
ABSTRACT
The fungus Candida albicans is a ubiquitous member of the human gut microbiota. Hundreds or thousands of bacterial taxa reside together with this fungus in the intestine, creating a milieu with myriad opportunities for inter-kingdom interactions. Indeed, recent studies examining the broader composition – that is, monitoring not only bacteria but also the often neglected fungal component – of the gut microbiota hint that there are significant interdependencies between fungi and bacteria. Gut bacteria closely associate with C. albicans cells in the colon, break down and feed on complex sugars decorating the fungal cell wall, and shape the intestinal microhabitats occupied by the fungus. Peptidoglycan subunits released by bacteria upon antibiotic treatment can promote C. albicans dissemination from the intestine, seeding bloodstream infections that often become life-threatening. Elucidating the principles that govern the fungus-bacteria interplay may open the door to novel approaches to prevent C. albicans infections originating in the gut.
The intestinal microbiota is a consortium of microorganisms including bacteria, fungi, protists, archaea, and viruses. Although the vast majority of studies in the microbiota field are still focused on the bacterial component, the last decade has seen a steady increase of research featuring the biology and critical importance of non-bacterial members of this microbial community.Citation1–5Gut commensal fungi (reviewed in Ref. 5) have become a subject of particular interest because recent studies are revealing beneficial effects of fungal colonization for the mammalian host including resistance to pathogens and tuning of the immune system,Citation6,Citation7 as well as roles in human diseases such as inflammatory disorders and specific cancers.Citation8 Yet neither fungi nor any other microbial taxa are likely to act alone in such a crowded ecosystem. The fact that fungi cohabit with bacteria in the human intestine rather implies that myriad interactions occur between members of these two kingdoms. In this review, I highlight current developments in the study of fungal–bacterial interactions that take place in the mammalian gut. While research looking at the entire gut fungal community is outlined in some sections of the paper, the focus of the review is on the most prevalent fungus in the human intestinal tract, Candida albicans.Citation9
Interdependencies between bacteria and fungi in the mammalian gut
There is a large body of literature examining how particular bacteria modulate fungal traits (e.g. in C. albicans) and vice versa. However, the vast majority of the data on bacteria–fungi interactions have been collected using either in vitro systems (e.g. biofilm formation) or non-intestinal mucosal settings (e.g. oral or vaginal). Much of this literature is neatly summarized in recent, comprehensive reviewsCitation10–13and will not be detailed here. A fundamental question that should be asked, however, is whether the interactions and effects that have been documented under in vitro settings (e.g. co-culture under laboratory conditions) reflect in any way what takes place in the intestinal tract. While the paucity of mechanistic studies in well-defined mammalian animal models precludes an answer to this question in many specific cases, broader analyses and observations clearly support the notion that there are interdependencies between fungi and bacteria in the gut. The interdependencies can be seen both ways: Bacteria impinging upon fungi and vice versa.
Eliminating bacteria through antibiotic treatment results in fungal dysbiosis
The yeast C. albicans is a dweller of the human gastrointestinal tract (reviewed in Ref. 13) along with hundreds (or even thousands) of bacterial taxa. As early as in the 1960s it was observed that antibiotic treatment in humans resulted in C. albicans overgrowth,Citation14,Citation15 presumably due to the dampening of competing microbes. More recent studies equipped with the tools to monitor the entire intestinal fungal community have revealed that antibiotic treatments that target bacteria inadvertently alter the fungal composition as well.Citation16,Citation17 Thus, fungal dysbiosis can result from eliminating or reducing gut bacteria.
Changes in gut fungal communities lead to bacterial dysbiosis
Evidence is also accumulating that targeting fungi can lead to changes in bacterial communities. For instance, in one of these studies mice treated with antifungal drugs exhibited pronounced alterations in the composition of their bacterial community.Citation18 In another study in gnotobiotic mice, a small community of fungi (five species) was found to induce strong ecological changes in the assembly of gut bacteria.Citation19 Candida albicans itself has been shown to impact the reassembly of gut bacterial communities after antibiotic treatment.Citation20,Citation21 The fungus also has a role in the ability of intestinal microbial consortia to resist change in response to antibiotic disruption, which is termed ecological resistance.Citation22 Furthermore, gut fungal dysbiosis has been associated with reduced efficacy in fecal transplants currently in use to treat recurrent Clostridium difficile infections.Citation23
In a longitudinal study of a cohort of 178 preterm infants in which the assembly of the intestinal microbiota was monitored, Rao et al.Citation24 were able to infer several interdependencies between specific fungi and bacterial taxa. For instance, they inferred that C. albicans antagonizes Klebsiella pneumoniae whereas Staphylococcus spp. may antagonize Candida. Some of the predicted pairwise interactions were further confirmed in gut colonization experiments in rodents indicating that, at least in microbial communities of relatively low complexity such as the infant gut, it may be possible to single out significant fungus-bacteria interactions. Another large study (942 human adults) found a positive correlation between fungal and bacterial richness in the gut, with is suggestive of mutualism between both kingdoms, yet failed to uncover correlations between fungi and bacteria at the species level.Citation25 Taken together, the studies described in this section, while short of providing mechanistic details, do indicate that fungi and bacteria impact on the proliferation of each other in the mammalian gut.
