
ABSTRACT
The mammalian target of rapamycin (mTOR) and the phosphatidylinositol-3-kinase (PI3K)/protein kinase B or Akt (PKB/Akt) signaling pathways are considered as two but somewhat interconnected significant immune pathways which play complex roles in a variety of physiological processes as well as pathological conditions. Aberrant activation of PI3K/Akt/mTOR signaling pathways has been reported to be associated in a wide variety of human diseases. Over the past few years, growing evidence in in vitro and in vivo models suggest that this sophisticated and subtle cascade mediates the orchestration of the immune response in health and disease through exposure to probiotics. An expanding body of literature has highlighted the contribution of probiotics and PI3K/Akt/mTOR signaling pathways in gastrointestinal disorders, metabolic syndrome, skin diseases, allergy, salmonella infection, and aging. However, longitudinal human studies are possibly required to verify more conclusively whether the investigational tools used to understand the regulation of these pathways might provide effective approaches in the prevention and treatment of various disorders. In this Review, we summarize the experimental evidence from recent peer-reviewed studies and provide a brief overview of the causal relationship between the effects of probiotics and their metabolites on the components of PI3K/Akt/mTOR signaling pathways and human disease.
Introduction
Probiotics are live microorganisms which when administered in adequate amounts confer a health benefit on the host.Citation1 Decades of research have clearly demonstrated the beneficial effects of probiotics. These effects include improvement of gastrointestinal health and gut immunity, prevention of potential colonization by pathogenic bacteria and reducing the risk of certain type of cancers.Citation2–5 Most probiotic bacteria belong to the genus Lactobacillus, Bifidobacterium, Lactococcus, and Enterococcus spp., and these microorganisms offer valuable alternative approaches for future immunomodulatory and cancer prevention therapies.Citation6–9
Protein kinase B, also known as Akt (PKB/Akt), a serine/threonine protein kinase and a direct downstream effector of phosphoinositide 3-kinase (PI3K), is a key component of the PI3K/mechanistic target of rapamycin (mTOR)/Akt signaling network.Citation10,Citation11 mTOR, the target molecule of rapamycin, is a serine/threonine kinase located downstream of the PI3K/Akt pathway.Citation12 The mTOR signaling cascade involves two multiprotein complexes with different functions, mTORC1 and mTORC2.Citation13 This is highlighted in extensive studies showing that the mTOR signaling pathway has apparent regulatory impact on immune function and T-cell differentiation by integrating various micro-environmental signals.Citation14,Citation15 Detailed investigations of the mTORC signaling pathway showed that activation of mTORC1 follows activation of PI3K, which in turn, by different interactions with pyruvate dehydrogenase kinase 1 (PDK1), can phosphorylate and partially activate Akt at Threonine (Thr)-308.Citation16 Moreover, subsequent phosphorylation of Akt at Serine (Ser)-473 by mTORC2 leads to its full activation and impact on regulation of stress resistance, glucose metabolism, apoptosis, and cell proliferation through blocking of transcription factors forkhead box O1 (FOXO1)/3a.Citation17,Citation18 It has been demonstrated that up-regulation of Akt is followed by the activation of the lipid kinase PI3K via RAS guanosine triphosphate-binding proteins (GTPases).Citation19 Additionally, several evidences indicate that Akt can indirectly promote the activation of ribosomal S6 kinase (S6K) and eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1 (4E-BP1) through direct phosphorylation of mTOR.Citation11,Citation20 Studies in different models clarify the role of mTOR signaling pathway in the regulation of cell cycle via the PI3K/Akt/mTOR/S6K cascade.Citation21,Citation22 Since the PI3K/Akt/mTOR signaling pathway is extensively reviewed elsewhere, additional comprehensive description may be found in other reviews.Citation23–25 However, studies in the past few years have made progressively clear that the PI3K/Akt/mTOR cascade can respond to different stimuli to regulate signaling pathways and essential processes of cellular biology, including crucial processes such as growth, survival, proliferation, and cell metabolism that are dysregulated in different disorders.Citation23,Citation26 Aberrant activation of the PI3K/Akt/mTOR network contributes to pathological conditions, including type 2 diabetes (T2D), nonalcoholic fatty liver disease (NAFLD), and cancer. Therefore, modulating the components of the PI3K/Akt/mTOR pathway has recently been proposed as a vital and potential therapeutic option for preventing and/or treating a quite diverse host of conditions whose chronic complications are important burdens in modern communities.Citation17,Citation27,Citation28
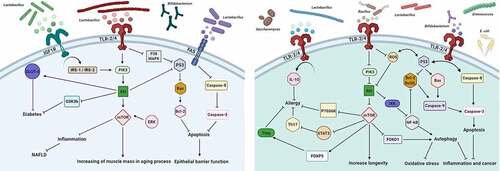
Within the last decade, the fields of immunology and microbiology have turned out to be more entwined than previously thought. Up until now, considerable effort and an ever-growing number of in vitro and in vivo studies have been conducted to investigate the possible intriguing link between the effects of probiotics on the PI3K/Akt/mTOR signaling network and their effects on different aspects of health and disease. Major efforts in recent years have been made to shed light on the mechanisms accounting for the interconnections between probiotics metabolism and PI3K/Akt/mTOR signaling pathway (). For example, several findings document a possible relationship between the consumption of probiotic strains and their derived metabolites, and the downregulation of mTOR signaling, resulting in improvement of allergic responses.Citation29 In line with this hypothesis, an important in vivo study conducted by Jeong et al., looking at the effectiveness of Lactobacillus plantarum KY1032 in controlling lipidemia in rats, discovered that the oral consumption of this probiotic can result in concomitant reduction of Akt and mTOR phosphorylation.Citation33 Similarly, the suppressing effect of Lactobacillus rhamnosus GG (LGG) strain on the phosphorylation levels of Akt/mTOR and extracellular-signal regulated kinases1/2 (ERK1/2) has been demonstrated in parallel with symptom improvement in animal models of obstructive sleep apnea (OSA).Citation45 Keeping these results in mind, probiotic strains should be selected based on their ability to regulate PI3K/Akt/mTOR signaling intermediates for targeted therapy of diseases in which activation of this pathway plays a role. In this Review, we wish to provide the Reader an exhaustive overview of how such studies have contributed to improve our understanding of the causal relationships between probiotics and the PI3K/Akt/mTOR signaling pathway (). We herein summarize our current understanding of the modulatory effects of probiotics on the PI3K/Akt/mTOR signaling pathway in specific disease models to help pave the way for novel therapeutic strategies.
