faecalis or Escherichia coli NC101 trigger severe colitis in Il10-/- mice mono-associated with either strain.Citation27,Citation28 E. coli NC101 is considered to belong to the pathovar of adherent invasive E. coli (AIEC).Citation29 An increased abundance of Enterobacteriaceae, especially of E. coli, has been observed in patients with inflammatory bowel disease and in different animal models of acute and chronic intestinal inflammation.Citation30–Citation35 Specifically AIEC are more prevalent in CD patients than in healthy subjects indicating their involvement in CD development.Citation31,Citation36 Since colitis can be caused by individual bacterial species in the Il10-/- mouse, this model is particularly suitable for investigating the effect of A. muciniphila on intestinal inflammation in a genetically predisposed host.
Effects observed in mono- or dual-associated mice cannot be extrapolated to a complex microbial community as microbe-host and microbe-microbe interactions may play a major role in the development of intestinal inflammation in Il10-/- mice. The simplified human intestinal microbiota (SIHUMI) is a model community that has previously been applied to several mouse models to investigate the role of individual intestinal microbes in disease development under defined and standardized conditions.Citation17,Citation37,Citation38 The SIHUMI consortium consists of Anaerostipes caccae, Bacteroides thetaiotaomicron, Bifidobacterium longum, Blautia producta, Clostridium butyricum, Clostridium ramosum, Escherichia coli and Lactobacillus plantarum.Citation37 The members of the SIHUMI consortium have been selected because of their occurrence in the human gut and their ability to mimic important functions of the human gut microbiota.Citation37
This study aimed to investigate the role of A. muciniphila strain ATCC BAA-835, which was isolated from human feces,Citation4 in promoting short-term intestinal inflammation in Il10-/- mice associated with this organism either alone or in combination with colitogenic E. coli NC101 or with a simplified human intestinal microbiota.
Results
Mono-association of IL-10-deficient mice with A. muciniphila does not trigger intestinal inflammation within three weeks after its application
To investigate whether A. muciniphila alone is capable of inducing inflammation in colitis-prone animals within a short period of time, germ-free Il10-/- mice were mono-associated with A. muciniphila. Bacterial cells numbers reached 6 × 109 cells per gram feces at day six after colonization. Thereafter cell numbers remained unchanged until the mice were euthanized (Figure S1A). A. muciniphila cells were detected in all sections of the intestinal tract with cell numbers ranging from 3 × 107 cells per gram content in duodenum to 6 × 109 cells per gram content in cecum and colon (Figure S2).
Within three weeks after colonization the body-weight development of Il10-/- mice mono-associated with A. muciniphila was similar to that of germ-free Il10-/- mice even though the weight gain of the former was higher at day 21 (). Histopathological scoring of cecal and colonic tissues revealed no significant differences between germ-free Il10-/- mice and Il10-/- mice mono-associated with A. muciniphila (, , , S3A and S3B). Shortening of the colon length, which is indicative of the severity of inflammation in this part of the intestine,Citation39 was not observed either (). The mRNA levels of the pro-inflammatory cytokines tumor necrosis factor alpha (Tnfa), interferon gamma (Ifng), and the antibacterial defense marker regenerating islet-derived protein 3 gamma (Reg3g) were determined as markers for inflammation in the mono-associated mice, but they were not significantly increased in the mucosa of distal small intestine, cecum and colon in comparison to germ-free Il10-/- mice (). Fecal lipocalin-2 is indicative of colitis but also a sensitive marker of mild immune-cell infiltration and low-grade inflammation in Il10-/- mice.Citation40 Fecal lipocalin-2 concentrations were slightly elevated in Il10-/- mice mono-associated with A. muciniphila in comparison to germ-free Il10-/- mice (). Altogether, these data indicate that the bacterial colonization of germ-free Il10-/- mice induced a mild immune response, but A. muciniphila did not cause intestinal inflammation within three weeks after colonization.
Figure 1. Experimental design.
(A) Germ-free Il10-/- mice (n = 6) were compared with Il10-/- mice colonized with A. muciniphila at the age of 8 weeks (n = 6). Three weeks after start of the experiment the mice were euthanized. (B) Germ-free lL-10-/- mice were colonized with E. coli NC101 at the age of 9 weeks (n = 6) and compared with mice additionally colonized with A. muciniphila one week before E. coli NC101 colonization (n = 6). Two weeks after E. coli NC101 colonization the mice were euthanized. (C) Il10-/- mice colonized with the SIHUMI consortium (n = 6) were compared to Il10-/- mice, which were additionally colonized with A. muciniphila at the age of 8 weeks (n = 6). Three weeks after start of the experiment the mice were euthanized.
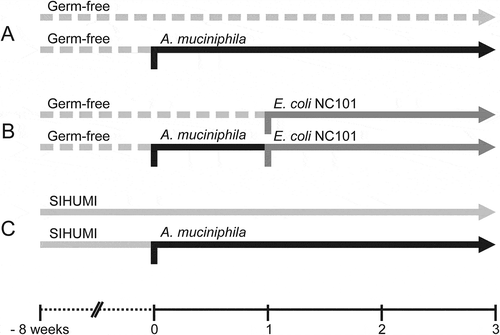
Figure 2. Mono-association of Il10-/- mice with A. muciniphila did not induce intestinal inflammation within three weeks.
(A) Body-weight development of A. muciniphila (Amuc) mono-associated Il10-/- mice within 3 weeks after colonization in comparison to germ-free (GF) Il10-/- mice. Mean ± SD, n = 6, *p < 0.05. (B) Cecal and colonic histopathology score of germ-free (GF) Il10-/- mice and Il10-/- mice mono-associated with A. muciniphila (Amuc). Horizontal dotted line: Maximal score. Median + range, n = 5–6, *p < 0.05. (C) Colon length of germ-free (GF) Il10-/- mice and Il10-/- mice mono-associated with A. muciniphila (Amuc). Mean ± SD, n = 5–6, *p < 0.05. (D) Relative mRNA levels of Tnfa, Ifng and Reg3g in mucosa of distal small intestine (SI), cecum and colon of Il10-/- mice mono-associated with A. muciniphila (Amuc) in comparison to germ-free (GF) Il10-/- mice. Mean ± SD, n = 3–4, *p < 0.05. (E) Fecal lipocalin-2 concentration of germ-free (GF) Il10-/- mice and Il10-/- mice mono-associated with A. muciniphila (Amuc). Median, n = 4–6, *p < 0.05, **p < 0.01.
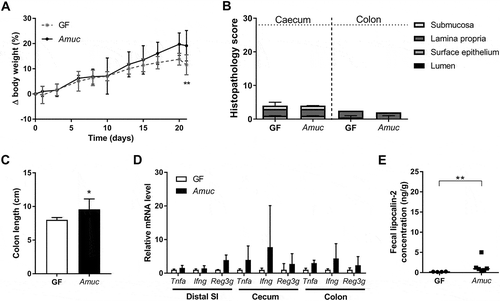
Figure 3. A. muciniphila does not modulate intestinal inflammation triggered by E. coli NC101 in Il10-/- mice.