The yeast Candida albicans is a pathobiont of the human intestine
Several yeasts of the genus Candida are considered true symbionts of the human gut.Citation26 Candida albicans, however, is the species of the genus Candida most frequently detected in feces of healthy humans.Citation27–29 This species, therefore, is considered a ubiquitous member of the human gut microbiota. Candida albicans appears to have no major environmental reservoir, suggesting that it has extensively coevolved with humans and cohabiting microbes. The fungus can be found in other body sites beyond the intestine (e.g. mouth, skin, vagina) and is a common cause of fastidious mucosal disease in otherwise healthy people.Citation30 C. albicans can also disseminate from the human gut into the bloodstream and invade internal organs producing invasive, life-threatening infections.Citation31,Citation32 Invasive Candida infections rank in the top four hospital-associated bloodstream infections in the United States.Citation33,Citation34 Common risk factors for invasive candidiasis include intensive care unit stay, central venous catheter use, broad-spectrum antibiotics, recent abdominal surgery and immune suppression.Citation33,Citation35
Candida albicans cells can adopt a variety of specialized morphologies that considerably differ from one another in gene expression profile, cell surface composition and how they interact with host cells.Citation36–40 Two of the most readily found C. albicans morphologies are the oval-shaped ‘yeast’ form and the long filaments termed ‘hyphae.’ The yeast and hyphae morphologies are not only easy to score but also the transition between both forms is critical for the fungus to invade host tissues and cause disease (see Ref. 40 for a recent review). The yeast-to-hyphae transition can be driven by numerous environmental signals, including temperature, presence of serum and CO2, among others. As described in the following sections, many studies examining whether bacteria impinge upon the biology of C. albicans have looked at effects on the yeast-to-hyphae transition.
While less studied in the context of microbial interactions, other C. albicans cell morphologies such as white/opaqueCitation41 and GUT (gastrointestinally induced transition)Citation42 may also be relevant for the fungus in the intestine. The WOR1 transcript, which encodes the “master” regulator of white-opaque switching, was strongly upregulated by in vitro co-culture with the intestinal bacteria K. pneumoniae, Escherichia coli, and Enterococcus faecalis.Citation43 In addition to WOR1, the authors found a number of opaque-specific genes also upregulated in the co-culture experiment, yet the full opaque-specific gene expression pattern was not observed.Citation43 Employing a C. albicans strain genetically modified to overproduce WOR1, another group reported that in the murine intestine the fungus adopted a morphology resembling – but distinct from – that of opaque cells.Citation42 The transition to these so-called GUT cells was postulated to confer enhanced fitness in the mammalian gastrointestinal tract.Citation42
Direct effects of bacterial products on C. albicans
Gut bacteria could influence the proliferation of C. albicans or other fungi (or impact any other fungal trait) by at least two broad mechanisms: One is the release of molecules such as cell surface components, peptides or metabolic products that can directly impact the biology of the fungus. An alternative scenario involves an indirect route in which bacterial molecules (or the microbes themselves) elicit responses from host cells; in this scenario, the microbe-induced host response is what ultimately targets the fungus. I detail here a few examples illustrating the former mechanism and will cover the latter scenario in the following section.
Some of the most recognizable bacterial components are those found in their outer membrane and cell wall. Molecules derived from these structures are also among the most abundant microbial products in the gut.Citation44 Peptidoglycan, for example, is a major component of the bacterial cell wall. It consists of alternating β-(1,4)-linked N-acetylglucosamine and N-acetylmuramic acid cross-linked by regularly spaced short peptides. Certain bacterial peptidoglycan subunits such as 1,6-anhydro-N-acetylmuramyl peptides are strong hypha-inducing agents in C. albicans.Citation45 This is consistent with the observation that the amino sugar N-acetylglucosamine also promotes hyphal morphogenesis.Citation46,Citation47 In a recent study in mice,Citation48 it was shown that β-lactam antibiotic treatment promoted C. albicans hyphal growth in the gastrointestinal tract due to the release of bacterial peptidoglycan subunits in the intestinal lumen (). Furthermore, the authors established that the filamenting C. albicans cells under these conditions could disseminate from the gut causing invasive candidiasis. The β-lactam mediated effect on the fungus could explain, at least in part, the general observation that antibiotic treatment constitutes a major risk factor for Candida dissemination from the gut.