Table 1. List of probiotics strain and their effect on different disorders
Figure 1. Schematic representation of the complex crosstalk between probiotics and the PI3K/AKT/mTOR signaling network, whereby extracellular and intracellular signals converge to orchestrate canonical upstream and downstream pathways to modulate a wide range of biological processes involved in various disorders. A detailed description of these interactions is provided in the text. Right: Up-regulation of PI3K/AKT/mTOR signaling cascade by probiotics. Left: Down-regulation of hyperactivated PI3K/AKT/mTOR signaling pathway by probiotics. Arrows indicate positive regulation (activation/stimulation), bar-headed lines indicate negative regulation (inhibition). The abbreviations shown in the figure can be found in the main text. (Figure was designed by https://biorender.com)
Probiotics, autophagy, and apoptosis
Autophagy, or programmed cell death (PCD) type 2, an evolutionarily conserved pathway, is a lysosome-mediated catabolic pathway and plays a critical role in degradation of the organelles and superfluous proteins that happens ubiquitously in all eukaryotic cells.Citation68 It has recently been highlighted as an innate defense or vital homeostasis mechanism against bacterial pathogens and a variety of stimuli and metabolic stress conditions, including nutrient deprivation, which is fundamental for cytoplasmic recycling, cellular bioenergy homeostasis, cellular lipid metabolism, cell survival, and lifespan extension.Citation69 Studies in animal models and in humans are starting to unravel the opposing link between autophagy and inflammation. Of note, the inflammatory response mediated by the nuclear factor kappa B (NF-κB) signaling cascade would result in inhibition of autophagy, while, conversely, an inflammatory response would be attenuated after activation of autophagy.Citation70 Within the past years, it has become increasingly clear that several key molecular and signaling pathways play a crucial role in regulating the balance of autophagy vs. non-apoptotic cell death. Among the most studies are the PI3K/Akt/mTOR signaling pathway, adenosine 50-monophosphate (AMP)-activated protein kinase (AMPK), and mitogen-activated protein kinase (MAPK)/ERK signaling pathways, which have the capability to regulate autophagy at diverse steps of autophagosome formation.Citation71,Citation72 In cancerous cells, in response to cellular stress induced by chemotherapeutics, mTOR has the potential to regulate the balance between autophagy and cell proliferation.Citation73 Nevertheless, in the presence of growth factors or nutrients, autophagy would be down-regulated following the activation of TORC1 and TORC2 by PI3K-I (class I PI3Ks) and Akt, and the phosphorylation of their downstream molecules.Citation74 In contrast, in the absence of nutrients and growth factors and/or the presence of other stressors, autophagy will be up-regulated due to the inhibition of Akt/mTOR activation.Citation75 The observation that there is negative cross-regulation between autophagy and the PI3K/Akt/mTOR signaling pathway can have interesting implications in the regulation of cellular lipid metabolism.Citation76 Recent experimental evidence gathered from animal models characterized the effects of probiotics on PI3K/Akt/mTOR signaling, autophagy, and indicators of inflammation. All in all, such studies highlighted a novel mechanism and a theoretical foundation for the inhibitory effects of probiotics on expression of the pro-inflammatory cytokines, IL-1β, IL-6, and TNF-α, and their association with reduced mTOR/FOXO1/NF-κB activity and the promotion of autophagy processes in normal cellular lipid biosynthesis.Citation77 These observations have been documented by both in vitro and in vivo studies which have probed that some strains of probiotics (e.g. Bacillus amyloliquefaciens SC06 and Bacillus licheniformis SC08) could improve oxidative stress by promoting the intestinal autophagy machinery system following inhibition of the PI3K/Akt pathway.Citation37
Aging is a multi-factorial deleterious process that accounts for increased morbidity and mortality in elderly and is accentuated in certain disease states, especially in people living with HIV (PLWH). Increasing evidence indicates that the gathering of damaged cellular components related to the aging process, due to the accumulation of reactive oxygen species (ROS), contributes to dysregulated autophagy. Remarkably, increased activity of the PI3K/Akt axis, together with inhibition of autophagy, are a causative node in several diseases attributed to the aging process, including type 2 diabetes mellitus (T2DM), neurodegeneration, cancer, and heart disease. During aging, skeletal muscle mass could be reduced due to alterations in the activity of autophagy.Citation78,Citation79 In this context, studies have documented the unique role of probiotics-induced mTORC signaling in cytoskeletal organization. In fact, increased skeletal muscle mass has been demonstrated in rats administered heat-killed Bifidobacterium breve B-3, possibly mediated by increased phosphorylation of Akt/mTOR signaling intermediates along with AMPK.Citation57 It is well known that cellular senescence plays an essential role in the aging process. In recent years, it has been demonstrated that the PI3K/Akt/mTOR cascade critically regulates various processes associated with cell senescence during aging, and can be successfully engaged to extend the longevity in aged mice and humans by improving immune function.Citation80 Striking results from animal models support the pro-longevity effects of probiotics. For example, administration of the secretory metabolites of Lactobacillus fermentum in mice models of senescence can effectively attenuate the development and progression of senescence by decreasing senescence markers such as p21WAF1, p53, p38MAPK, ROS, cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS), and senescence-associated-β-galactosidase (SA-β-gal), by inhibiting NF-κB activation and the DNA damage response, and by down-regulating the phosphorylation of PI3K/Akt/mTOR signaling intermediates.Citation39 Similar results were seen in aged Fischer 344 rats administered Lactobacillus plantarum KY1032, Lactobacillus curvatus HY7601, and Lactobacillus pentosus var. plantarum C29, in which these probiotics caused inhibition of the phosphorylation and activation of Akt/mTOR/NF-κB pathway intermediates and expression of p53, p16, and COX-2, and returned alternation of age reduced spontaneous in aged rats.Citation33,Citation67 Additional approaches will be essential to elucidate the interactions of probiotics with the PI3K/Akt/mTOR signaling pathway in the regulation of aging processes.
Apoptosis is another important form of PCD. A numbers of excellent studies have shown that the intrinsic and extrinsic apoptosis pathways play vital roles in tumor regression and are in turn regulated by the PI3K/Akt, p38 MAPK, c-Jun N-terminal kinase (JNK), and AMPK pathways.Citation81,Citation82 It has been shown that Lactobacillus rhamnosus GG and its metabolites could prevent cytokine-induced apoptosis in human or mouse intestinal epithelial cells by down-regulating the activation of p38 MAPK and up-regulating the PI3K/Akt cascade.Citation42 Both live and dead cells of Bifidobacterium animalis subsp. lactis BI-04 strain can retard Benzo(a)pyrene (BaP)-induced apoptosis of the colonic epithelial cells by up-regulating the PI3K/Akt signaling pathway and down-regulating p53 gene expression.Citation56 In light of these findings, some researchers have hypothesized that the induction of apoptosis in the SW480 cell line could occur in the presence of heat-killed preparations of Saccharomyces cerevisiae through enhanced expression of BAX, cleaved caspase-3, and cleaved caspase-9 and reduced expression of p-Akt1, Bcl-XL, pro-caspase 3 and 9, which are involved in the Akt/NF-κB signaling pathway.Citation36 Therefore, by targeting apoptosis and survival-related signaling pathways these probiotics may offer important therapeutic options for cancer management.
Ample evidence indicates a critical role for vascular events resulting in impaired blood flow and blood vessel damage in the brain in the development of vascular dementia (VaD). VaD is implicated in behavioral deterioration, progressive cognitive, memory and learning deficits, associated with neurodegeneration and neural lesions. Conclusive findings linking neuronal apoptosis to a low ratio of Bcl-2/BAX have been obtained in studies of hippocampal lesions in rat models of VaD. Conversely, it has been shown that the up-regulation of PI3K/Akt pathway can rescue neurons from apoptosis activated in cerebral ischemia-reperfusion (IR) injury.Citation83,Citation84 Recent studies highlight the potential role of Akt/mTOR signaling in the prevention of vascular cognitive damage by probiotics. These observations indicate that probiotics such as Clostridium butyricum may prevent VaD by increasing the levels of Bcl-2, BDN, and Akt phosphorylation and decreasing the levels of BAX, leading to reduced neuronal apoptosis and improvement of cognitive function in bilateral common carotid artery occlusion (BCCAO)-induced VaD. Taken together, the aforementioned data strongly point to the modulation of apoptosis, via the regulation of Bcl-2/BAX ratio, as a possible therapeutic target in VaD.Citation52
Probiotics and their surface components