(A) Body-weight development of originally germ-free and A. muciniphila (Amuc) mono-associated Il10-/- mice additionally colonized with E. coli NC101 (Ecoli) at day 7 (Vertical dotted line) in comparison to germ-free (GF) Il10-/- mice. Mean ± SD, n = 6, *p < 0.05, **p < 0.01, ***p < 0.001 compared to GF, #p < 0.05 for comparison between colonized Il10-/- mice. (B) Cecal and colonic histopathology score of germ-free (GF) Il10-/- mice, Il10-/- mice mono-associated with E. coli NC101 (Ecoli) and Il10-/- mice dual-associated with A. muciniphila and E. coli NC101 (Amuc + Ecoli). Horizontal dotted line: Maximal score. Median + range, n = 5–6, *p < 0.05. (C) Colon length of germ-free (GF) Il10-/- mice, Il10-/- mice mono-associated with E. coli NC101 (Ecoli) and Il10-/- mice dual-associated with A. muciniphila and E. coli NC101 (Amuc + Ecoli). Mean ± SD, n = 5–6, *p < 0.05. (D) Relative mRNA levels of Tnfa, Ifng and Reg3g in mucosa of distal small intestine (SI), cecum and colon of E. coli NC101 mono-associated Il10-/- mice and Il10-/- mice dual-associated with A. muciniphila and E. coli NC101 (Amuc + Ecoli) in comparison to germ-free (GF) Il10-/- mice. Mean ± SD, n = 4–6, *p < 0.05, **p < 0.01. (E) Fecal lipocalin-2 concentration of germ-free (GF) Il10-/- mice, Il10-/- mice mono-associated with E. coli NC101 (Ecoli) and Il10-/- mice dual-associated with A. muciniphila and E. coli NC101 (Amuc + Ecoli). Median, n = 4–6, *p < 0.05.
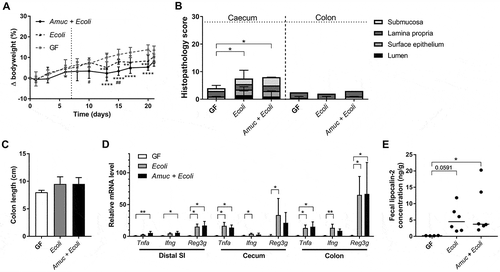
Figure 4. Colonization with A. muciniphila has no short-term effect on the colitogenic activity of the SIHUMI consortium in Il10-/- mice.
(A) Body-weight development of Il10-/- mice born with the SIHUMI consortium (SIHUMI) and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc) in comparison to germ-free (GF) Il10-/- mice. Mean ± SD, n = 6, *p < 0.05 compared to GF, #p < 0.05, ##p < 0.01 for comparison between colonized Il10-/- mice. (B) Cecal and colonic histopathology score of germ-free (GF) Il10-/- mice, SIHUMI-Il10-/- mice (SIHUMI) and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc). Horizontal dotted line: Maximal score. Median + range, n = 5–6, *p < 0.05, **p < 0.01. (C) Colon length of germ-free (GF) Il10-/- mice, SIHUMI-Il10-/- mice (SIHUMI) and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc). Mean ± SD, n = 5–6, *p < 0.05. (D) Relative mRNA levels of Tnfa, Ifng and Reg3g in mucosa of distal small intestine (SI), cecum and colon of SIHUMI-Il10-/- mice (SIHUMI) and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc) in comparison to germ-free (GF) Il10-/- mice. Mean ± SD, n = 4–6, *p < 0.05, **p < 0.01. (E) Fecal lipocalin-2 concentration of germ-free (GF) Il10-/- mice, SIHUMI-Il10-/- mice (SIHUMI) and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc). Median, n = 4–6, *p < 0.05.
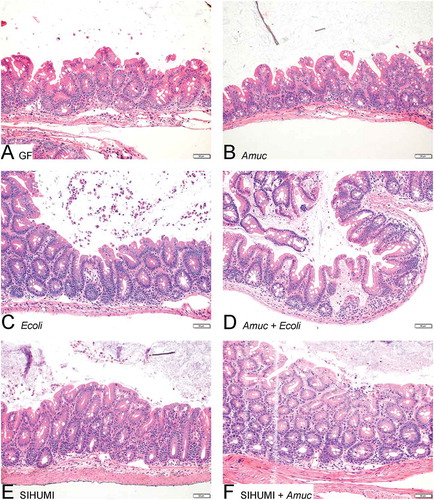
Figure 5. The cecal histology of gnotobiotic Il10-/- mice is not affected by A. muciniphila.
Histology of haematoxylin and eosin-stained cecal tissue sections from (A) germ-free (GF) Il10-/- mice, (B) Il10-/- mice mono-associated with A. muciniphila (Amuc) or (C) E. coli NC101 (Ecoli), (D) Il10-/- mice colonized with E. coli NC101 after pre-colonization with A. muciniphila (Amuc + Ecoli), (E) SIHUMI-Il10-/- mice (SIHUMI) and (F) SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc). Bar: 50 μm.
A. muciniphila does not influence the short-term colitogenic effect of E. coli NC101 in IL-10-deficient mice
Since A. muciniphila did not induce colitis in mono-associated Il10-/- mice, we subsequently investigated whether A. muciniphila promotes short-term intestinal inflammation induced by a colitogenic bacterium. Therefore, one group of germ-free Il10-/- mice was first associated with A. muciniphila and seven days later additionally with colitogenic E. coli NC101, while germ-free Il10-/- control mice were mono-associated with E. coli NC101 at the same time (). There were no significant differences detected with respect to colonization density of E. coli NC101 between mono- and dual-associated Il10-/- mice (Figures S1, S2). A. muciniphila cell numbers were not affected in small intestine and cecum but slightly reduced in colon following the colonization with E. coli NC101. In general, A. muciniphila reached a higher bacterial cell density in cecum and colon than E. coli NC101 in dual-associated Il10-/- mice (Figure S2).
Il10-/- mice mono-associated with E. coli NC101 and Il10-/- mice dual-associated with A. muciniphila and E. coli NC101 gained less body weight than germ-free Il10-/- mice starting six days after colonization with E. coli NC101. Apart from days three and eight after colonization with E. coli NC101, the body-weight gain of both colonized groups was similar (). Mono-association with E. coli NC101 caused cecal inflammation in our Il10-/- mice within two weeks as evident from the histopathological scoring (). Signs of inflammation in cecal tissue of Il10-/- mice mono-associated with E. coli NC101 included the presence of luminal necrotic epithelial cells, epithelial desquamation, an increased abundance of submucosal lymphocytes, and the development of edema. (). However, colon length and colonic histology were not affected by colonization with E. coli NC101 (, and S3C). Pre-colonization with A. muciniphila did not influence the intestinal pathology in mice associated with E. coli NC101 in any way (, and S3D). Il10-/- mice mono-associated with E. coli NC101 showed higher mRNA levels of the inflammatory markers Tnfa, Ifng and Reg3g in mucosa of cecum and colon, and of Reg3g in mucosa of the distal small intestine in comparison with germ-free Il10-/- mice (). Fecal lipocalin-2 concentrations tended to be elevated in Il10-/- mice mono-associated with E. coli NC101 (). Taken together, the data confirm that the mono-association of Il10-/- mice with E. coli NC101 induced intestinal inflammation within two weeks, as demonstrated previously.Citation27,Citation41 Importantly, pre-colonization with A. muciniphila did not result in significant differences in the mRNA levels of Tnfa, Ifng and Reg3g in cecal and colonic mucosa and in fecal lipocalin-2 concentrations of the dual-associated Il10-/- mice compared to the mice associated with E. coli NC101 only (, ). These results demonstrate that A. muciniphila did not modulate the short-term colitogenic activity of E. coli NC101 in Il10-/- mice.