Figure 1. The fungus C. albicans inhabits the mammalian gut along with numerous and diverse bacterial species. Cartoon depicts the mammalian colon. The inner mucus layer which is largely devoid of microbes separates intestinal epithelial cells from the microbiota. C. albicans cells adopt the oval-shaped ‘yeast’ morphology and occupy the outer mucus layer as well as the intestinal lumen. Illustrated are four instances of documented bacterial interactions with C. albicans: (a) β-lactam antibiotic treatment promotes C. albicans filamentation due to the release of bacterial peptidoglycan subunits in the intestinal lumen. Candida hyphae disseminate from the gut seeding bloodstream infections. (b) The bacterium E. faecalis secretes the EntV peptide which inhibits hyphal morphogenesis. (c) Multiple bacterial species can provide a wide range of metabolic products which can alter C. albicans proliferation, albeit by unknown mechanisms. And (d) Bacteroides spp. closely associate with C. albicans cells in the outer mucus layer and can feed on mannan, a complex carbohydrate decorating the cell surface of the fungus. See main text for further details
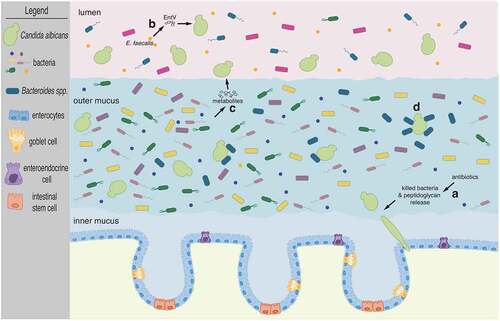
Bacteria often secrete peptides and/or toxins that can act on nearby cells. Enterococcus faecalis, a Gram-positive, commensal bacterium inhabiting the gastrointestinal tract of humans, produces a 170-amino-acid prepropeptide which is processed by several enzymes to generate and secrete an active 68-amino-acid peptide known as EntV.Citation49 The secreted peptide has been shown to inhibit the formation of C. albicans filaments (), preventing biofilm formation.Citation50 While it remains to be established whether EntV has any role in the interplay between these two species in their natural habitat [i.e. within the mammalian intestine] this study is significant because it established the molecular basis of an antagonistic relationship between E. faecalis and C. albicans which had originally been discovered in a co-infection model.Citation51
Metabolic products derived from gut bacteria may be rationalized as the most likely molecules directly affecting C. albicans proliferation in the intestine (). Indeed, metabolites produced by a consortium of 60 bacterial strains derived from human feces and grown in a continuous-culture bioreactor system inhibited to some extent the proliferation of C. albicans in liquid culture and also its ability to filament.Citation52 The identity of these metabolites and the specific bacterial species producing them, however, remains to be established. Short-chain fatty acids (SCFAs) [in particular acetic, butyric and propionic acid] have been shown to have multiple effects on C. albicans under in vitro culture conditions: SCFAs inhibit germ tub formation, reduce metabolic activity in biofilms and impair growth.Citation53 Acetic, propionic and cis-5-dodecenoic acid also reduce C. albicans-induced damage of cultured human epithelial cells.Citation16 While metabolites produced by bacteria not typically associated with the gut, such as Pseudomonas aeruginosa, have been shown to directly target C. albicans functions,Citation54,Citation55 there is still a significant gap in our knowledge of metabolic products released by bacteria in the intestinal milieu that directly influence intestinal colonization by C. albicans.
While the majority of studies on bacterial–fungal interactions have looked on bacteria having effects on the fungus, it is important to notice that C. albicans can also have a significant influence on co-habiting bacteria. For instance, Candida enhances the in vitro growth of the strict anaerobes Bacteroides fragilis and Bacteroides vulgatus.Citation56 The authors of this study observed similar effects when supplied spent C. albicans medium or dead C. albicans cells, which led them to postulate that carbohydrates located on the fungal surface could be fueling bacterial growth. Consistent with this hypothesis, Bacteroides spp. have been shown to harbor enzymes that are specialized in breaking down the sugar mannan,Citation57 which is abundant on Candida’s surface. The simpler sugars resulting from the mannan digestion by the Bacteroides enzymes fuel the proliferation of Bacteroides.
The examples outlined in this section clearly indicate that gut bacteria-derived molecules, including cell wall components, peptides, and metabolic products, directly impinge upon multiple C. albicans traits including its proliferation in the intestine. Likewise, molecules derived from the fungus can also have a significant influence on co-habiting bacteria.
Host-mediated effects of bacteria on C. albicans gut colonization
Priming responses by the host’s immune system is a well-established mechanism whereby commensal bacteria protect the mammalian host from invading pathogens.Citation58 This ‘host-mediated’ or ‘indirect’ mechanism has often been invoked in the context of bacteria–bacteria antagonistic relationships. Here I briefly describe two studies that have examined the basis of antagonistic relationships between bacteria and C. albicans in murine models of intestinal colonization and that indicate that the host’s immune system is involved. Such evidence suggests that the concept of ‘host-mediated’ interactions can be extended to cases of fungus-bacteria interplay as well.
In one of the studies,Citation59 C. albicans was found to protect against lethal murine Clostridium difficile infections. The effect appeared to be mediated, at least in part, by the fungus promoting the production of IL-17, a pro-inflammatory cytokine. In the other study,Citation60 several Bacteroidetes and clostridial Firmicutes were found to antagonize C. albicans in the murine intestine. The authors convincingly showed that these bacteria restricted proliferation of the fungus by stimulating the production of gut mucosal immune defenses, in particular the production of the antimicrobial peptide CRAMP, which targeted C. albicans. It is noteworthy that, based on the two studies described here, C. albicans has dual roles: First, as a commensal that can elicit a response to protect the host from a bacterial pathogen; and, second, as a fungal pathogen from which the host is protected via immune responses elicited by commensal bacteria.
In addition to the two cases described in the previous paragraph, it is worth mentioning a study in a model of recurrent mucosal Candida infection in antibiotic-treated mice.Citation61 The authors found that SCFA administration in these animals resulted in fungal clearance and inflammation resolution. Antibiotic administration dramatically reduced the intestinal regulatory T cell population (Foxp3+ Tregs) whereas SCFA treatment partially restored its numbers. Regulatory T cells, therefore, may also be part of the bacteria-modulated host cells that shape responses to C. albicans.