Emerging studies have demonstrated that the anti-inflammatory immune mechanisms and anti-tumoral effects of probiotic are closely associated with their surface components. This has prompted investigators to look at the key components of the cell surface of probiotics such as lipoteichoic acid (LTA), surface-layer protein (Slp), and exopolysaccharides (EPS).Citation85 LTA, an amphiphilic negatively charged glycolipid, is an immune-stimulatory component of the cell wall of probiotics that promotes the attachment of probiotics to host cells, colonization, and invasion.Citation46 At present, there are conflicting findings in the sparse literature on the role of LTA in the regulation of inflammation. In some of these studies, LTA appears to play an important role in severe inflammatory responses and the pathogenesis of septic shock by stimulating the production of cytokines, such as IL-6 and TNF-α, via toll-like receptors (TLRs) such as TLR2.Citation46,Citation86 The stimulatory role of LTA in the development of septic shock is supported by in vivo evidence in a mouse model in which oral administration of Lactobacillus acidophilus strain NCFM deficient in LTA (NCK2025), compared to administration of the wild-type parental strain (NCK56), can down-regulate the phosphorylation of Akt1, p38 and cytosolic group IV PLA2 in colonic epithelial cells and dendritic cells, but significantly augment phosphorylation of ERK1/2, which highlights the overall immunosuppressive effect of NCK2025 in T-cell-induced colitis.Citation46 However, in apparent conflict with these findings, data strongly suggest that LTA can play a major role in the generation of anti-inflammatory cytokines, such as IL-10, and in maintaining intestinal cell homeostasis through up-regulation of ERK1/2 signaling.Citation32,Citation87 These opposing effects are important caveats when trying to envisage and/or interpret the aftereffects of probiotics treatment. As of today, the functional properties of LTA are under intensive scrutiny in in vivo studies to fully elucidate its overall effects on innate immunity. It has long been known that inflammation can be promoted by macrophages through the production of different cytokines, concomitant with the uptake and eradication of pathogens.Citation46,Citation88 Regarding this issue, it is becoming increasingly clear that probiotics can sustain the immune response by promoting autophagy in pathogen-loaded macrophages. It is generally accepted that Enterococcus faecalis LTA can stimulate macrophages autophagy in vitro via hindering the phosphorylation of mTOR and Akt.Citation38 The molecular mechanism underlying the anti-tumoral properties of Slp derived from Lactobacillus acidophilus NCFM were reported in HCT116 cells. These data support the notion that Slp can induce autophagic cell death following inhibition of cell proliferation by controlling the JNK and mTOR signaling pathways via reducing the phosphorylation levels of 4E-BP1, p70, and S6, and by up-regulating the expression of p53 and p21, and generation of ROS.Citation47 It has been established that, in addition to Lactobacillus, Kluyveromyces marxianus and Pichia kudriavzevii fulfill the major criteria for probiotics definition. Of note, also the EPSs of these strains can induce apoptosis and may treat CRC by interfering with the mTOR, Janus Kinase 1 (JAK-1), and Akt-1 pathways.Citation48
Probiotics and Gastrointestinal (GI) disorders
Disruption in barrier function and alterations in tight junctions (TJ) structure are linked to the development of chronic inflammation. These changes may lead to various complications, including immune cell infiltration, expression of inflammatory cytokines, translocation of bacteria, and finally stimulation of systemic inflammatory responses.Citation89 Several findings also suggest that mechanisms controlling cell apoptosis and proliferation, that is, epithelial turnover, are deeply implicated in preserving intestinal integrity, whereby an augmented rate of apoptosis in epithelial cells is associated with injury of the intestinal mucosa.Citation43,Citation90,Citation91 This is highlighted in extensive studies showing that an impairment of epithelial barrier function and reduced synthesis of TJ-associated proteins were subsequent to p-S6K and p-mTOR down-regulation in response to reduced Akt and p-Akt protein expression in dextran sulfate sodium (DSS)-treated mice.Citation20,Citation40 Studies conducted over the last few years have contributed to our understanding of the causal association between the activation of the PI3K/Akt/mTOR signaling pathway, the induction of TJ-associated proteins and promotion of intestinal epithelial barrier function. Given the potential connection between gut microbiota and GI inflammatory disorders, several authors have been prompted to investigate the properties of beneficial microbes in experimental models of inflammation to identify new possible approaches to manage these conditions. It was long believed that Lactobacillus strains have the ability to prevent many human diseases and cancer development, and enhance the production of anti-inflammatory cytokines involved in innate immunity through the modulation of the PI3K/Akt/mTOR pathway, given its involvement in various cellular processes, such as apoptosis, inflammatory responses, and tumor angiogenesis.Citation63 Subsequent studies revealed that epithelial barrier function both in in vitro and in vivo could be markedly improved by pretreatment with such probiotics as Bifidobacterium bifidum, Lactobacillus reuteri, and LGG and its proteins (p75 and p40).Citation43,Citation51,Citation92 Similar results were seen in mice models, where ingestion of LGG and Lactobacillus reuteri ZJ617, by reducing autophagy and apoptosis via the activation of the mTOR cascades, and therefore improving TJ integrity, hinder lipopolysaccharides (LPS)-stimulated intestinal barrier dysfunction.Citation43 This view is also supported to a certain extent in studies showing that Clostridium butyricum can contribute to improving the intestinal barrier function in a mouse model of DSS-induced colitis. The above findings provided a basis to further explore the underlying mechanism of these beneficial properties of Clostridium butyricum, which was found to increase the expression of TJ-associated proteins and diminish intestinal mucosal permeability via the up-regulation of the Akt/mTOR axis and the phosphorylation of their downstream signaling molecules such as p70S6k.Citation20 Accumulating in vitro and in vivo evidence indicates that such probiotic strains as Lactobacillus fermentum L930BB, Lactobacillus paragasseri K7, Bifidobacterium animalis subsp. animalis (IM386), and Lactobacillus plantarum WCFS1, via the up-regulation of protein kinase C (PKC) and PI3K/Akt anti-apoptotic pathways, can regulate the actin cytoskeleton and TJ structure to ensure reconstitution of the intestinal epithelial barrier.Citation34,Citation40 Most importantly, the PI3K/Akt/mTOR network complex, via cooperation with TLR-delivered signals and NF-κB, plays fundamental roles in the development of inflammatory responses. In this context, it is documented that Lactobacillus gasseri JM1 could improve inflammation in Caco-2 cells treated with LPS by up-regulation of TLR2 and nucleotide-binding oligomerization domain containing 2 (NOD2)-mediated PI3K/Akt network.Citation54
Dysregulation and hyperactivation of the PI3K/Akt/mTOR signaling network are closely associated with cell proliferation, resistance to apoptosis, and metastasis formation, and ultimately contribute to the progression of inflammatory bowel disease (IBD), colorectal cancer (CRC), and gastric cancer.Citation93,Citation94 Additional studies have addressed how the PI3K/Akt/mTOR signaling pathway can mediate control of colon and gastric cancer development by probiotics. In keeping with this, studies in murine models have demonstrated that Lactobacillus casei Zhang (LCZ) and VSL#3 could prevent and treat ulcerative colitis by decreasing the production of host inflammatory cytokines, inhibiting the hyperactivation of the PI3K/Akt/NF-κB pathways and the phosphorylation of signal transducer and activator of transcription 3 (STAT3).Citation63,Citation95 Well-designed studies in murine models have also shown that administration of Leuconostoc mesenteroides and Lactobacillus salivarius Ren could significantly induce cell apoptosis in colon cancer cell lines and block colon cancer progression. The proposed mechanisms underlying the anti-tumoral effects of these strains included inactivation of the NF-κB and Akt signaling cascades and their downstream molecules (cyclin D1 and COX-2), down-regulation of anti-apoptotic protein like Bcl-XL and Bcl-2, and up-regulation of MAPK1, caspase 3 and 8, and BAX.Citation64,Citation65 In light of these findings, it is worth mentioning that Lactobacillus paracasei such as species X12 can block the G1 phase of human colorectal adenocarcinoma cell cycle by down-regulating the mTOR/4E-BP1 signaling pathway through the up-regulation of p27 and the down-regulation of cyclin E1.Citation61 Similarly reduced phosphorylation of PI3K/Akt/NF-κB signaling intermediates by probiotics was later confirmed in gastric cancer cells by Hwang and colleagues. They showed that Lactobacillus casei extracts can up-regulate apoptosis in gastric cancer cells and prevent gastric cancer by down-regulating the phosphorylation of PI3K/Akt/NF-κB signaling components.Citation60 As shown in new report, Lactobacillus plantarum can inhibit both Helicobacter pylori colonization and a gastric cancer cell line (AGS) through the up-regulation of PTEN, BAX, TLR4, and the down-regulation of Akt genes.Citation30 Hence, blocking the hyperactivation of the PI3K/Akt/mTOR signaling pathway has emerged as a plausible therapeutic target for CRC and gastric cancers because of its involvement in cell growth and proliferation. However, the mechanisms of action involved in the beneficial effects of probiotics mediated through the PI3K/Akt/mTOR signaling pathway in GI disorders are not fully elucidated in relevant human models. Thus, determining the exact mechanisms of action is not only crucial to determining the pathogenesis of GI disorders, but would also provide a new basis for emerging therapies. To this end, prospective clinical trials should be launched to better characterize patients diagnosed with GI disorders.
Probiotics and Metabolic syndrome
The metabolic syndrome (MetS) is defined by the coexistence of risk factors such as increased abdominal fat, obesity, high blood sugar, high blood pressure, and hyperlipidemia that predispose to the development of T2D, cardiovascular disease (CVD), and NAFLD.Citation96 As indicated earlier, over-activation of the mTOR pathway is associated with the process of autophagy, leading to several metabolic disorders.Citation17 Given the possible connection between probiotics and MetS, many authors have also been encouraged to investigate the effects of probiotics on PI3K/Akt signaling and MetS development in murine models. The following paragraphs provide an overview of the probiotics investigated in studies of the PI3K/Akt/mTOR signaling pathway and of their effects on MetS progression.