The SIHUMI consortium causes inflammatory changes in IL-10-deficient mice
To investigate the effect of A. muciniphila on the pathogenesis of chronic intestinal inflammation in Il10-/- mice colonized with the SIHUMI consortium, we first examined SIHUMI-Il10-/- mice for signs of intestinal inflammation in the absence of A. muciniphila. The body-weight development of these mice in the time period extending from eight to eleven weeks of age was similar to that of germ-free Il10-/- mice (). At eleven weeks of age, the cecum of the SIHUMI-Il10-/- mice had a significantly increased histopathology score in comparison to the germ-free Il10-/- mice ( and ), while the colon did not show signs of inflammation based on length and histopathological score (, and S3E). No significant differences in mRNA levels of Tnfa, Ifng and Reg3g were detected in mucosa of distal small intestine, cecum and colon between Il10-/- mice associated with the SIHUMI and germ-free Il10-/- mice (). Fecal lipocalin-2 concentrations tended to be elevated in Il10-/- mice colonized with the SIHUMI in comparison to germ-free Il10-/- mice ().
Colonization with A. muciniphila has no short-term impact on the colitogenic activity of the SIHUMI consortium in IL-10-deficient gnotobiotic mice
To assess the short-term effect of A. muciniphila on chronic intestinal inflammation developing in SIHUMI-Il10-/- mice, SIHUMI-Il10-/- mice were additionally colonized with A. muciniphila at the age of eight weeks and analyzed three weeks later (). The colonization of SIHUMI-Il10-/- mice with A. muciniphila did not significantly affect total bacterial cell numbers in cecal contents or those of the individual community members with the exception of B. producta, whose cell number increased slightly (). Cell numbers of L. plantarum were below the detection limit. The cell number of A. muciniphila in the presence of the SIHUMI consortium was approximately 7 × 109 cells per gram of cecal content, which is in the same range as that determined in Il10-/- mice mono-associated with A. muciniphila.
Table 1. Bacterial cell numbers in cecal contents of gnotobiotic Il10-/- mice.
SIHUMI-Il10-/- mice and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila had for the most part a similar body-weight development during the three weeks of co-colonization (). The histopathology scores for cecal tissue of Il10-/- mice associated with the SIHUMI were not affected by the co-colonization with A. muciniphila (). The additional colonization of SIHUMI-Il10-/- mice with A. muciniphila neither reduced the colon length () nor induced histological signs of inflammation in the colonic tissue ( and S3F). Furthermore, the colonization with A. muciniphila had no effect on mRNA levels of the inflammatory markers Tnfa and Ifng in cecal and colonic mucosa of SIHUMI-Il10-/- mice, but the levels of ileal Reg3g were significantly higher in these mice than in SIHUMI-Il10-/- mice without A. muciniphila and in germ-free Il10-/- mice (). Fecal lipocalin-2 concentrations were similar in SIHUMI-Il10-/- mice no matter whether A. muciniphila was present or not (). The data suggest that A. muciniphila did neither promote nor attenuate chronic intestinal inflammation induced by the SIHUMI consortium in the Il10-/- mice within three weeks of co-colonization.
The mucus turnover in IL-10-deficient mice is not influenced by A. muciniphila, E. coli NC101 or the SIHUMI consortium
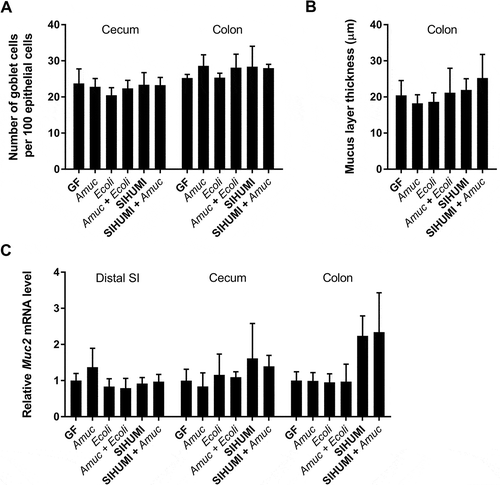
A. muciniphila is known to degrade mucins,Citation4 which form a protective layer and represent a barrier between luminal antigens and the host epithelium. We investigated whether A. muciniphila affected the mucus layer and mucus production in gnotobiotic Il10-/- mice. The proportion of acidic mucin-filled goblet cells relative to the total number of cells forming the epithelial layer () and also the mRNA levels of the mucin 2-encoding Muc2 gene () were similar in cecum and colon of all investigated mouse groups. No reduction in the thickness of the colonic mucus layer was detected following colonization of the gnotobiotic Il10-/- mice with A. muciniphila () suggesting that the mucus turnover in Il10-/- mice was not influenced by A. muciniphila, E. coli NC101 or the SIHUMI consortium.
Figure 6. The mucus turnover in gnotobiotic Il10-/- mice is not influenced by A. muciniphila.
(A) Ratio of acidic mucin filled goblet cells in the cecal and colonic epithelium, (B) thickness of the colonic mucus layer and (C) relative mRNA levels of Muc2 in mucosa of distal small intestine (SI), cecum and colon of germ-free (GF) Il10-/- mice, Il10-/- mice mono-associated with A. muciniphila (Amuc) or E. coli NC101 (Ecoli), Il10-/- mice colonized with E. coli NC101 after pre-colonization with A. muciniphila (Amuc + Ecoli), SIHUMI-Il10-/- mice (SIHUMI) and SIHUMI-Il10-/- mice additionally colonized with A. muciniphila (SIHUMI + Amuc). Mean ± SD, n = 3–6, *p < 0.05.
Discussion
On the one hand A. muciniphila has previously been reported to promote metabolic health in mice fed a high-fat diet,Citation12–Citation15 on the other hand there are reports about the ability of this bacterium to exacerbate intestinal inflammation in murine models of acute colitis.Citation17,Citation18,Citation42 The beneficial effects of A. muciniphila in mouse models for metabolic diseases have prompted suggestions to apply this organism to individuals who suffer from such diseases. However, in view of the above reports about A. muciniphila‘s ability to promote gut inflammation, it is of importance to clarify under which circumstances such negative effects are of relevance. Therefore, in this study we analyzed whether A. muciniphila affects the inflammatory status in a murine model, which is genetically predisposed to the development of chronic colitis, namely the Il10-/- mouse. In our study, which investigated the short-term pro-inflammatory potential of A. muciniphila, the association of gnotobiotic Il10-/- mice with A. muciniphila strain ATCC BAA-835, a human fecal isolate, did not induce or exacerbate intestinal inflammation, no matter whether the mice were germfree, associated with the colitogenic E. coli strain NC101 or associated with a simplified human intestinal microbiota.
Mono-association of Il10-/- mice with A. muciniphila did not induce signs of intestinal inflammation within three weeks but only mildly activated the immune response based on slightly increased fecal lipocalin-2 concentrations. This result indicates that A. muciniphila did not disturb gut immune homeostasis but was perceived as a non-inflammatory, commensal bacterium by the host immune system as observed for wild type mice,Citation43 even if the host, the Il10-/- mouse, was genetically predisposed to developing colitis.Citation20 Indeed, mono-association of NMRI wild type mice with A. muciniphila induces the expression of genes involved in antigen presentation in cecum and upregulates the expression of genes involved in innate and adaptive immune responses in the colon without inducing a pro-inflammatory response in these mice.Citation43 Recently, A. muciniphila was reported to trigger intestinal inflammation in germ-free Il10-/- mice within eight weeks following colonization and to increase colitis in specific-pathogen-free (SPF) Il10-/- mice.Citation42
In our study we used the type strain of A. muciniphila, which was isolated from human feces,Citation4 while Ref. Citation42 used a newly isolated murine A. muciniphila strain. Different bacterial isolates of the same species can differ in more than 1% of their genomes but are still assigned to the same species. The authors of Ref. Citation11 addressed this point by analyzing 39 isolates of A. muciniphila derived from mice and humans and identified three phylogroups within this species. The genetic diversity of A. muciniphila isolates is higher in the phylogroup that preferentially occurs in humans in comparison to the phylogroup that occurs in both humans and mice. The genetic diversity between the phylogroups is reflected by a broader repertoire of substrate metabolizing enzymes in the former phylogroup.Citation11 However, the consequences of genetic diversity in the context of specific effects and features of A. muciniphila still require further investigation. A direct comparison of different A. muciniphila strains regarding their colitogenic potential would be an interesting new aspect of the underlying research question but is beyond the scope of this investigation.