Gut bacteria shape the intestinal microhabitats occupied by C. albicans
Gut microbes inhabit a variety of distinct microhabitats along the longitudinal and cross-sectional axes of the intestinal tract.Citation62 Along the intestine, mucus provides a physical barrier that separates the gut lumen from the intestinal epithelial cells.Citation63 The mucus layer is a gel-like structure that represents a niche vastly different from the adjacent lumen.Citation64–66 C. albicans has been imaged throughout the digestive tract (from stomach to small and large intestine) in both gnotobiotic and antibiotic-treated conventional mice.Citation67,Citation68 High-resolution microscopy analyses conducted with colon sections derived from gnotobiotic mice co-colonized with C. albicans and single bacterial species indicate that a significant proportion of fungal cells (roughly 50%) localize to the outer mucus.Citation69 Fungal (in the round ‘yeast’ form) and bacterial cells (Bacteroides thetaiotaomicron in particular) were visualized in close association within the mucus (). In this experimental system, the formation of a substantial outer mucus layer was dependent on the presence of B. thetaiotaomicron suggesting that the spatial distribution of C. albicans, at least in the colon, is shaped by gut bacteria.
The partition of C. albicans in at least two distinct compartments, one luminal and another inside the mucus layer, raises the possibility that they represent disparate cell populations, each expressing a different set of genes. That is, the fungus could up- or down-regulate genes (to turn on or turn off associated biological functions) depending on whether it is in the lumen or mucus. In support of this notion, it has recently been shown that the gut bacteria B. fragilis expresses different sets of genes depending on the particular intestinal niche where the bacteria reside, namely lumen, mucus or closer to the epithelium.Citation70 Cell-to-cell variability in gene expression has been observed within C. albicans clonal populations. In fact, the expression of a major regulator of gut colonization, the transcription regulator Efg1p, exhibits extensive cell-to-cell variability.Citation71 It is tempting to speculate that such variability may, at least in part, be related to the particular microhabitat that the fungus occupies in the colon. Experimental approaches designed to carry out single-cell gene expression measurements spatially within specific gut microenvironments – such as the one described for B. fragilis in Ref. 70 – will allow researchers to explore these questions.
Metabolic niche occupied by C. albicans in the intestine
A fundamental question regarding C. albicans dwelling in the gut – and more generally about all fungi that reside in this organ – is what metabolic niche the species occupies. That is, what are the primary sources of key nutrients such as carbon and nitrogen that Candida depends on in the gut? And, at the other end, what metabolic products does Candida make available to nearby microbes? Answers to these questions could provide a basic framework to infer potential interdependencies of this fungus with specific gut bacteria. The metabolic niche(s) occupied by each member of the microbiota is shaped both by nutrient availability as well as the presence of other competing or cooperating species.Citation72 For example, bacteria of the genus Bacteroides are able to use a wide range of polysaccharidesCitation73,Citation74 and a number of closely‐related species can co‐exist in the gut by cross‐feeding, which may result in complete polysaccharide utilization.Citation75 While in silico metabolic modeling has been used to predict metabolic interactions between C. albicans and gut bacteria,Citation76 the niche of the fungus in the mammalian gastrointestinal tract remains largely undefined. (Based on exchange reaction fluxes, the in silico modelingCitation76 predicts, for example, that the amino acids proline and aspartate, and molecules such as nitrite and putrescine, constitute the top differences when it comes to consumption by fungus or bacteria.)
On the primary nutrient sources that C. albicans may use in the intestine, it has been argued that metabolic flexibility and mixed‐substrate utilization are common strategies for survival in the face of ever‐present nutrient fluctuations.Citation72 Metabolic flexibility is indeed a hallmark of C. albicans, and has been hypothesized to contribute to survival in the mammalian host, particularly in the gut.Citation77,Citation78 Transcriptome profiling experiments that have compared C. albicans cells growing inside the mouse intestine to cells cultured under laboratory conditions have revealed the upregulation of genes associated with translation, sugar transport and stress responses, among others.Citation79,Citation80 These findings point to the presence of metabolically active fungal cells that take up simple sugars (monosaccharides) in the cecum. C. albicans is unique among studied yeasts in that it can continuously assimilate nonpreferred carbon sources even in the presence of glucose.Citation78,Citation81 The mechanism for this phenomenon involves, at least in part, the rewiring of ubiquitination targetsCitation81 and is predicted to make the fungus more fit in an environment in which carbon availability is constantly changing in both quantity and quality.Citation78
On the metabolic products that the fungus may contribute to the intestinal milieu, a recent metabolome study comparing the gut content of antibiotic-treated mice carrying or not C. albicans found surprisingly few, if any, differences caused by the presence of the fungus.Citation82 The ~100 metabolites screened in this study included amino acids, carbohydrates, lipids, peptides, xenobiotics and bile acids. Another study found that the presence of C. albicans altered the levels of several lipid species in the mouse cecum, including non-esterified, unsaturated long-chain fatty acids.Citation83 Clearly, more comprehensive metabolome studies, ideally carried out in gnotobiotic animals, are needed to start delineating the metabolic footprint of the fungus in the intestine.