Type 2 diabetes (T2D)
T2D is characterized by disordered glucose metabolism as a result of insulin resistance. Attachment of insulin to insulin receptor substrate 2 (IRS-2) results in phosphorylation of PI3K and Akt, which improves glucose metabolism by phosphorylating glycogen synthase kinase 3 beta (GSK-3β). Conversely, blocking the activity of the PI3K/Akt signaling pathway following the inhibition of IRS-2 phosphorylation may cause high glucose concentrations because of the hyperphosphorylation of GSK-3β.Citation97 A mounting body of evidence indicates that this dysfunction could be reversed in the presence of probiotic microorganisms. For example, oral administration of Lactobacillus paracasei TD062 to diabetic mice could ameliorate the insulin response and glucose homeostasis via decreasing the levels of GSK-3β and elevating those of IRS-2, PI3K and Akt, thus preventing the development of T2DM.Citation62
Two significant factors contributing to lipogenesis and insulin resistance are life-span determinant p66Shc (a 66 kDa proto-oncogene Src homologous-collagen homolog (Shc) adaptor protein) and the mTOR/S6K cascades. Data so far collected indicate that dysregulation in insulin signaling pathway, insulin resistance in muscle and liver tissues and lipid accumulation occurs following aberrant up-regulation of mTOR signaling. In particular, over-activation of mTORC1 and mTORC2 results in insulin resistance and gluconeogenesis suppression by up-regulation of S6K1 and Akt signaling, respectively. These events are followed by deposition of extra fat in the liver, hindering insulin signaling, and activation of glycolysis.Citation98,Citation99 In vivo studies observed that Bifidobacterium spp. such as Bifidobacterium lactis Bb-12 can reduce blood glucose levels by up-regulating the expressions of proteins involving in the insulin signaling pathway like insulin receptor substrate 1 and insulin receptor beta, down-regulating the excess activation of the mTOR and p66Shc pathways, and increasing the expression of certain adipokines, nuclear factor-kappa B inhibitor alpha (IκBα), and IκB kinase alpha (IKKα).Citation55,Citation100
It has been known for long time that the pancreas is the only tissue that secretes insulin and has a significant role in regulating glucose metabolism. Recent in vivo findings reveal that protection of the pancreas from β-cell apoptosis is mediated by probiotics via induction of the PI3K/Akt signaling pathway. These data are consistent with the results of an in vivo study by Wang et al., in which probiotics could protect β-cells against apoptosis by up-regulating the expression of anti-apoptotic proteins and the PI3K/Akt signaling cascade, and down-regulating the expression of inflammatory factors and pro-apoptotic proteins.Citation101 In line with this concept, a recent in vitro study by Huang et al. looked at the relation between surface components of probiotics and the Akt/mTOR pathway and its inverse association during diabetes onset. Their findings suggest that EPS of Lactobacillus plantarum H31 exerts anti-diabetic effects and plays an overall important role in glucose metabolism by up-regulating the expression of Akt-2, AMPK, and human glucose transporter 4 (GLUT-4), by interfering with the pancreas α-amylase activity.Citation35
Cardio vascular disease (CVD)
Several pathways have been implicated in orchestrating the cellular response in CVD. Findings reveal that molecules involved in insulin-like growth hormone (IGF-I)-related survival pathways, including IGF-1 receptor (IGF-IR), IGF-I, p-Akt and p-PI3K, regulate cardiac survival pathways.Citation102 A substantial amount of evidence indicates that an important mechanism in the development of obesity-related heart disease in high-fat diet-fed rats is the dysregulation of anti-apoptotic along with cardiac IGF-I/PI3K/Akt-dependent survival cascades.Citation103 Collectively, previous studies have provided strong evidence showing that probiotic supplementation can enhance the activity of survival pathways in obese hearts, suggesting an interplay between IGF1/PI3K/Akt cell survival pathways and some strains of probiotics in the regulation of cardiovascular homeostasis. Along the same line, the in vivo effects of probiotics on CVD were investigated by Lin et al. and Wang et al., who found that oral administration of multi-strain probiotic groups can decrease cardiac apoptosis by up-regulating the IGF-I/PI3K/Akt survival pathway.Citation103,Citation104 In parallel studies, it was also reported that high expression of Fas ligand and its receptor Fas are linked to the progression of cardiomyocyte apoptosis. Accordingly, in vivo studies documented that heat-killed Lactobacillus reuteri GMNL-263 has similar effects on the reduction of cardiomyocyte apoptosis through down-regulating Fas ligand and up-regulating the IGF1R/PI3K/Akt cell survival pathway, thus recovering the myocardial disarray.Citation50
Nonalcoholic fatty liver disease (NAFLD)
One of the hepatic manifestations of MetS is NAFLD, a spectrum of diseases such as nonalcoholic steatohepatitis (NASH), cirrhosis, hepatocellular carcinoma (HCC), and steatosis, characterized by inflammation, increased risk for liver carcinogenesis and fibrosis, and hepatocyte damage/death.Citation105,Citation106 Emerging lines of evidence have shown that modulation of autophagy and NF-κB-mediated inflammatory responses are important mechanisms in the pathogenesis of NAFLD.Citation107 It has been found that both Lactobacillus rhamnosus and its metabolites, along with bone marrow mesenchymal stem cells (BMMSCs), by decreasing autophagy and inflammation through down-regulation of the PI3K/mTOR/NF-κB pathways, have the potential to mitigate alcoholic hepatitis and alleviate its symptoms in in vivo.Citation41
Some findings imply that high-fructose diets can enhance the prevalence of NAFLD, lipid accumulation, expression of fatty acid synthase (FASN), hepatic weight, but inhibit the expression and phosphorylation of IRS-1/Akt/endothelial nitric oxide synthase (eNOS) signaling intermediates in the liver. On the other hand, these events could be reversed following Lactobacillus plantarum supplementation in high-fructose-fed rats, revealing a possible beneficial effect of probiotics on renal and hepatic dysfunction and the prevention of NAFLD.Citation31 In this respect, Clostridium butyricum MIYAIRI 588 has been used as a butyrate-producing probiotic in NAFLD rats subject to choline-deficient/L-amino acid-defined (CDAA)-diet. In this study Clostridium butyricum MIYAIRI 588 could indeed prevent NAFLD progression by reducing the deposition of hepatic lipid and significantly decreasing the content of triglycerides, reversing insulin resistance, and hepatic inflammatory indexes. This preventing effect is a result of substantially elevated levels of expression of proteins contributing to lipolysis or lipogenesis, along with Akt and hepatic AMPK.Citation53 In line with this notion, an in vivo study documented a reversing effect of Lactobacillus johnsonii N6.2 supplementation on the hyperactivation of mTORC1-activating phosphorylation of pAkt in high-fat diet-fed regime and the reduction of metabolic syndrome-associated changes.Citation59
Probiotics and other disorders
With increasing challenges to human health, probiotics have attracted much attention for their capability to modulate the TLR/PI3K/Akt signaling pathway and ameliorate pharyngitis.Citation66 TLRs as pattern-recognition receptors (PRRs) can activate innate immunity via sensing invasion of microbial pathogens. Among TLRs, TLR2 orchestrates innate immune and inflammatory responses by recruiting macrophages and modulating PI3K/Akt pathway-dependent autophagy, respectively, when encountering pathogens such as Staphylococcus aureus.Citation108 Activation of TLR/PI3K/Akt pathway contributes to the activation of the downstream NF-κB signaling cascade, thereby stimulating the expression of pro-inflammatory mediators and the development of inflammatory diseases such as pharyngitis.Citation109 Recent in vitro and in vivo studies recognized that probiotic Lactobacillus salivarius AR809 could attenuate the inflammatory response produced by S. aureus by elevating autophagic protein level and blocking the TLR/PI3K/Akt/mTOR/NF-κB signaling network.Citation66 A substantial amount of evidence demonstrates that administration of probiotics such as Bifidobacterium animalis subsp. lactis DSM10140 and/or LGG in Caco-2 cells can differentially affect the IL-8 response to Salmonella based on the time of administration, in that administration of these probiotics before Salmonella infection can enhance the activation of the PI3K/Akt signaling pathway, while it inhibits PI3K/Akt pathway activation after Salmonella infection.Citation44 As in many other cases, this information is not highly predictive of the overall effect of probiotics and provides an incomplete picture of how they regulate the PI3K/Akt pathway in Salmonella infection. Thus, further experimental studies are required to elucidate this point.
Mucosal delivery of probiotics has been the subject of growing interest due to its proven therapeutic effects in inflammatory and allergic disorders.Citation110 Development of food hypersensitivity reactions occurs due to production of food protein-specific immunoglobulin E (IgE) and stimulation of basophils or mast cells, as a result of the activation of imbalanced, T helper 2 (Th2)-biased immune responses.Citation111 Extensive studies have been conducted to support the efficacy of probiotic microorganisms in decreasing food allergy symptoms via the regulation of mTOR and Treg cells. An in vivo study showed that consumption of Bacillus coagulans 09.712 up-regulates the transcription factor forkhead box P3 (Foxp3) and down-regulates the phosphorylation of Akt, 4E-BP1, STAT3, serum/glucocorticoid regulated kinase 1 (SGK1) and p70S6K by inhibiting mTOR. Hence, the induction of the anti-inflammatory cytokine IL-10 and of CD4+Foxp3+ regulatory T cells (Tregs), and, conversely, the inhibition of T helper 17 (Th17) and Th2-predominant can cause the alleviation of food allergic inflammation.Citation29 This notion is further confirmed in studies showing that Bifidobacterium breve (BbC50sn), by regulating p38MAPK, ERK, and PI3K/Akt signaling pathways, can exert beneficial effects on allergic and inflammatory disorders depending on its interaction with monocyte-derived dendritic cells (DCs), leading to DC-induced IL-10 production and DC maturation, activation, and survival.Citation58 Whereas most similar studies are concordant on the beneficial effects of probiotics on alleviation of allergic disorders, further research efforts are required to answer critical questions on the mechanisms underlying definite aspects of human-probiotic relations to fully understand the therapeutic potential of probiotics in allergic and inflammatory disorders.
The process of melanin synthesis called melanogenesis contributes significantly to protection of the skin from UV radiation. Extreme accumulation of melanin causes hyperpigmentation disorders.Citation112 Topical depigmentation agents have long been employed in the treatment and/or prevention of these disorders. Notably, it is well documented that the PI3K/Akt signaling cascade in cutaneous stem cells is critically involved in hyperpigmentation disorders and can control their apoptosis and proliferation both in human and murine models.Citation113 As a result, this pathway has been considered a suitable target in several types of skin disorders. Studies in B16F10 mouse melanoma cells have provided evidence to suggest that LTA isolated from Lactobacillus plantarum can prevent melanogenesis by inhibiting the expression of melanogenic enzymes and cellular activity of tyrosinase, while activating the PI3K/Akt pathway and ERK.Citation32 A very recent study has demonstrated that exposure of a cell-free extract of Enterococcus faecium L-15 to mouse skin-derived precursor cells (SKPs) could result in improvement of self-renewal and proliferation of SKPs by activation of the PI3K/Akt signaling pathway. Therapies with mesenchymal stem cells (MSCs) have been confirmed to accelerate wound healing of skin and mucosa due to increased tissue regeneration rates.Citation6 However, a microecological imbalance can impair the function of MSCs, leading to retarded wound healing. Recent in vivo investigations indicate that Lactobacillus reuteri extracts can accelerate the wound healing process in the oral mucosa by stimulating the functions of gingiva MSCs (GMSCs). Mechanisms accounting for these effects include the enhancement of active-β-catenin and transforming growth factor β1 (TGFβ1) expression and of PI3K/Akt pathway phosphorylation.Citation49 Additional work and more clinical evidence are needed to fully understand whether the effects of probiotics on hyperpigmentation disorders are mediated by an underlying regulation of the PI3K/Akt signaling cascade.