Even though A. muciniphila has been shown to respond to different diets, for example obesity-inducing high-fat diet,Citation12 and specific dietary supplements such as polyphenols,Citation14 we suppose that diet plays only a minor role in explaining the discrepant results of both studies, because no specific experimental diets were fed.
In the present study the cell numbers of A. muciniphila detected in the feces of the Il10-/- mice exceeded 109 cells/g. These cell numbers are approximately one order of magnitude higher than those detected in humans but in the same range as those reported for mono-associated wild-type mice.Citation2,Citation43 This suggests that the lack of a pro-inflammatory effect of A. muciniphila is not due to an insufficient colonization efficiency of the A. muciniphila strain used in our experiments. It is important to note that we analyzed the short-term effect of A. muciniphila strain ATCC BAA-835 three weeks after its application. Therefore, we cannot rule out that intestinal inflammation might have developed at a later point in time.
The administration of A. muciniphila to DSS-treated mice aggravates intestinal inflammation, whereas A. muciniphila-derived extracellular vesicles have the opposite effect.Citation18 In contrast, pre-treatment of mice with A. muciniphila before DSS administration exerts an anti-inflammatory effect, which is reproducible in rats that harbor a human fecal microbiota.Citation44 Furthermore, the administration of chlorogenic acid, a polyphenolic compound, which promotes the growth of A. muciniphila, before and during DSS treatment attenuates colitis.Citation45 In the present study, A. muciniphila had neither a preventive effect on colitis induced by E. coli NC101 nor could the inflammatory symptoms induced by the SIHUMI be improved by treatment with A. muciniphila. The underlying mechanisms explaining the varying effects of A. muciniphila on intestinal inflammation in the different models remain to be elucidated.
A. muciniphila did not enhance the cecal inflammation developing in Il10-/- mice within two weeks after colonization with the colitogenic bacterium E. coli NC101. The progression of cecal colitis after mono-association with E. coli NC101 in the Il10-/- C57BL/6 mice is comparable to that observed for Il10-/- 129S6 mice, in which the first histological signs of cecal inflammation were observed three weeks after colonization with E. coli NC101.Citation27,Citation29 A tendency towards an increased expression of inflammatory markers and a slightly increased cecal histopathology score three weeks after mono-association with E. coli NC101 were also reported for Il10-/- C3H mice.Citation46 The colitis developing within three weeks after colonization with E. coli NC101 was described as mild or moderate in case of strain 129S6.Citation27,Citation29 In C3H mice, the signs of mild colitis detected after eight weeks were not yet significant after three weeks.Citation46 In conclusion, the colitogenic effect of E. coli NC101 in Il10-/- C57BL/6 mice observed in the present study is in accordance with previous publications.
Similar fecal and intestinal cell numbers of A. muciniphila and E. coli NC101, in both dual-associated and mono-associated Il10-/- mice suggest that these two species neither compete for the same ecological niche in the intestine nor influence each other’s niche. This may be one reason why A. muciniphila influenced the acute inflammatory response of wildtype mice harboring a simplified human intestinal microbiota to Salmonella enterica Typhimurium infection,Citation17 but not the chronic inflammatory response of Il10-/- mice to E. coli NC101. Another reason probably lies in the fact that the pathogenic mechanisms underlying the pro-inflammatory effects of Salmonella enterica Typhimurium in wildtype mice and of E. coli NC101 in Il10-/- mice differ considerably.
In view of its immuno- and mucus-modulating properties,Citation4,Citation43 A. muciniphila was originally hypothesized to be able to alter the susceptibility of previously germ-free Il10-/- mice to the colitogenic E. coli NC101. The finding that the utilization of mucin by A. muciniphila in vivo seems to be dependent on the presence of other bacteria suggests that A. muciniphila can only exploit its full potential within a microbial community.Citation47 Therefore, we investigated whether A. muciniphila promotes intestinal inflammation in Il10-/- mice harboring the same SIHUMI consortium that was used in the Salmonella-infection model,Citation17 but there were no indications that A. muciniphila promoted inflammation in SIHUMI-Il10-/- mice within three weeks after its application. In contrast to the Il10-/- mice, in which inflammation was induced by E. coli NC101, the SIHUMI-Il10-/- mice showed histological changes in cecal tissue but no elevated mRNA levels of the inflammatory markers Tnfa, Ifng, and Reg3g, and the fecal lipocalin-2 concentrations remained very low. While the colitogenic effect of E. coli NC101 was analyzed two weeks after administration of the bacterium, the Il10-/- mice harboring the SIHUMI consortium from birth on were analyzed at eleven weeks of age. There might have been an initial innate immune response that was already attenuated at the time point of analysis. This view is supported by the fact that bacterial colonization of Il10-/- mice is accompanied by initially high levels of IFNγ in cecum and colon, which later on are down-regulated although they remain persistently higher than in wild type mice.Citation48 Furthermore, ileal Reg3γ levels increase two weeks after conventionalization of germ-free mice and thereafter return to basal levels.Citation49
Administration of A. muciniphila to SIHUMI-Il10-/- mice did not influence the composition of the SIHUMI consortium. Similarly, no major changes in the microbiota profile were observed following the oral administration of A. muciniphila to conventional mice.Citation12 This may be explained by the fact that A. muciniphila is a late colonizer of the human gut compared to E. coli, Lactobacillus, Clostridium and Bifidobacterium,Citation2,Citation50,Citation51 but still finds its niche due to its ability to use mucus as carbon and nitrogen source.Citation4 A simplified human microbiota consortium also named SIHUMI has previously been reported to induce colitis in Il10-/- mice.Citation52 However, this consortium differs in species composition. It includes for example Enterococcus faecalis, which has previously been demonstrated to induce colonic inflammation in Il10-/- mice mono-associated with this organism.Citation27,Citation28 Some members of our SIHUMI consortium, Bacteroides thetaiotaomicron and Bifidobacterium longum, are known to modulate the host immune response.Citation53 Effects described for E. coli are strain-dependent. E. coli Nissle, in contrast to E. coli NC101, does not induce the expression of inflammatory markers in Il10-/- mice.Citation46 Because of the low expression of inflammatory markers in our SIHUMI-Il10-/- mice it is tempting to speculate that Escherichia coli K-12 MG1655, which is a member of the SIHUMI consortium, resembles E. coli Nissle in that it is not pro-inflammatory in Il10-/- mice.
In the DSS- or Salmonella-induced colitis models, pretreatment of mice with A. muciniphila or growth stimulation of endogenous A. muciniphila two weeks or less prior to colitis induction was sufficient to detect the inflammation-modulating effects of this organism.Citation17,Citation18,Citation44,Citation45 For example, disturbance of mucus homeostasis in gnotobiotic mice was already observed two weeks after colonization with A. muciniphila.Citation17 The authors of this study proposed that the aggravation of the inflammation observed in response to colonization of the mice with A. muciniphila was due to the weakening of the intestinal barrier.Citation17 In the present study, A. muciniphila did not affect the mucus layer or promote colitis development in the gnotobiotic Il10-/- mice within a similar period of time. By using Il10-/- mice mono-associated with E. coli NC101 as a control group, we confirm that intestinal inflammation in Il10-/- mice can be induced by a single gut bacterium such as E. coli NC101 within two weeks.