Potential for translational research
Candida albicans and related species such as C. parapsilosis are opportunistic pathogens. That is, they reside in the human intestinal tract as commensals, without causing any overt disease. Occasionally, however, and typically in individuals with debilitated defenses, these Candida species can disseminate from the intestine and cause deep-seated, invasive infections. Because they reside in the gut, it has been hypothesized that the intestinal microbiota may play a critical role in Candida’s pathogenesis. Recent clinical evidence supports this notion. In patients with disseminated Candida infections, C. albicans and C. parapsilosis translocation into the bloodstream was found to be preceded by an expansion of both species in the gastrointestinal tract.Citation31 Furthermore, fungal dysbiosis was found to be tightly associated with bacterial dysbiosis, particularly the loss of anaerobic bacteria.Citation31 These observations suggest that it may be possible to devise interventions that target the gut microbiome to inhibit Candida dissemination from the gastrointestinal tract in people at risk of infections. Elucidating the principles that govern the interplay between gut fungi and bacteria, therefore, has the potential to open the door to a novel way of treating and preventing infections: Rationally designed, microbiota-based therapeutics.Citation84 Studies on infections caused by the bacteria Vibrio choleraeCitation85 and Clostridium difficileCitation86 indicate that such approaches may indeed be feasible.
Conclusions and Outlook
There is growing evidence that significant interactions take place between fungi and bacteria in the mammalian gastrointestinal tract. Yet only limited progress has been made on the actual molecular mechanisms underlying specific interactions. The enormous complexity of the mammalian gut microbiota as well as the lack of a convenient and tractable animal model of gut fungal colonization may account for the paucity of mechanistic studies dissecting physiologically relevant interactions directly in the intestine. Nevertheless, future studies exploring the interplay between C. albicans and other members of the microbiota are likely to reveal novel insights into inter-kingdom microbial interactions that occur in our mucosal surfaces; bacterial vs. fungal strategies for colonizing the mammalian gut; and, rational microbiota-targeted approaches to prevent and treat infections.
Acknowledgments
I thank Hailey Nussbaum for the artwork employed in .
DATA AVAILABILITY STATEMENT
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Additional information
Funding
References
- Chudnovskiy A, Mortha A, Kana V, Kennard A, Ramirez JD, Rahman A, Remark R, Mogno I, Ng R, Gnjatic S, et al. Host-Protozoan Interactions Protect from Mucosal Infections through Activation of the Inflammasome. Cell. 2016;167(2):444–11. doi:10.1016/j.cell.2016.08.076.
- Huseyin CE, O’Toole PW, Cotter PD, Scanlan PD. Forgotten fungi-the gut mycobiome in human health and disease. FEMS Microbiol Rev. 2017;41:479–511.
- Iliev ID, Funari VA, Taylor KD, Nguyen Q, Reyes CN, Strom SP, Brown J, Becker CA, Fleshner PR, Dubinsky M, et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science. 2012;336:1314–1317.
- Li J, Chen D, Yu B, He J, Zheng P, Mao X, Yu J, Luo J, Tian G, Huang Z, et al. Fungi in Gastrointestinal Tracts of Human and Mice: from Community to Functions. Microb Ecol. 2018;75:821–829.
- Perez JC. Fungi of the human gut microbiota: roles and significance. Int J Med Microbiol. 2021;311:151490.
- Scheffold A, Bacher P, LeibundGut-Landmann S. T cell immunity to commensal fungi. Curr Opin Microbiol. 2020;58:116–123.
- Wiesner DL. Local farming of gut fungi protects against dangerous imports. Cell Host Microbe. 2021;29:311–312.
- Kong HH, Segre JA. Cultivating fungal research. Science. 2020;368:365–366.
- Romo JA, Kumamoto CA. On Commensalism of Candida. J Fungi. 2020;6:16.
- d’Enfert C, Kaune AK, Alaban LR, Chakraborty S, Cole N, Delavy M, Kosmala D, Marsaux B, Frois-Martins R, Morelli M, et al. The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: current knowledge and new perspectives. FEMS Microbiol Rev. 2021;45:fuaa060.
- Richard ML, Sokol H. The gut mycobiota: insights into analysis, environmental interactions and role in gastrointestinal diseases. Nat Rev Gastroenterol Hepatol. 2019;16:331–345.
- Santus W, Devlin JR, Crossing Kingdoms: BJ. How the Mycobiota and Fungal-Bacterial Interactions Impact Host Health and Disease. Infect Immun. 2021;89:e00648–20.
- Perez JC. Candida albicans dwelling in the mammalian gut. Curr Opin Microbiol. 2019;52:41–46.
- Seelig MS. Mechanisms by which antibiotics increase the incidence and severity of candidiasis and alter the immunological defenses. Bacteriol Rev. 1966;30:442–459.
- Seelig MS. The role of antibiotics in the pathogenesis of Candida infections. Am J Med. 1966;40:887–917.
- Seelbinder B, Chen J, Brunke S, Vazquez-Uribe R, Santhaman R, Meyer AC, de Oliveira Lino FS, Chan KF, Loos D, Imamovic L, et al. Antibiotics create a shift from mutualism to competition in human gut communities with a longer-lasting impact on fungi than bacteria. Microbiome. 2020;8:133.
- Sovran B, Planchais J, Jegou S, Straube M, Lamas B, Natividad JM, Agus A, Dupraz L, Glodt J, Da Costa G, et al. Enterobacteriaceae are essential for the modulation of colitis severity by fungi. Microbiome. 2018;6:152.