Conclusion
The PI3K/Akt/mTOR signaling pathway is one of the most important signal transduction pathways that have many biological functions and drives numerous cellular and physiological processes in the body. Extensive research has provided a better knowledge of the molecular mechanisms controlling the PI3K/Akt/mTOR signaling cascades. As reviewed herein, a vast body of evidence demonstrates that aberrant activation and/or dysregulation in the major components of the PI3K/Akt/mTOR signaling pathways are identifiable in different diseases, including most, if not all, human cancers. The last decade has witnessed much progress in our knowledge of the properties of lactic acid bacteria (LAB) as probiotic candidates,Citation7,Citation114 which include their modulatory functions on specific targets of the PI3K/Akt/mTOR signaling pathway, and their promising effects on infection control. Driven by earlier evidence of their effects on the PI3K/AKT/mTOR axis, research focused on elucidating the molecular mechanisms of various probiotics and their derived metabolites has rapidly gained momentum. Most current efforts are directed toward the characterization of innovative approaches to target this important pathway in the attempt to develop promising and selective treatment options. In this review, we extensively discussed a number of in vitro and in vivo studies conducted with several strains of probiotics to understand their mechanisms of action on the PI3K/Akt/mTOR network in several disease models. However, despite intensive efforts by many researchers, nearly all of the studies conducted to date have been conducted in vitro or in animal models. Thus, definitive evidence for their beneficial effects in human disease is still lacking, and more research needs to be carried out in human subjects and clinical samples. Such studies, and in particular those concentrating on probiotic strains with documented effects on components of the PI3K/Akt/mTOR pathway, would be likely to provide more conclusive outcomes and support further exploration for new therapeutic candidates for the treatment of various, highly prevalent disorders.
Authors’ contributions
All the authors equally contributed to this work. Also, all authors read and approved the final manuscript.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Acknowledgments
We deeply appreciate the continued support both scientific and intellectual of our groups who collaborated sincerely in this study deprived of any expectations and whose names have not been mentioned.
Additional information
Funding
References
- Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, Morelli L, Canani RB, Flint HJ, Salminen S, et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014;11:506–17. doi:10.1038/nrgastro.2014.66.
- Guarner F, Malagelada JR. Gut flora in health and disease. Lancet. 2003;361:512–519. doi:10.1016/S0140-6736(03)12489-0.
- Kim JY, Woo HJ, Kim Y-S, Lee HJ. Screening for antiproliferative effects of cellular components from lactic acid bacteria against human cancer cell lines. Biotechnol Lett. 2002;24:1431–1436. doi:10.1023/A:1019875204323.
- Wang SM, Zhang LW, Fan RB, Han X, Yi HX, Zhang LL, Xue C-H, Li H-B, Zhang Y-H, Shigwedha N, et al. Induction of HT-29 cells apoptosis by lactobacilli isolated from fermented products. Res Microbiol. 2014;165:202–214. doi:10.1016/j.resmic.2014.02.004.
- Hirayama K, Rafter J. The role of probiotic bacteria in cancer prevention. Microbes Infect. 2000;2:681–686. doi:10.1016/S1286-4579(00)00357-9.
- Kim K, Park S, Kim H, Min S, Ku S, Seo J, Roh S. Enterococcus faecium L-15 Extract Enhances the Self-Renewal and Proliferation of Mouse Skin-Derived Precursor Cells. Probiotics Antimicrob Proteins. 2020;12:1492–1501. doi:10.1007/s12602-020-09635-w.
- Taghinezhad-S S, Keyvani H, Bermúdez-Humarán LG, Donders GGG, Fu X, Mohseni AH. Twenty years of research on HPV vaccines based on genetically modified lactic acid bacteria: an overview on the gut-vagina axis. Cell Mol Life Sci. 2020:1–16. doi:10.1007/s00018-020-03652-2.
- Kim MJ, Ku S, Kim SY, Lee HH, Jin H, Kang S, et al. Safety Evaluations of Bifidobacterium bifidum BGN4 and Bifidobacterium longum BORI. Int J Mol Sci. 2018;19:1422. doi:10.3390/ijms19051422.
- Singh S, Sharma RK, Malhotra S, Pothuraju R, Shandilya UK. Lactobacillus rhamnosus NCDC17 ameliorates type-2 diabetes by improving gut function, oxidative stress and inflammation in high-fat-diet fed and streptozotocintreated rats. Benef Microbes. 2017;8:243–255. doi:10.3920/BM2016.0090.
- Cicenas J. The potential role of Akt phosphorylation in human cancers. Int J Biol Markers. 2008;23:1–9. doi:10.1177/172460080802300101.
- Manning BD, Toker A. AKT/PKB Signaling: navigating the Network. Cell. 2017;169:381–405. doi:10.1016/j.cell.2017.04.001.
- Huang S, Houghton PJ. Targeting mTOR signaling for cancer therapy. Curr Opin Pharmacol. 2003;3:371–377. doi:10.1016/S1471-4892(03)00071-7.
- Fingar DC, Blenis J. Target of rapamycin (TOR): an integrator of nutrient and growth factor signals and coordinator of cell growth and cell cycle progression. Oncogene. 2004;23:3151–3171. doi:10.1038/sj.onc.1207542.
- Powell JD, Pollizzi KN, Heikamp EB, Horton MR. Regulation of immune responses by mTOR. Annu Rev Immunol. 2012;30:39–68. doi:10.1146/annurev-immunol-020711-075024.
- Albert MH, Mannert J, Fleischmann KK, Schiemann M, Pagel P, Schmid I, Magg T. MiRNome and transcriptome aided pathway analysis in human regulatory T cells. Genes Immun. 2014;15:303–312. doi:10.1038/gene.2014.20.
- Sarbassov DD, Guertin DA, Ali SM, Sabatini DM. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science. 2005;307:1098–1101. doi:10.1126/science.1106148.
- Saxton RA, Sabatini DM. mTOR Signaling in Growth, Metabolism, and Disease. Cell. 2017;168:960–976. doi:10.1016/j.cell.2017.02.004.
- Xiong X, Tao R, DePinho RA, Dong XC. Deletion of hepatic FoxO1/3/4 genes in mice significantly impacts on glucose metabolism through downregulation of gluconeogenesis and upregulation of glycolysis. PLoS One. 2013;8:e74340. doi:10.1371/journal.pone.0074340.
- Bossler F, Hoppe-Seyler K, Hoppe-Seyler F. PI3K/AKT/mTOR Signaling Regulates the Virus/Host Cell Crosstalk in HPV-Positive Cervical Cancer Cells. Int J Mol Sci. 2019;20:2188. doi:10.3390/ijms20092188.
- Liu M, Xie W, Wan X, Deng T. Clostridium butyricum protects intestinal barrier function via upregulation of tight junction proteins and activation of the Akt/mTOR signaling pathway in a mouse model of dextran sodium sulfate-induced colitis. Exp Ther Med. 2020;20:25. doi:10.3892/etm.2020.9138.
- Roulin D, Cerantola Y, Dormond-Meuwly A, Demartines N, Dormond O. Targeting mTORC2 inhibits colon cancer cell proliferation in vitro and tumor formation in vivo. Mol Cancer. 2010;9:1476–4598. doi:10.1186/1476-4598-9-57.
- Gulhati P, Bowen KA, Liu J, Stevens PD, Rychahou PG, Chen M, Lee EY, Weiss HL, O’Connor KL, Gao T, et al. mTORC1 and mTORC2 regulate EMT, motility, and metastasis of colorectal cancer via RhoA and Rac1 signaling pathways. Cancer Res. 2011;71:3246–3256. doi:10.1158/0008-5472.CAN-10-4058.
- Porta C, Paglino C, Mosca A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front Oncol. 2014;4. doi:10.3389/fonc.2014.00064.
- Liu GY, Sabatini DM. mTOR at the nexus of nutrition, growth, ageing and disease. Nat Rev Mol Cell Biol. 2020;21:183–203. doi:10.1038/s41580-019-0199-y.
- Ersahin T, Tuncbag N, Cetin-Atalay R. The PI3K/AKT/mTOR interactive pathway. Mol Biosyst. 2015;11:1946–1954. doi:10.1039/C5MB00101C.
- Lim HJ, Crowe P, Yang JL. Current clinical regulation of PI3K/PTEN/Akt/mTOR signalling in treatment of human cancer. J Cancer Res Clin Oncol. 2015;141:671–689. doi:10.1007/s00432-014-1803-3.
- Janku F, Yap TA, Meric-Bernstam F. Targeting the PI3K pathway in cancer: are we making headway? Nat Rev Clin Oncol. 2018;15:273–291. doi:10.1038/nrclinonc.2018.28.