The mucus layer of Il10-/- mice is more penetrable by bacteria compared to that of wild type mice although there is no difference in total thickness of the mucus layer with the inner adherent layer being even thicker in the Il10-/- mice.Citation54 We tested whether the mucus degrader A. muciniphila influences mucus production and mucus layer thickness in our Il10-/- mice, but there were no indications that A. muciniphila affected mucus production in any of the gnotobiotic Il10-/- mice investigated in our study. A reduction of the mucus layer thickness without changes in mucus production in response to an increased colonization with A. muciniphila was reported for SPF Il10-/- mice.Citation42 No such an effect of A. muciniphila could be seen in our study because the gnotobiotic Il10-/- mice displayed a relatively thin mucus layer already in the absence of A. muciniphila. Although A. muciniphila is known to degrade mucus,Citation4 it is capable of restoring the mucus layer after its depletion caused by feeding a high-fat diet.Citation12 The complex relationship between A. muciniphila and the intestinal mucus layer is still obscure and needs further clarification.
In summary, this study did not find any indication that A. muciniphila strain ATCC BAA-835, which was isolated from human feces, induces or promotes chronic intestinal inflammation in gnotobiotic mice genetically predisposed to colitis within two to three weeks. Our study indicates that the link of A. muciniphila with intestinal inflammation in animal models is not an intrinsic feature of this organism but is only observed under very specific conditions, which still need to be defined more precisely.
Owing to the reduction of body weight and the improvement of metabolic parameters observed in obese mice in response to the oral uptake of A. muciniphila it has been proposed to use this organism for the treatment of human metabolic disorders.Citation12,Citation15,Citation16,Citation55 The association of a higher abundance of A. muciniphila with normal body weight and a healthier metabolic status of humans,Citation8,Citation11 the more effective body-weight reduction and metabolic improvement after caloric restriction in overweight patients harboring higher numbers of A. muciniphila,Citation9 together with first clinical trials that did not detect any side effects of orally applied A. muciniphila support this objective.Citation15,Citation56 The results of our study do not argue against the use of A. muciniphila for the prevention or treatment of metabolic disorders.
Materials and methods
Bacterial strains and culture conditions
Akkermansia muciniphila ATCC BAA-835 and Escherichia coli NC101 were grown in Yeast Hemin Brain-Heart-Infusion Broth (YH-BHI; BHI supplemented with 5 g/l yeast extract and 5 mg/l hemin) at 37°C under strictly anaerobic conditions with N2/CO2 (80/20, v/v) as a gas phase. For colonization of mice, bacteria were harvested under anaerobic conditions in the exponential growth phase. Bacterial cell numbers were calculated from OD600 measurements.
Animals and experimental design
Male C57BL/6.129P2-Il10tm1Cgn offspring from germ-free mice and mice harboring a simplified human intestinal microbiota (SIHUMI) consisting of Anaerostipes caccae DSM 14662, Bacteroides thetaiotaomicron DSM 2079, Bifidobacterium longum NCC 2705, Blautia producta DSM 2950, Clostridium butyricum DSM 10702, Clostridium ramosum DSM 1402, Escherichia coli K-12 MG1655 and Lactobacillus plantarum DSM 20174 were used.Citation37 Successful colonization of mice with SIHUMI was confirmed in feces by fluorescence in situ hybridization (FISH) (except Lactobacillus, which was below the detection limit in feces samples). The mutation of the IL-10 gene was verified by genotyping. Mice were kept in positive-pressure isolators (Metall & Plastik, Radolfzell, Germany) in the gnotobiotic animal facility of the German Institute of Human Nutrition, Potsdam-Rehbruecke, Germany, on irradiated wood chips (25–50 kGy) with free access to irradiated standard chow (Altromin fortified type 1310; Altromin, Lage, Germany) and autoclaved water in a 12 h light-dark cycle, at 22 ± 2°C and 55 ± 5% air humidity. A. muciniphila or E. coli NC101 were administered to mice by gastric gavage of 108 cells. The animal experiments were approved by the Animal Welfare Committee of the State of Brandenburg (Approval no. 2347–11-2014 + T-03–15-GAMI).
Experiment A Germ-free Il10-/- mice (n = 6) were colonized with A. muciniphila at the age of 8 weeks. After determination of body weight and feces collection for three weeks, mice were euthanized. Germ-free Il10-/- mice (n = 6) of the same age served as control group ().
Experiment B Germ-free Il10-/- mice (n = 6) were first colonized with A. muciniphila at 8 weeks of age and one week later with E. coli NC101. The corresponding control Il10-/- mice (n = 6) were mono-associated with E. coli NC101 at the same time point. Body weight was determined and fresh feces were collected until the end of the experiment, i.e. three weeks after the first intervention ().
Experiment C SIHUMI-Il10-/- mice (n = 6) were additionally colonized with A. muciniphila at 8 weeks of age and compared to untreated SIHUMI- Il10-/- mice (n = 6). After determining body weight and collecting feces for three weeks, mice were euthanized ().
The gut was divided into proximal, middle and distal small intestine, cecum and colon. Tissue samples from each part were fixed in Carnoy’s solution and both mucosa and contents were collected. Mucosa was immediately frozen in liquid nitrogen and stored at -80°C until use. Bacteria in fresh cecal content were immediately fixed for bacterial enumeration by FISH.
Fluorescence in situ hybridization and quantification of bacterial cells
Bacteria in feces or gut contents were enumerated by FISH using fluorescently labeled oligonucleotide probes targeting the 16S rRNA of the bacteria. Fresh feces or gut contents were diluted ten-fold with phosphate-buffered saline (PBS; 137 mM NaCl, 10 mM Na2HPO4, 2 mM NaH2PO4; pH 7.4) and homogenized. After centrifugation to remove debris (300 × g, 1 min, 4 °C) the supernatant was diluted with two volumes of 4% paraformaldehyde and incubated for 1 h at 4°C. After washing with PBS (8000 × g, 5 min, 4°C) cells were suspended in PBS/ethanol (1:1, v/v) and stored at -20°C until analysis.