- Wheeler ML, Limon JJ, Bar AS, Leal CA, Gargus M, Tang J, Brown J, Funari VA, Wang HL, Crother TR, et al. Immunological Consequences of Intestinal Fungal Dysbiosis. Cell Host Microbe. 2016;19:865–873.
- van Tilburg Bernardes E, Pettersen VK, Gutierrez MW, Laforest-Lapointe I, Jendzjowsky NG, Cavin JB, Vicentini FA, Keenan CM, Ramay HR, Samara J, et al. Intestinal fungi are causally implicated in microbiome assembly and immune development in mice. Nat Commun. 2020;11:2577.
- Mason KL, Erb Downward JR, Falkowski NR, Young VB, Kao JY, Huffnagle GB. Interplay between the gastric bacterial microbiota and Candida albicans during postantibiotic recolonization and gastritis. Infect Immun. 2012;80:150–158.
- Erb Downward JR, Falkowski NR, Mason KL, Muraglia R, Huffnagle GB. Modulation of post-antibiotic bacterial community reassembly and host response by Candida albicans. Sci Rep. 2013;3:2191.
- Markey L, Pugliese A, Tian T, Roy F, Lee K, Kumamoto CA. Decreased Ecological Resistance of the Gut Microbiota in Response to Clindamycin Challenge in Mice Colonized with the Fungus Candida albicans. mSphere. 2021;6:e00982–20.
- Zuo T, Wong SH, Cheung CP, Lam K, Lui R, Cheung K, Zhang F, Tang W, Ching JYL, Wu JCY, et al. Gut fungal dysbiosis correlates with reduced efficacy of fecal microbiota transplantation in Clostridium difficile infection. Nat Commun. 2018;9:3663.
- Rao C, Coyte KZ, Bainter W, Geha RS, Martin CR, Rakoff-Nahoum S. Multi-kingdom ecological drivers of microbiota assembly in preterm infants. Nature. 2021;591:633–638.
- Sun Y, Zuo T, Cheung CP, Gu W, Wan Y, Zhang F, Chen N, Zhan H, Yeoh YK, Niu J, et al. Population-Level Configurations of Gut Mycobiome Across 6 Ethnicities in Urban and Rural China. Gastroenterology. 2021;160:272–286.
- Fiers WD, Gao IH, Iliev ID. Gut mycobiota under scrutiny: fungal symbionts or environmental transients? Curr Opin Microbiol. 2019;50:79–86.
- Hallen-Adams HE, Kachman SD, Kim J, Legge RM, Martinez I. Fungi inhabiting the healthy human gastrointestinal tract: a diverse and dynamic community. Fungal Ecol. 2015;15:9–17.
- Hoffmann C, Dollive S, Grunberg S, Chen J, Li H, Wu GD, Lewis JD, Bushman FD. Archaea and fungi of the human gut microbiome: correlations with diet and bacterial residents. PLoS One. 2013;8:e66019.
- Nash AK, Auchtung TA, Wong MC, Smith DP, Gesell JR, Ross MC, Stewart CJ, Metcalf GA, Muzny DM, Gibbs RA, et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome. 2017;5:153.
- Revankar SG, Sobel JD. Mucosal candidiasis. In: Calderone RA, Clancy CJ editors. Candida and Candidiasis. 2nd. Washington (DC): ASM Press; 2012. p. 419–427.
- Zhai B, Ola M, Rolling T, Tosini NL, Joshowitz S, Littmann ER, Amoretti LA, Fontana E, Wright RJ, Miranda E, et al. High-resolution mycobiota analysis reveals dynamic intestinal translocation preceding invasive candidiasis. Nat Med 2020;26:59–64.
- Koh AY, Kohler JR, Coggshall KT, Van Rooijen N, Pier GB. Mucosal damage and neutropenia are required for Candida albicans dissemination. PLoS Pathog. 2008;4:e35.
- Kullberg BJ, Arendrup MC. Invasive Candidiasis. N Engl J Med. 2015;373:1445–1456.
- Ricotta EE, Lai YL, Babiker A, Strich JR, Kadri SS, Lionakis MS, Prevots DR, Adjemian J. Invasive candidiasis species distribution and trends, United States, 2009–2017. J Infect Dis. 2020;223:1295–1302
- Pappas PG, Lionakis MS, Arendrup MC, Ostrosky-Zeichner L, Kullberg BJ. Invasive candidiasis. Nat Rev Dis Primers. 2018;4:18026.
- Moyes DL, Runglall M, Murciano C, Shen C, Nayar D, Thavaraj S, Kohli A, Islam A, Mora-Montes H, Challacombe SJ, et al. A biphasic innate immune MAPK response discriminates between the yeast and hyphal forms of Candida albicans in epithelial cells. Cell Host Microbe. 2010;8:225–235.
- Gow NA, Yadav B. Microbe Profile: candida albicans: a shape-changing, opportunistic pathogenic fungus of humans. Microbiology. 2017;163:1145–1147.
- Gow NA, Van De Veerdonk FL, Brown AJ, Netea MG. Candida albicans morphogenesis and host defence: discriminating invasion from colonization. Nat Rev Microbiol. 2012;10:112–122.
- Noble SM, Gianetti BA, Witchley JN. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nat Rev Microbiol. 2017;15:96–108.
- Kadosh D, Mundodi V. A Re-Evaluation of the Relationship between Morphology and Pathogenicity in Candida Species. J Fungi. 2020;6:13.