- Procaccini C, De Rosa V, Galgani M, Abanni L, Calì G, Porcellini A, Carbone F, Fontana S, Horvath TL, La Cava A, et al. An oscillatory switch in mTOR kinase activity sets regulatory T cell responsiveness. Immunity. 2010;33:929–941. doi:10.1016/j.immuni.2010.11.024.
- Fu L, Peng J, Zhao S, Zhang Y, Su X, Wang Y. Lactic acid bacteria-specific induction of CD4(+)Foxp3(+)T cells ameliorates shrimp tropomyosin-induced allergic response in mice via suppression of mTOR signaling. Sci Rep. 2017;7:017–02260. doi:10.1038/s41598-017-02260-8.
- Maleki-Kakelar H, Dehghani J, Barzegari A, Barar J, Shirmohamadi M, Sadeghi J, Omidi Y. Lactobacillus plantarum induces apoptosis in gastric cancer cells via modulation of signaling pathways in Helicobacter pylori. Bioimpacts. 2020;10:65–72. doi:10.34172/bi.2020.09.
- Sumlu E, Bostancı A, Sadi G, Alçığır ME, Akar F. Lactobacillus plantarum improves lipogenesis and IRS-1/AKT/eNOS signalling pathway in the liver of high-fructose-fed rats. Arch Physiol Biochem. 2020;18:1–9. doi:10.1080/13813455.2020.1727527.
- Kim HR, Kim H, Jung BJ, You GE, Jang S, Chung DK. Lipoteichoic acid isolated from Lactobacillus plantarum inhibits melanogenesis in B16F10 mouse melanoma cells. Mol Cells. 2015;38:163–170. doi:10.14348/molcells.2015.2263.
- Jeong JJ, Kim KA, Ahn YT, Sim JH, Woo JY, Huh CS, Kim D-H. Probiotic Mixture KF Attenuates Age-Dependent Memory Deficit and Lipidemia in Fischer 344 Rats. J Microbiol Biotechnol. 2015;25:1532–1536. doi:10.4014/jmb.1505.05002.
- Paveljšek D, Ivičak-Kocjan K, Treven P, Benčina M, Jerala R, Rogelj I. Distinctive probiotic features share common TLR2-dependent signalling in intestinal epithelial cells. Cell Microbiol. 2021;23:1. doi:10.1111/cmi.13264.
- Huang Z, Lin F, Zhu X, Zhang C, Jiang M, Lu Z. An exopolysaccharide from Lactobacillus plantarum H31 in pickled cabbage inhibits pancreas α-amylase and regulating metabolic markers in HepG2 cells by AMPK/PI3K/Akt pathway. Int J Biol Macromol. 2020;143:775–784. doi:10.1016/j.ijbiomac.2019.09.137.
- Shamekhi S, Abdolalizadeh J, Ostadrahimi A, Mohammadi SA, Barzegari A, Lotfi H, Bonabi E, Zarghami N. Apoptotic Effect of Saccharomyces cerevisiae on Human Colon Cancer SW480 Cells by Regulation of Akt/NF-ĸB Signaling Pathway. Probiotics Antimicrob Proteins. 2020;12:311–319. doi:10.1007/s12602-019-09528-7.
- Wu Y, Wang B, Xu H, Tang L, Li Y, Gong L, Wang Y, Li W. Probiotic Bacillus Attenuates Oxidative Stress-Induced Intestinal Injury via p38-Mediated Autophagy. Front Microbiol. 2019;10. doi:10.3389/fmicb.2019.02185.
- Lin D, Gao Y, Zhao L, Chen Y, An S, Peng Z. Enterococcus faecalis lipoteichoic acid regulates macrophages autophagy via PI3K/Akt/mTOR pathway. Biochem Biophys Res Commun. 2018;498:1028–1036. doi:10.1016/j.bbrc.2018.03.109.
- Kumar R, Sharma A, Gupta M, Padwad Y, Sharma R. Cell-Free Culture Supernatant of Probiotic Lactobacillus fermentum Protects Against H2O2-Induced Premature Senescence by Suppressing ROS-Akt-mTOR Axis in Murine Preadipocytes. Probiotics Antimicrob Proteins. 2020;12:563–576. doi:10.1007/s12602-019-09576-z.
- Paveljšek D, Juvan P, Košir R, Rozman D, Hacin B, Ivičak-Kocjan K, Rogelj I. Lactobacillus fermentum L930BB and Bifidobacterium animalis subsp. animalis IM386 initiate signalling pathways involved in intestinal epithelial barrier protection. Benef Microbes. 2018;9:515–525. doi:10.3920/BM2017.0107.
- Cai C, Chen DZ, Ge LC, Chen WK, Ye SS, Ye WW, Tao Y, Wang R, Li J, Lin Z. Synergistic effects of Lactobacillus rhamnosus culture supernatant and bone marrow mesenchymal stem cells on the development of alcoholic steatohepatitis in mice. Am J Transl Res. 2019;11:5703–5715.
- Yan F, Cao H, Cover TL, Whitehead R, Washington MK, Polk DB. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology. 2007;132:562–575. doi:10.1053/j.gastro.2006.11.022.
- Cui Y, Liu L, Dou X, Wang C, Zhang W, Gao K, Liu J, Wang H. Lactobacillus reuteri ZJ617 maintains intestinal integrity via regulating tight junction, autophagy and apoptosis in mice challenged with lipopolysaccharide. Oncotarget. 2017;8:77489–77499. doi:10.18632/oncotarget.20536.
- Huang FC, Huang SC. The different effects of probiotics treatment on Salmonella-induced interleukin-8 response in intestinal epithelia cells via PI3K/Akt and NOD2 expression. Benef Microbes. 2016;7:739–748. doi:10.3920/BM2015.0181.
- Liu J, Li T, Wu H, Shi H, Bai J, Zhao W, Jiang D, Jiang X. Lactobacillus rhamnosus GG strain mitigated the development of obstructive sleep apnea-induced hypertension in a high salt diet via regulating TMAO level and CD4(+) T cell induced-type I inflammation. Biomed Pharmacother. 2019;112:18. doi:10.1016/j.biopha.2019.01.041.
- Saber R, Zadeh M, Pakanati KC, Bere P, Klaenhammer T, Mohamadzadeh M. Lipoteichoic acid-deficient Lactobacillus acidophilus regulates downstream signals. Immunotherapy. 2011;3:337–347. doi:10.2217/imt.10.119.
- Wang H, Cheng X, Zhang L, Xu S, Zhang Q, Lu R. A surface-layer protein from Lactobacillus acidophilus NCFM induces autophagic death in HCT116 cells requiring ROS-mediated modulation of mTOR and JNK signaling pathways. Food Funct. 2019;10:4102–4112. doi:10.1039/C9FO00109C.
- Rahbar Saadat Y, Yari Khosroushahi A, Movassaghpour AA, Talebi M, Pourghassem Gargari B. Modulatory role of exopolysaccharides of Kluyveromyces marxianus and Pichia kudriavzevii as probiotic yeasts from dairy products in human colon cancer cells. J Funct Foods. 2020;64:103675. doi:10.1016/j.jff.2019.103675.
- Han N, Jia L, Su Y, Du J, Guo L, Luo Z, Liu Y. Lactobacillus reuteri extracts promoted wound healing via PI3K/AKT/β-catenin/TGFβ1 pathway. Stem Cell Res Ther. 2019;10:019–1324. doi:10.1186/s13287-019-1324-8.
- Ting WJ, Kuo WW, Kuo CH, Yeh YL, Shen CY, Chen YH, Ho T-J, Viswanadha VP, Chen Y-H, Huang C-Y, et al. Supplementary heat-killed Lactobacillus reuteri GMNL-263 ameliorates hyperlipidaemic and cardiac apoptosis in high-fat diet-fed hamsters to maintain cardiovascular function. Br J Nutr. 2015;114:706–712. doi:10.1017/S0007114515002469.
- Han C, Ding Z, Shi H, Qian W, Hou X, Lin R. The Role of Probiotics in Lipopolysaccharide-Induced Autophagy in Intestinal Epithelial Cells. Cell Physiol Biochem. 2016;38:2464–2478. doi:10.1159/000445597.
- Liu J, Sun J, Wang F, Yu X, Ling Z, Li H, et al. Neuroprotective Effects of Clostridium butyricum against Vascular Dementia in Mice via Metabolic Butyrate. Biomed Res Int. 2015;412946:7.
- Endo H, Niioka M, Kobayashi N, Tanaka M, Watanabe T. Butyrate-producing probiotics reduce nonalcoholic fatty liver disease progression in rats: new insight into the probiotics for the gut-liver axis. PLoS One. 2013;8:e63388. doi:10.1371/journal.pone.0063388.
- Sun L, Tian W, Guo X, Zhang Y, Liu X, Li X, Tian Y, Man C, Jiang Y. Lactobacillus gasseri JM1 with potential probiotic characteristics alleviates inflammatory response by activating the PI3K/Akt signaling pathway in vitro. J Dairy Sci. 2020;103:7851–7864. doi:10.3168/jds.2020-18187.