Bacterial cells were quantified by FISH as described in Ref. Citation57. Lysozyme treatment was only used for the enumeration of gram-positive bacteria. Hybridization was done overnight with 1 µl probe solution (10 pM, Eub probes 1.67 pM) and 10 µl hybridization buffer (900 mM NaCl, 10 mM TrisHCl pH 7.4, 0.01% SDS) per well except for Bac303, which was hybridized for 2 h, at the hybridization conditions indicated for each probe. The oligonucleotide probes used in this study were labeled with Cyanine-3 at their 5ʹ end, except MUC-1437 (50°C, 20% formamide), which was labeled at the 5ʹ and the 3ʹ end.Citation3 The probes used for detection of SIHUMI-bacteria were Acac0194 (46°C), Bac303 (46°C), Bif164 (54°C), ProCo1264 (35°C, 30% formamide), Cbut183 (35°C, 30% formamide), Cra757 (35°C, 30% formamide), ECOLI (50°C) and Lab158 (46°C).Citation37 An equimolar mixture of the Eub probes (Eub338, Eub785, Eub927, Eub1055, Eub1088, Eub338Akk (5ʹ- GCT GCC ACC CGT AGG TGT-3‘)) was used to determine total bacterial cell counts.Citation57–Citation60
Measurement of mRNA levels of inflammatory markers
To quantify relative mRNA levels of the murine inflammatory markers interferon gamma (Ifng, NM_008337.4, forward primer 5ʹ-CAG GCC ATC AGC AAC AAC ATA AGC G-3ʹ, reverse primer 5ʹ-TGG ACC TGT GGG TTG TTG ACC TCA-3ʹ), tumor necrosis factor alpha (Tnfa, NM_013693.3, forward primer 5ʹ- GAC CCT CAC ACT CAG ATC ATC TTC T-3ʹ, reverse primer 5ʹ- CCA CTT GGT GGT TTG CTA CGA-3ʹ) and regenerating islet-derived protein 3 gamma (Reg3g, NM_011260.2, forward primer 5- TCC ATG ACC CGA CAC TGG GCT-3ʹ, reverse primer 5ʹ- TGC CTG AGG AAG AGG AAG GAT TCG T-3ʹ) and also of mucin 2 (Muc2, NM_023566.3, forward primer 5ʹ- CTG ACC AAG AGC GAA CAC AA-3ʹ, reverse primer 5ʹ-CAT GAC TGG AAG CAA CTG GA-3ʹ), RNA was extracted from intestinal mucosa using the peqGOLD TriFast reagent (VWR, 30–2010). Residual genomic DNA was removed with the Turbo DNA-free kit (Thermo Fisher Scientific, AM1907). Elimination of DNA was considered successful, if no amplification of 18S rRNA genes was detectable. Integrity of the purified RNA was checked on agarose gels stained with ethidium bromide. RNA was converted to cDNA using the RevertAid H Minus first-strand cDNA synthesis kit (Thermo Fisher Scientific, K1631). Quantitative real-time PCR was carried out using the Viia 7 – Realtime PCR System (Applied Biosystems, Foster City, CA). The reaction mix contained 2.5 µl Power SYBR green PCR master mix (Thermo Fisher Scientific, 4367659), 0.5 µl of forward and reverse primers (3 µM) and 1 µl cDNA sample corresponding to 5 ng of RNA adjusted with water to 5 µl. Relative gene expression was calculated with the threshold cycle (ΔΔCT) method.Citation61 Results were normalized to hypoxanthine guanine phosphoribosyl transferase (Hprt, NM_013556.2, forward primer 5ʹ- CAG TCC CAG CGT CGT GAT TA-3ʹ, reverse primer 5ʹ- AGC AAG TCT TTC AGT CCT GTC-3ʹ) as reference gene and are shown relative to the germ-free group normalized to a value of 1.
Measurement of fecal lipocalin-2 concentrations
To quantify lipocalin-2 in fecal samples collected one day before euthanization of mice, supernatant of frozen feces was obtained as described in Ref. Citation40. The lipocalin-2 concentration in fecal supernatant was determined using the Mouse Lipocalin-2/NGAL Quantikine ELISA Kit (R&D Systems, MLCN20).
Histological scoring
Cecal and colonic tissue samples were fixed in Carnoy’s solution and embedded in paraffin for histological analysis. After staining with haematoxylin and eosin, sections were evaluated by an experienced pathologist in blinded fashion for intestinal inflammation. The following parameters were scoredCitation17: Lumen: necrotic epithelial cells: 1 = scant, 2 = moderate, 3 = dense; neutrophils: 1 = scant, 2 = moderate, 3 = dense. Surface epithelium: villous atrophy and fusion: 1 = scant, 2 = moderate, 3 = dense; epithelial hyperplasia: 0 = none, 1 = present; desquamation: 1 = patchy (<30%), 2 = diffuse (>30%); ulceration: 0 = no, 1 = present; crypt abscesses: 0 = none, 1 = rare, 2 = moderate, 3 = abundant. Lamina propria: infiltration: 0 = minimal, 1 = mild, 2 = moderate, 3 = abundant; lymphocytes: 0 = very few, 1 = some, 2 = numerous. Submucosa: lymphocytes: 0 = very few, 1 = some, 2 = numerous; polymorph nuclear cells (PMNs): 0 = none, 1 = few, 2 = many; edema: 0 = none to mild, <10% of the mucosa, 1 = moderate, 2 = severe; Peyer patch hyperplasia: 0 = none, 1 = present.
Quantification of mucin-filled goblet cells and mucus layer thickness
After fixation in Carnoy’s solution, cecal and colonic tissue samples were embedded in paraffin for histological analysis. Acidic mucins were stained with alcian blue at pH 2.5 and cell nuclei with haematoxylin. Mucin-filled goblet cells and total cell nuclei in the epithelium were enumerated and mucus layer thickness was determined using an Axioplan2 Imaging Microscope (Carl Zeiss) with a Plan-Neofluar 40x objective.
Statistical analysis
Data are presented as means with standard deviation (SD) if not indicated otherwise. Differences are considered significant, if p < 0.05. Comparison of two groups was done using the Mann Whitney test. More than two groups were compared using the Kruskal-Wallis test with Dunn’s multiple comparisons test. Body-weight change over time was compared using 2way RM ANOVA followed by Sidak’s multiple comparisons test in case of two groups or followed by Tukey’s multiple comparisons test in case of three groups. Statistical analysis was performed using GraphPad Prism version 6.07 (GraphPad Software, La Jolla, CA).
Supplemental Material
Download MS Word (4.7 MB)Acknowledgments
We thank Lisa Richter for taking care of the animals. We acknowledge Hannah-Maria Ebert, Meike Braun, Sabine Schmidt, Marion Urbich and Elisabeth Meyer for their technical assistance.
Disclosure statement
The authors have no conflicts of interest to disclose.
Supplementary data
Supplemental data for this article can be accessed here
Additional information
Funding
References
- Blaut M, Clavel T. Metabolic diversity of the intestinal microbiota: implications for health and disease. J Nutr. 2007;137(3):751S–755S. doi:10.1093/jn/137.3.751S.
- Collado MC, Derrien M, Isolauri E, de Vos WM, Salminen S. Intestinal integrity and Akkermansia muciniphila, a mucin-degrading member of the intestinal microbiota present in infants, adults, and the elderly. Appl Environ Microbiol. 2007;73(23):7767–7770. doi:10.1128/AEM.01477-07.
- Derrien M, Collado MC, Ben-Amor K, Salminen S, de Vos WM. The mucin degrader Akkermansia muciniphila is an abundant resident of the human intestinal tract. Appl Environ Microbiol. 2008;74(5):1646–1648. doi:10.1128/AEM.01226-07.
- Derrien M, Vaughan EE, Plugge CM, de Vos WM. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int J Syst Evol Microbiol. 2004;54(5):1469–1476. doi:10.1099/ijs.0.02873-0.
- Derrien M, Belzer C, de Vos WM. Akkermansia muciniphila and its role in regulating host functions. Microb Pathog. 2017;106:171–181. doi:10.1016/j.micpath.2016.02.005.
- Png CW, Lindén SK, Gilshenan KS, Zoetendal EG, McSweeney CS, Sly LI, McGuckin MA, Florin TH. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am J Gastroenterol. 2010;105(11):2420–2428. doi:10.1038/ajg.2010.281.
- Swidsinski A, Dörffel Y, Loening-Baucke V, Theissig F, Rückert JC, Ismail M, Rau WA, Gaschler D, Weizenegger M, Kühn S, et al. Acute appendicitis is characterised by local invasion with Fusobacterium nucleatum/necrophorum. Gut. 2011;60(1):34–40. doi:10.1136/gut.2009.191320.