- Lohse MB, Johnson AD. White-opaque switching in Candida albicans. Curr Opin Microbiol. 2009;12:650–654.
- Pande K, Chen C, Noble SM. Passage through the mammalian gut triggers a phenotypic switch that promotes Candida albicans commensalism. Nat Genet. 2013;45:1088–1091.
- Fox EP, Cowley ES, Nobile CJ, Hartooni N, Newman DK, Johnson AD. Anaerobic bacteria grow within Candida albicans biofilms and induce biofilm formation in suspension cultures. Curr Biol. 2014;24:2411–2416.
- Hill JH, Round JL. SnapShot: microbiota effects on host physiology. Cell. 2021;184:2796.
- Xu XL, Lee RT, Fang HM, Wang YM, Li R, Zou H, Zhu Y, Wang Y. Bacterial peptidoglycan triggers Candida albicans hyphal growth by directly activating the adenylyl cyclase Cyr1p. Cell Host Microbe. 2008;4:28–39.
- Alvarez FJ, Konopka JB. Identification of an N-acetylglucosamine transporter that mediates hyphal induction in Candida albicans. Mol Biol Cell. 2007;18:965–975.
- Naseem S, Min K, Spitzer D, Gardin J, Konopka JB. Regulation of Hyphal Growth and N-Acetylglucosamine Catabolism by Two Transcription Factors in Candida albicans. Genetics. 2017;206:299–314.
- Tan CT, Xu X, Qiao Y, Wang Y. A peptidoglycan storm caused by beta-lactam antibiotic’s action on host microbiota drives Candida albicans infection. Nat Commun. 2021;12:2560.
- Brown AO, Graham CE, Cruz MR, Singh KV, Murray BE, Lorenz MC, Garsin DA. Antifungal Activity of the Enterococcus faecalis Peptide EntV Requires Protease Cleavage and Disulfide Bond Formation. mBio. 2019;10:e01334–19.
- Graham CE, Cruz MR, Garsin DA, Lorenz MC. Enterococcus faecalis bacteriocin EntV inhibits hyphal morphogenesis, biofilm formation, and virulence of Candida albicans. Proc Natl Acad Sci USA. 2017;114:4507–4512.
- Cruz MR, Graham CE, Gagliano BC, Lorenz MC, Garsin DA. Enterococcus faecalis inhibits hyphal morphogenesis and virulence of Candida albicans. Infect Immun. 2013;81:189–200.
- Garcia C, Tebbji F, Daigneault M, Liu NN, Kohler JR, Allen-Vercoe E, Sellam A. The Human Gut Microbial Metabolome Modulates Fungal Growth via the TOR Signaling Pathway. mSphere. 2017;2:e00555–17.
- Guinan J, Wang S, Hazbun TR, Yadav H, Thangamani S. Antibiotic-induced decreases in the levels of microbial-derived short-chain fatty acids correlate with increased gastrointestinal colonization of Candida albicans. Sci Rep. 2019;9:8872.
- Hogan DA, Kolter R. Pseudomonas-Candida interactions: an ecological role for virulence factors. Science. 2002;296:2229–2232.
- Morales DK, Grahl N, Okegbe C, Dietrich LE, Jacobs NJ, Hogan DA. Control of Candida albicans metabolism and biofilm formation by Pseudomonas aeruginosa phenazines. mBio. 2013;4:e00526–00512.
- Valentine M, Benade E, Mouton M, Khan W, Botha A. Binary interactions between the yeast Candida albicans and two gut-associated Bacteroides species. Microb Pathog. 2019;135:103619.
- Cuskin F, Lowe EC, Temple MJ, Zhu Y, Cameron EA, Pudlo NA, Porter NT, Urs K, Thompson AJ, Cartmell A, et al. Human gut Bacteroidetes can utilize yeast mannan through a selfish mechanism. Nature. 2015;517:165–169.
- Khosravi A, Mazmanian SK. Disruption of the gut microbiome as a risk factor for microbial infections. Curr Opin Microbiol. 2013;16:221–227.
- Markey L, Shaban L, Green ER, Lemon KP, Mecsas J, Kumamoto CA. Pre-colonization with the commensal fungus Candida albicans reduces murine susceptibility to Clostridium difficile infection. Gut Microbes. 2018;9:497–509.
- Fan D, Coughlin LA, Neubauer MM, Kim J, Kim MS, Zhan X, Simms-Waldrip TR, Xie Y, Hooper LV, Koh AY. Activation of HIF-1alpha and LL-37 by commensal bacteria inhibits Candida albicans colonization. Nat Med. 2015;21:808–814.
- Bhaskaran N, Quigley C, Paw C, Butala S, Schneider E, Pandiyan P. Role of Short Chain Fatty Acids in Controlling Tregs and Immunopathology During Mucosal Infection. Front Microbiol. 2018;9:1995.
- Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol. 2016;14:20–32.
- Johansson ME, Larsson JM, Hansson GC. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions. Proc Natl Acad Sci USA. 2011;108:4659–4665.
- Albenberg L, Esipova TV, Judge CP, Bittinger K, Chen J, Laughlin A, Grunberg S, Baldassano RN, Lewis JD, Li H, et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology. 2014;147:1055–1063.