- Li X, Wu Q, Sui Y, Li S, Xie B, Sun Z. Dietary supplementation of A-type procyanidins from litchi pericarp improves glucose homeostasis by modulating mTOR signaling and oxidative stress in diabetic ICR mice. J Funct Foods. 2018;44:155–165. doi:10.1016/j.jff.2017.12.024.
- Xu M, Fu L, Zhang J, Wang T, Fan J, Zhu B, Dziugan P, Zhang B, Zhao H. Potential of Inactivated Bifidobacterium Strain in Attenuating Benzo(A)Pyrene Exposure-Induced Damage in Colon Epithelial Cells In Vitro. Toxics. 2020;8:12. doi:10.3390/toxics8010012.
- Toda K, Yamauchi Y, Tanaka A, Kuhara T, Odamaki T, Yoshimoto S, Xiao J-Z. Heat-Killed Bifidobacterium breve B-3 Enhances Muscle Functions: possible Involvement of Increases in Muscle Mass and Mitochondrial Biogenesis. Nutrients. 2020;12:219. doi:10.3390/nu12010219.
- Hoarau C, Martin L, Faugaret D, Baron C, Dauba A, Aubert-Jacquin C, Velge-Roussel F, Lebranchu Y. Supernatant from bifidobacterium differentially modulates transduction signaling pathways for biological functions of human dendritic cells. PLoS One. 2008;3:0002753. doi:10.1371/journal.pone.0002753.
- Kling DN, DeBose-Scarlett EM, Teixeira LD, Gezan SA, Lorca GL, Gonzalez CF. Sex Modulates Lactobacillus johnsonii N6.2 and Phytophenol Effectiveness in Reducing High Fat Diet Induced mTOR Activation in Sprague-Dawley Rats. Front Microbiol. 2018;9. doi:10.3389/fmicb.2018.02649.
- Hwang JW, Baek YM, Yang KE, Yoo HS, Cho CK, Lee YW, Park J, Eom C-Y, Lee Z-W, Choi J-S, et al. Lactobacillus casei Extract Induces Apoptosis in Gastric Cancer by Inhibiting NF-κB and mTOR-Mediated Signaling. Integr Cancer Ther. 2013;12:165–173. doi:10.1177/1534735412442380.
- Huang L, Shan Y-J, He C-X, Ren M-H, Tian P-J, Song W. Effects of L. paracasei subp. paracasei X12 on cell cycle of colon cancer HT-29 cells and regulation of mTOR signalling pathway. J Funct Foods. 2016;21:431–439. doi:10.1016/j.jff.2015.12.024.
- Dang F, Jiang Y, Pan R, Zhou Y, Wu S, Wang R, Zhuang K, Zhang W, Li T, Man C, et al. Administration of Lactobacillus paracasei ameliorates type 2 diabetes in mice. Food Funct. 2018;9:3630–3639. doi:10.1039/C8FO00081F.
- Dai C, Zheng CQ, Meng FJ, Zhou Z, Sang LX, Jiang M. VSL#3 probiotics exerts the anti-inflammatory activity via PI3k/Akt and NF-κB pathway in rat model of DSS-induced colitis. Mol Cell Biochem. 2013;374:1–11. doi:10.1007/s11010-012-1488-3.
- Zununi Vahed S, Barzegari A, Rahbar Saadat Y, Goreyshi A, Omidi Y. Leuconostoc mesenteroides-derived anticancer pharmaceuticals hinder inflammation and cell survival in colon cancer cells by modulating NF-κB/AKT/PTEN/MAPK pathways. Biomed Pharmacother. 2017;94:1094–1100. doi:10.1016/j.biopha.2017.08.033.
- Dong Y, Zhu J, Zhang M, Ge S, Zhao L. Probiotic Lactobacillus salivarius Ren prevent dimethylhydrazine-induced colorectal cancer through protein kinase B inhibition. Appl Microbiol Biotechnol. 2020;104:7377–7389. doi:10.1007/s00253-020-10775-w.
- Jia G, Liu X, Che N, Xia Y, Wang G, Xiong Z, Zhang H, Ai L. Human-origin Lactobacillus salivarius AR809 protects against immunosuppression in S. aureus-induced pharyngitis via Akt-mediated NF-κB and autophagy signaling pathways. Food Funct. 2020;11:270–284. doi:10.1039/C9FO02476J.
- Jeong JJ, Woo JY, Kim KA, Han MJ, Kim DH. Lactobacillus pentosus var. plantarum C29 ameliorates age-dependent memory impairment in Fischer 344 rats. Lett Appl Microbiol. 2015;60:307–314. doi:10.1111/lam.12393.
- Singh BN, Kumar D, Shankar S, Srivastava RK. Rottlerin induces autophagy which leads to apoptotic cell death through inhibition of PI3K/Akt/mTOR pathway in human pancreatic cancer stem cells. Biochem Pharmacol. 2012;84:1154–1163. doi:10.1016/j.bcp.2012.08.007.
- Levine B, Mizushima N, Virgin HW. Autophagy in immunity and inflammation. Nature. 2011;469:323–335. doi:10.1038/nature09782.
- Erickson BJ, Bach BR, Cohen MS, Bush-Joseph CA, Cole BJ, Verma NN, et al. Ulnar Collateral Ligament Reconstruction: the Rush Experience. Orthop J Sports Med. 2016;4:2325967115626876. doi:10.1177/2325967115626876.
- Heras-Sandoval D, Pérez-Rojas JM, Hernández-Damián J, Pedraza-Chaverri J. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal. 2014;26:2694–2701. doi:10.1016/j.cellsig.2014.08.019.
- Zhou YY, Li Y, Jiang WQ, Zhou LF. MAPK/JNK signalling: a potential autophagy regulation pathway. Biosci Rep. 2015;35. doi:10.1042/BSR20140141.
- Chiarini F, Evangelisti C, McCubrey JA, Martelli AM. Current treatment strategies for inhibiting mTOR in cancer. Trends Pharmacol Sci. 2015;36:124–135. doi:10.1016/j.tips.2014.11.004.
- Yu X, Long YC, Shen HM. Differential regulatory functions of three classes of phosphatidylinositol and phosphoinositide 3-kinases in autophagy. Autophagy. 2015;11:1711–1728. doi:10.1080/15548627.2015.1043076.
- Mizushima N. The role of the Atg1/ULK1 complex in autophagy regulation. Curr Opin Cell Biol. 2010;22:132–139. doi:10.1016/j.ceb.2009.12.004.
- Singh R, Kaushik S, Wang Y, Xiang Y, Novak I, Komatsu M, Tanaka K, Cuervo AM, Czaja MJ. Autophagy regulates lipid metabolism. Nature. 2009;458:1131–1135. doi:10.1038/nature07976.
- Ghadimi D, Herrmann J, De Vrese M, Heller KJ. Commensal lactic acid-producing bacteria affect host cellular lipid metabolism through various cellular metabolic pathways: role of mTOR, FOXO1, and autophagy machinery system. PharmaNutrition. 2018;6:215–235. doi:10.1016/j.phanu.2018.10.004.
- ON C. PI3-kinase/Akt/mTOR signaling: impaired on/off switches in aging, cognitive decline and Alzheimer’s disease. Exp Gerontol. 2013;48:647–653. doi:10.1016/j.exger.2013.02.025.
- Pratiwi YS, Lesmana R, Goenawan H, Sylviana N, Setiawan I, Tarawan VM, et al. Nutmeg Extract Increases Skeletal Muscle Mass in Aging Rats Partly via IGF1-AKT-mTOR Pathway and Inhibition of Autophagy. Evid Based Complement Alternat Med. 2018;2018:2810840. doi:10.1155/2018/2810840.
- Mannick JB, Del Giudice G, Lattanzi M, Valiante NM, Praestgaard J, Huang B, Lonetto MA, Maecker HT, Kovarik J, Carson S, et al. mTOR inhibition improves immune function in the elderly. Sci Transl Med. 2014;6:3009892. doi:10.1126/scitranslmed.3009892.
- Sui X, Kong N, Ye L, Han W, Zhou J, Zhang Q, He C, Pan H. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014;344:174–179. doi:10.1016/j.canlet.2013.11.019.
- Mi Y, Xiao C, Du Q, Wu W, Qi G, Liu X. Momordin Ic couples apoptosis with autophagy in human hepatoblastoma cancer cells by reactive oxygen species (ROS)-mediated PI3K/Akt and MAPK signaling pathways. Free Radic Biol Med. 2016;90:230–242. doi:10.1016/j.freeradbiomed.2015.11.022.
- Snyder HM, Corriveau RA, Craft S, Faber JE, Greenberg SM, Knopman D, Lamb BT, Montine TJ, Nedergaard M, Schaffer CB, et al. Vascular contributions to cognitive impairment and dementia including Alzheimer’s disease. Alzheimers Dement. 2015;11:710–717. doi:10.1016/j.jalz.2014.10.008.
- Téglás T, Ábrahám D, Jókai M, Kondo S, Mohammadi R, Fehér J, Szabó D, Wilhelm M, Radák Z. Exercise combined with a probiotics treatment alters the microbiome, but moderately affects signalling pathways in the liver of male APP/PS1 transgenic mice. Biogerontology. 2020;21:807–815. doi:10.1007/s10522-020-09895-7.