- Karlsson CL, Önnerfält J, Xu J, Molin G, Ahrné S, Thorngren-Jerneck K. The microbiota of the gut in preschool children with normal and excessive body weight. Obesity. 2012;20(11):2257–2261. doi:10.1038/oby.2012.110.
- Dao MC, Everard A, Aron-Wisnewsky J, Sokolovska N, Prifti E, Verger EO, Kayser BD, Levenez F, Chilloux J, Hoyles L, et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut. 2015;65(3):426–436. doi:10.1136/gutjnl-2014-308778.
- Schneeberger M, Everard A, Gómez-Valadés AG, Matamoros S, Ramírez S, Delzenne NM, Gomis R, Claret M, Cani PD. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci Rep. 2015;5:16643. doi:10.1038/srep16643.
- Guo X, Li S, Zhang J, Wu F, Li X, Wu D, Zhang M, Ou Z, Jie Z, Yan Q, et al. Genome sequencing of 39 Akkermansia muciniphila isolates reveals its population structure, genomic and functional diverisity, and global distribution in mammalian gut microbiotas. BMC Genomics. 2017;18(1):800. doi:10.1186/s12864-017-4195-3.
- Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, Guiot Y, Derrien M, Muccioli GG, Delzenne NM, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. 2013;110(22):9066–9071. doi:10.1073/pnas.1219451110.
- Shin NR, Lee JC, Lee HY, Kim MS, Whon TW, Lee MS, Bae JW. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut. 2014;63(5):727–735. doi:10.1136/gutjnl-2012-303839.
- Roopchand DE, Carmody RN, Kuhn P, Moskal K, Rojas-Silva P, Turnbaugh PJ, Raskin I. Dietary polyphenols promote growth of the gut bacterium Akkermansia muciniphila and attenuate high-fat diet-induced metabolic syndrome. Diabetes. 2015;64(8):2847–2858. doi:10.2337/db14-1916.
- Plovier H, Everard A, Druart C, Depommier C, Van Hul M, Geurts L, Chilloux J, Ottman N, Duparc T, Lichtenstein L, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat Med. 2017;23(1):107–113. doi:10.1038/nm.4236.
- Ouwerkerk JP, Aalvink S, Belzer C, de Vos WM. Preparation and preservation of viable Akkermansia muciniphila cells for therapeutic interventions. Benef Microbes. 2017;8(2):163–169. doi:10.3920/BM2016.0096.
- Ganesh BP, Klopfleisch R, Loh G, Blaut M. Commensal Akkermansia muciniphila exacerbates gut inflammation in Salmonella Typhimurium-infected gnotobiotic mice. PloS One. 2013;8(9):e74963. doi:10.1371/journal.pone.0074963.
- Kang CS, Ban M, Choi EJ, Moon HG, Jeon JS, Kim DK, Park SK, Jeon SG, Roh TY, Myung SJ, et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One. 2013;8(10):e76520. doi:10.1371/journal.pone.0076520.
- Weir TL, Manter DK, Sheflin AM, Barnett BA, Heuberger AL, Ryan EP. Stool microbiome and metabolome differences between colorectal cancer patients and healthy adults. PloS One. 2013;8(8):e70803. doi:10.1371/journal.pone.0070803.
- Kühn R, Löhler J, Rennick D, Rajewsky K, Müller W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell. 1993;75(2):263–274. doi:10.1016/0092-8674(93)80068-P.
- Berg DJ, Davidson N, Kühn R, Müller W, Menon S, Holland G, Thompson-Snipes L, Leach MW, Rennick D. Enterocolitis and colon cancer in interleukin-10-deficient mice are associated with aberrant cytokine production and CD4(+) TH1-like responses. J Clin Invest. 1996;98(4):1010–1020. doi:10.1172/JCI118861.
- Büchler G, Wos-Oxley ML, Smoczek A, Zschemisch NH, Neumann D, Pieper DH, Hedrich HJ, Bleich A. Strain-specific colitis susceptibility in IL10-deficient mice depends on complex gut microbiota-host interactions. Inflamm Bowel Dis. 2012;18(5):943–954. doi:10.1002/ibd.21895.
- Sellon RK, Tonkonogy S, Schultz M, Dieleman LA, Grenther W, Balish E, Rennick DM, Sartor RB. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect Immun. 1998;66(11):5224–5231.
- Keubler LM, Buettner M, Häger C, Bleich A. A multihit model: colitis lessons from the Interleukin-10-deficient mouse. Inflamm Bowel Dis. 2015;21(8):1967–1975. doi:10.1097/MIB.0000000000000468.
- Dieleman LA, Arends A, Tonkonogy SL, Goerres MS, Craft DW, Grenther W, Sellon RK, Balish E, Sartor RB. Helicobacter hepaticus does not induce or potentiate colitis in interleukin-10-deficient mice. Infect Immun. 2000;68(9):5107–5113. doi:10.1128/IAI.68.9.5107-5113.2000.
- Sydora BC, Tavernini MM, Doyle JS, Fedorak RN. Association with selected bacteria does not cause enterocolitis in IL-10 gene-deficient mice despite a systemic immune response. Dig Dis Sci. 2005;50(5):905–913. doi:10.1007/s10620-005-2663-0.
- Kim SC, Tonkonogy SL, Albright CA, Tsang J, Balish EJ, Braun J, Huycke MM, Sartor RB. Variable phenotypes of enterocolitis in interleukin 10-deficient mice monoassociated with two different commensal bacteria. Gastroenterology. 2005;128(4):891–906. doi:10.1053/j.gastro.2005.02.009.
- Balish E, Warner T. Enterococcus faecalis induces inflammatory bowel disease in interleukin-10 knockout mice. Am J Pathol. 2002;160(6):2253–2257. doi:10.1016/S0002-9440(10)61172-8.
- Patwa LG, Fan TJ, Tchaptchet S, Liu Y, Lussier YA, Sartor RB, Hansen JJ. Chronic intestinal inflammation induces stress-response genes in commensal Escherichia coli. Gastroenterology. 2011;141(5):1842–1851. doi:10.1053/j.gastro.2011.06.064.
- Kotlowski R, Bernstein CN, Sepehri S, Krause DO. High prevalence of Escherichia coli belonging to the B2+D phylogenetic group in inflammatory bowel disease. Gut. 2007;56(5):669–675. doi:10.1136/gut.2006.099796.
- Baumgart M, Dogan B, Rishniw M, Weitzman G, Bosworth B, Yantiss R, Orsi RH, Wiedmann M, McDonough P, Kim SG, et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 2007;1(5):403–418. doi:10.1038/ismej.2007.52.
- Heimesaat MM, Fischer A, Jahn HK, Niebergall J, Freudenberg M, Blaut M, Liesenfeld O, Schumann RR, Göbel UB, Bereswill S. Exacerbation of murine ileitis by Toll-like receptor 4 mediated sensing of lipopolysaccharide from commensal Escherichia coli. Gut. 2007;56(7):941–948. doi:10.1136/gut.2006.104497.
- Wohlgemuth S, Haller D, Blaut M, Loh G. Reduced microbial diversity and high numbers of one single Escherichia coli strain in the intestine of colitic mice. Environ Microbiol. 2009;11(6):1562–1571. doi:10.1111/j.1462-2920.2009.01883.x.
- Maharshak N, Packey CD, Ellermann M, Manick S, Siddle JP, Huh EY, Plevy S, Sartor RB, Carroll IM. Altered enteric microbiota ecology in interleukin 10-deficient mice during development and progression of intestinal inflammation. Gut Microbes. 2013;4(4):316–324. doi:10.4161/gmic.25486.