- Li H, Limenitakis JP, Fuhrer T, Geuking MB, Lawson MA, Wyss M, Brugiroux S, Keller I, Macpherson JA, Rupp S, et al. The outer mucus layer hosts a distinct intestinal microbial niche. Nat Commun. 2015;6:8292.
- Yasuda K, Oh K, Ren B, Tickle TL, Franzosa EA, Wachtman LM, Miller AD, Westmoreland SV, Mansfield KG, Vallender EJ, et al. Biogeography of the intestinal mucosal and lumenal microbiome in the rhesus macaque. Cell Host Microbe. 2015;17:385–391.
- Bohm L, Torsin S, Tint SH, Eckstein MT, Ludwig T, Perez JC. The yeast form of the fungus Candida albicans promotes persistence in the gut of gnotobiotic mice. PLoS Pathog. 2017;13:e1006699.
- Witchley JN, Penumetcha P, Abon NV, Woolford CA, Mitchell AP, Noble SM. Candida albicans Morphogenesis Programs Control the Balance between Gut Commensalism and Invasive Infection. Cell Host Microbe. 2019;25:432–443.
- Eckstein MT, Moreno-Velasquez SD, Perez JC. Gut Bacteria Shape Intestinal Microhabitats Occupied by the Fungus Candida albicans. Curr Biol. 2020;30:4799–4807.
- Donaldson GP, Chou WC, Manson AL, Rogov P, Abeel T, Bochicchio J, Ciulla D, Melnikov A, Ernst PB, Chu H, et al. Spatially distinct physiology of Bacteroides fragilis within the proximal colon of gnotobiotic mice. Nat Microbiol. 2020;5:746–756.
- Pierce JV, Kumamoto CA. Variation in Candida albicans EFG1 expression enables host-dependent changes in colonizing fungal populations. mBio. 2012;3:e00117–00112.
- Pereira FC, Berry D. Microbial nutrient niches in the gut. Environ Microbiol. 2017;19:1366–1378.
- Sonnenburg JL, Xu J, Leip DD, Chen CH, Westover BP, Weatherford J, Buhler JD, Gordon JI. Glycan foraging in vivo by an intestine-adapted bacterial symbiont. Science. 2005;307:1955–1959.
- Wu M, McNulty NP, Rodionov DA, Khoroshkin MS, Griffin NW, Cheng J, Latreille P, Kerstetter RA, Terrapon N, Henrissat B, et al. Genetic determinants of in vivo fitness and diet responsiveness in multiple human gut Bacteroides. Science. 2015;350:aac5992.
- Rakoff-Nahoum S, Coyne MJ, Comstock LE. An ecological network of polysaccharide utilization among human intestinal symbionts. Curr Biol. 2014;24:40–49.
- Mirhakkak MH, Schauble S, Klassert TE, Brunke S, Brandt P, Loos D, Uribe RV, Senne de Oliveira Lino F, Ni Y, Vylkova S, et al. Metabolic modeling predicts specific gut bacteria as key determinants for Candida albicans colonization levels. ISME J. 2021;15:1257–1270.
- Kumamoto CA, Gresnigt MS, Hube B. The gut, the bad and the harmless: candida albicans as a commensal and opportunistic pathogen in the intestine. Curr Opin Microbiol. 2020;56:7–15.
- Lorenz MC. Carbon catabolite control in Candida albicans: new wrinkles in metabolism. mBio. 2013;4:e00034–00013.
- Rosenbach A, Dignard D, Pierce JV, Whiteway M, Kumamoto CA. Adaptations of Candida albicans for growth in the mammalian intestinal tract. Eukaryot Cell. 2010;9:1075–1086.
- Witchley JN, Basso P, Brimacombe CA, Abon NV, Noble SM. Recording of DNA-binding events reveals the importance of a repurposed Candida albicans regulatory network for gut commensalism. Cell Host Microbe. 2021;29:1002–1013.
- Sandai D, Yin Z, Selway L, Stead D, Walker J, Leach MD, Bohovych I, Ene IV, Kastora S, Budge S, et al. The evolutionary rewiring of ubiquitination targets has reprogrammed the regulation of carbon assimilation in the pathogenic yeast Candida albicans. mBio. 2012;3:e00495–12.
- Gutierrez D, Weinstock A, Antharam VC, Gu H, Jasbi P, Shi X, Dirks B, Krajmalnik-Brown R, Maldonado J, Guinan J, et al. Antibiotic-induced gut metabolome and microbiome alterations increase the susceptibility to Candida albicans colonization in the gastrointestinal tract. FEMS Microbiol Ecol. 2020;96:fiz187.
- Romo JA, Markey L, Kumamoto CA. Lipid Species in the GI Tract are Increased by the Commensal Fungus Candida albicans and Decrease the Virulence of Clostridioides difficile. J Fungi. 2020;6:100.
- Kamada N, Chen GY, Inohara N, Nunez G. Control of pathogens and pathobionts by the gut microbiota. Nat Immunol. 2013;14:685–690.
- Hsiao A, Ahmed AM, Subramanian S, Griffin NW, Drewry LL, Petri WA Jr., Haque R, Ahmed T, Gordon JI. Members of the human gut microbiota involved in recovery from Vibrio cholerae infection. Nature. 2014;515:423–426.
- Buffie CG, Bucci V, Stein RR, McKenney PT, Ling L, Gobourne A, No D, Liu H, Kinnebrew M, Viale A, et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015;517:205–208.