- Johnson BR, O’Flaherty S, Goh YJ, Carroll I, Barrangou R, Klaenhammer TR. The S-layer Associated Serine Protease Homolog PrtX Impacts Cell Surface-Mediated Microbe-Host Interactions of Lactobacillus acidophilus NCFM. Front Microbiol. 2017;8. doi:10.3389/fmicb.2017.01185.
- Su SC, Hua KF, Lee H, Chao LK, Tan SK, Yang SF, Yang S-F, Hsu H-Y. LTA and LPS mediated activation of protein kinases in the regulation of inflammatory cytokines expression in macrophages. Clin Chim Acta. 2006;374:106–115. doi:10.1016/j.cca.2006.05.045.
- Kaji R, Kiyoshima-Shibata J, Nagaoka M, Nanno M, Shida K. Bacterial teichoic acids reverse predominant IL-12 production induced by certain lactobacillus strains into predominant IL-10 production via TLR2-dependent ERK activation in macrophages. J Immunol. 2010;184:3505–3513. doi:10.4049/jimmunol.0901569.
- Park OJ, Yang J, Kim J, Yun CH, Han SH. Enterococcus faecalis attenuates the differentiation of macrophages into osteoclasts. J Endod. 2015;41:658–662. doi:10.1016/j.joen.2014.12.015.
- Groschwitz KR, Hogan SP. Intestinal barrier function: molecular regulation and disease pathogenesis. J Allergy Clin Immunol. 2009;124:3–20. doi:10.1016/j.jaci.2009.05.038.
- Sun Z, Wang X, Wallen R, Deng X, Du X, Hallberg E, Andersson R. The influence of apoptosis on intestinal barrier integrity in rats. Scand J Gastroenterol. 1998;33:415–422. doi:10.1080/00365529850171053.
- Chin AC, Teoh DA, Scott KG, Meddings JB, Macnaughton WK, Buret AG. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infect Immun. 2002;70:3673–3680. doi:10.1128/IAI.70.7.3673-3680.2002.
- Wang Y, Kirpich I, Liu Y, Ma Z, Barve S, McClain CJ, Feng W. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Am J Pathol. 2011;179:2866–2875. doi:10.1016/j.ajpath.2011.08.039.
- Will M, Qin AC, Toy W, Yao Z, Rodrik-Outmezguine V, Schneider C, Huang X, Monian P, Jiang X, De Stanchina E, et al. Rapid induction of apoptosis by PI3K inhibitors is dependent upon their transient inhibition of RAS-ERK signaling. Cancer Discov. 2014;4:334–347. doi:10.1158/2159-8290.CD-13-0611.
- Johnson SM, Gulhati P, Rampy BA, Han Y, Rychahou PG, Doan HQ, Weiss HL, Evers BM. Novel expression patterns of PI3K/Akt/mTOR signaling pathway components in colorectal cancer. J Am Coll Surg. 2010;210:767–776. doi:10.1016/j.jamcollsurg.2009.12.008.
- Zhang Y, Hou Q, Ma C, Zhao J, Xu H, Li W, Wang Y, Ma H, Zhang H, Sun Z, et al. Lactobacillus casei protects dextran sodium sulfate- or rapamycin-induced colonic inflammation in the mouse. Eur J Nutr. 2020;59:1443–1451. doi:10.1007/s00394-019-02001-9.
- Wilson PW, D’Agostino RB, Parise H, Sullivan L, Meigs JB. Metabolic syndrome as a precursor of cardiovascular disease and type 2 diabetes mellitus. Circulation. 2005;112:3066–3072. doi:10.1161/CIRCULATIONAHA.105.539528.
- White MF. IRS proteins and the common path to diabetes. Am J Physiol Endocrinol Metab. 2002;283:E413–E422. doi:10.1152/ajpendo.00514.2001.
- Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell. 2012;149:274–293. doi:10.1016/j.cell.2012.03.017.
- Li X, Sui Y, Wu Q, Xie B, Sun Z. Attenuated mTOR Signaling and Enhanced Glucose Homeostasis by Dietary Supplementation with Lotus Seedpod Oligomeric Procyanidins in Streptozotocin (STZ)-Induced Diabetic Mice. J Agric Food Chem. 2017;65:3801–3810. doi:10.1021/acs.jafc.7b00233.
- Le TK, Hosaka T, Nguyen TT, Kassu A, Dang TO, Tran HB, PHAM TP, TRAN QB, LE THH, PHAM XD, et al. Bifidobacterium species lower serum glucose, increase expressions of insulin signaling proteins, and improve adipokine profile in diabetic mice . Biomed Res. 2015;36:63–70. doi:10.2220/biomedres.36.63.
- Wang Y, Dilidaxi D, Wu Y, Sailike J, Sun X, Nabi XH. Composite probiotics alleviate type 2 diabetes by regulating intestinal microbiota and inducing GLP-1 secretion in db/db mice. Biomed Pharmacother. 2020;125:5. doi:10.1016/j.biopha.2020.109914.
- Torella D, Rota M, Nurzynska D, Musso E, Monsen A, Shiraishi I, Zias E, Walsh K, Rosenzweig A, Sussman MA, et al. Cardiac stem cell and myocyte aging, heart failure, and insulin-like growth factor-1 overexpression. Circ Res. 2004;94:514–524. doi:10.1161/01.RES.0000117306.10142.50.
- Lin PP, Hsieh YM, Kuo WW, Lin YM, Yeh YL, Lin CC, TSAI F-J, TSAI C-H, HUANG C-Y, TSAI -C-C, et al. Probiotic-fermented purple sweet potato yogurt activates compensatory IGF‑IR/PI3K/Akt survival pathways and attenuates cardiac apoptosis in the hearts of spontaneously hypertensive rats. Int J Mol Med. 2013;32:1319–1328. doi:10.3892/ijmm.2013.1524.
- Wang HF, Lin PP, Chen CH, Yeh YL, Huang CC, Huang CY, TSAI -C-C. Effects of lactic acid bacteria on cardiac apoptosis are mediated by activation of the phosphatidylinositol-3 kinase/AKT survival-signalling pathway in rats fed a high-fat diet. Int J Mol Med. 2015;35:460–470. doi:10.3892/ijmm.2014.2021.
- Henao-Mejia J, Elinav E, Jin C, Hao L, Mehal WZ, Strowig T, Thaiss CA, Kau AL, Eisenbarth SC, Jurczak MJ, et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature. 2012;482:179–185. doi:10.1038/nature10809.
- Hashimoto E, Taniai M, Tokushige K. Characteristics and diagnosis of NAFLD/NASH. J Gastroenterol Hepatol. 2013;4:64–70. doi:10.1111/jgh.12271.
- Lin R, Wu D, Wu FJ, Meng Y, Zhang JH, Wang XG, Jia L-H. Non-alcoholic Fatty Liver Disease Induced by Perinatal Exposure to Bisphenol a Is Associated With Activated mTOR and TLR4/NF-κB Signaling Pathways in Offspring Rats. Front Endocrinol. 2019;10. doi:10.3389/fendo.2019.00620.
- Fang L, Wu HM, Ding PS, Liu RY. TLR2 mediates phagocytosis and autophagy through JNK signaling pathway in Staphylococcus aureus-stimulated RAW264.7 cells. Cell Signal. 2014;26:806–814. doi:10.1016/j.cellsig.2013.12.016.
- Zha L, Chen J, Sun S, Mao L, Chu X, Deng H, Cai J, Li X, Liu Z, Cao W, et al. Soyasaponins can blunt inflammation by inhibiting the reactive oxygen species-mediated activation of PI3K/Akt/NF-kB pathway. PLoS One. 2014;9:e107655. doi:10.1371/journal.pone.0107655.
- Kalliomäki M, Salminen S, Poussa T, Arvilommi H, Isolauri E. Probiotics and prevention of atopic disease: 4-year follow-up of a randomised placebo-controlled trial. Lancet. 2003;361:1869–1871. doi:10.1016/S0140-6736(03)13490-3.
- Sokol CL, Barton GM, Farr AG, Medzhitov R. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nat Immunol. 2008;9:310–318. doi:10.1038/ni1558.
- Brenner M, Hearing VJ. The protective role of melanin against UV damage in human skin. Photochem Photobiol. 2008;84:539–549. doi:10.1111/j.1751-1097.2007.00226.x.
- Chamcheu JC, Adhami VM, Esnault S, Sechi M, Siddiqui IA, Satyshur KA, Syed DN, Dodwad SJM, Chaves-Rodriquez M-I, Longley BJ, et al. Dual Inhibition of PI3K/Akt and mTOR by the Dietary Antioxidant, Delphinidin, Ameliorates Psoriatic Features In Vitro and in an Imiquimod-Induced Psoriasis-Like Disease in Mice. Antioxid Redox Signal. 2017;26:49–69. doi:10.1089/ars.2016.6769.
- Mohseni AH, Taghinezhad-Saroukalaei S, Voglmeir J. Recombinant Glycoenzyme Production in Gram-Positive Bacteria—An Overview. Trends Glycosci. Glycotechnol 2020;32:E99-E104. doi:10.4052/tigg.1824.1E.