- De Fazio L, Cavazza E, Spisni E, Strillacci A, Centanni M, Candela M, Praticò C, Campieri M, Ricci C, Valerii MC. Longitudinal analysis of inflammation and microbiota dynamics in a model of mild chronic dextran sulfate sodium-induced colitis in mice. World J Gastroenterol. 2014;20(8):2051–2061. doi:10.3748/wjg.v20.i8.2051.
- Darfeuille-Michaud A, Boudeau J, Bulois P, Neut C, Glasser AL, Barnich N, Bringer MA, Swidsinski A, Beaugerie L, Colombel JF. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology. 2004;127(2):412–421. doi:10.1053/j.gastro.2004.04.061.
- Becker N, Kunath J, Loh G, Blaut M. Human intestinal microbiota: characterization of a simplified and stable gnotobiotic rat model. Gut Microbes. 2011;2(1):25–33. doi:10.4161/gmic.2.1.14651.
- Woting A, Pfeiffer N, Loh G, Klaus S, Blaut M. Clostridium ramosum promotes high-fat diet-induced obesity in gnotobiotic mouse models. MBio. 2014;5(5):e01530–14. doi:10.1128/mBio.01530-14.
- Oh SY, Cho KA, Kang JL, Kim KH, Woo SY. Comparison of experimental mouse models of inflammatory bowel disease. Int J Mol Med. 2014 Feb;33(2):333–340. doi:10.3892/ijmm.2013.1569.
- Chassaing B, Srinivasan G, Delgado MA, Young AN, Gewirtz AT, Vijay-Kumar M. Fecal lipocalin 2, a sensitive and broadly dynamic non-invasive biomarker for intestinal inflammation. PLoS One. 2012;7(9):e44328. doi:10.1371/journal.pone.0044328.
- Kim SC, Tonkonogy SL, Karrasch T, Jobin C, Sartor RB. Dual-association of gnotobiotic IL-10-/- mice with 2 nonpathogenic commensal bacteria induces aggressive pancolitis. Inflamm Bowel Dis. 2007;13(12):1457–1466. doi:10.1002/ibd.20246.
- Seregin SS, Golovchenko N, Schaf B, Chen J, Pudlo NA, Mitchell J, Baxter NT, Zhao L, Schloss PD, Martens EC, et al. NLRP6 protects Il10-/- mice from colitis by limiting colonization of Akkermansia muciniphila. Cell Rep. 2017;19(10):733–745. doi:10.1016/j.celrep.2017.05.074.
- Derrien M, Van Baarlen P, Hooiveld G, Norin E, Müller M, de Vos WM. Modulation of mucosal immune response, tolerance, and proliferation in mice colonized by the mucin-degrader Akkermansia muciniphila. Front Microbiol. 2011;2:166. doi:10.3389/fmicb.2011.00166.
- Gobert AP, Sagrestani G, Delmas E, Wilson KT, Verriere TG, Dapoigny M, Del’homme C, Bernalier-Donadille A. The human intestinal microbiota of constipated-predominant irritable bowel syndrome patients exhibits anti-inflammatory properties. Sci Rep. 2016;6:39399. doi:10.1038/srep39399.
- Zhang Z, Wu X, Cao S, Cromie M, Shen Y, Feng Y, Yang H, Li L. Chlorogenic acid ameliorates experimental colitis by promoting growth of Akkermansia in mice. Nutrients. 2017;9(7):E677. doi:10.3390/nu9070677.
- Schumann S, Alpert C, Engst W, Klopfleisch R, Loh G, Bleich A, Blaut M. Mild gut inflammation modulates the proteome of intestinal Escherichia coli. Environ Microbiol. 2013;16(9):2966–2979. doi:10.1111/1462-2920.12192.
- Berry D, Stecher B, Schintlmeister A, Reichert J, Brugiroux S, Wild B, Wanek W, Richter A, Rauch I, Decker T, et al. Host-compound foraging by intestinal microbiota revealed by single-cell stable isotope probing. Proc Natl Acad Sci USA. 2013;110(12):4720–4725. doi:10.1073/pnas.1219247110.
- Sydora BC, Tavernini MM, Wessler A, Jewell LD, Fedorak RN. Lack of interleukin-10 leads to intestinal inflammation, independent of the time at which luminal microbial colonization occurs. Inflamm Bowel Dis. 2003;9(2):87–97. doi:10.1097/00054725-200303000-00002.
- Johansson ME, Jakobsson HE, Holmén-Larsson J, Schütte A, Ermund A, Rodríguez-Piñeiro AM, Arike L, Wising C, Svensson F, Bäckhed F, et al. Normalization of host intestinal mucus layers requires long-term microbial colonization. Cell Host Microbe. 2015;18(5):582–592. doi:10.1016/j.chom.2015.10.007.
- Mackie RI, Sghir A, Gaskins HR. Developmental microbial ecology of the neonatal gastrointestinal tract. Am J Clin Nutr. 1999;69(5):1035S–1045S. doi:10.1093/ajcn/69.5.1035s.
- Favier CF, Vaughan EE, de Vos WM, Akkermans AD. Molecular monitoring of succession of bacterial communities in human neonates. Appl Environ Microbiol. 2002;68(1):219–226. doi:10.1128/AEM.68.1.219-226.2002.
- Eun CS, Mishima Y, Wohlgemuth S, Liu B, Bower M, Carroll IM, Sartor RB. Induction of bacterial antigen-specific colitis by a simplified human microbiota consortium in gnotobiotic interleukin-10-/- mice. Infect Immun. 2014;82(6):2239–2246. doi:10.1128/IAI.01513-13.
- Sonnenburg JL, Chen CT, Gordon JI. Genomic and metabolic studies of the impact of probiotics on a model gut symbiont and host. PLoS Biol. 2006;4(12):e413. doi:10.1371/journal.pbio.0040413.
- Johansson MEV, Gustafsson JK, Holmén-Larsson J, Jabbar KS, Xia L, Xu H, Ghishan FK, Carvalho FA, Gewirtz AT, Sjövall H, et al. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut. 2014;63(2):281–291. doi:10.1136/gutjnl-2012-303207.
- Zhao S, Liu W, Wang J, Shi J, Sun Y, Wang W, Ning G, Liu R, Hong J. Akkermansia muciniphila improves metabolic profiles by reducing inflammation in chow diet-fed mice. J Mol Endocrinol. 2017;58(1):1–14. doi:10.1530/JME-16-0054.
- Cani PD, de Vos WM. Next-generation beneficial microbes: the case of Akkermansia muciniphila. Front Microbiol. 2017;8:1765. doi:10.3389/fmicb.2017.01765.
- Thiel R, Blaut M. An improved method for the automated enumeration of fluorescently labelled bacteria in human faeces. J Microbiol Methods. 2005;61(3):369–379. doi:10.1016/j.mimet.2004.12.014.
- Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, Stahl DA. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl Environ Microbiol. 1990;56(6):1919–1925.
- Lee SH, Malone C, Kemp PF. Use of multiple 16S ribosomal RNA-targeted fluorescent probes to increase signal strength and measure cellular RNA from natural planktonic bacteria. Mar Ecol Prog Ser. 1993;101:193–201. doi:10.3354/meps101193.
- Giovannoni SJ, Delong EF, Olsen GJ, Pace NR. Phylogenetic group-specific oligodeoxynucleotide probes for identification of single microbial cells. J Bacteriol. 1988;170(2):720–726. doi:10.1128/jb.170.2.720-726.1988.
- Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):e45. doi:10.1093/nar/29.9.e45.