
ABSTRACT
Phages are the most abundant and diverse biological entities on Earth and exert specific effects on bacterial hosts. The coexistence of phages and bacteria in the intestinal tract is dynamic and interdependent. Phages are involved in maintaining the stability and composition of the bacterial community, and an imbalance in phages and bacteria in the intestinal tract can cause diseases. This review elucidates interactions between phages and bacteria in the human intestinal tract and their roles in the pathogenesis and treatment of diseases. Understanding the relationship among phages, bacteria and host diseases is conducive to promoting the application of phages in the treatment of human diseases.
List of abbreviations
EMBL-EBI The European Bioinformatics Institute; E. coli Escherichia coli; E. faecalis Enterobacter faecalis; B. fragilis Bacteroides fragilis; B. vulgatus Bacteroides vulgatus; SaPIs Staphylococcus aureus pathogenicity islands; ARGs Antibiotic resistance genes; STEC Shiga toxigenic E. coli; Stx Shiga toxin; BLAST Basic Local Alignment Search Tool; TSST-1 Toxic shock toxin 1; RBPs Receptor-binding proteins; LPS lipopolysaccharide; OMVs Outer membrane vesicles; PT Phosphorothioate; BREX Bacteriophage exclusion; OCR Overcome classical restriction; Pgl Phage growth limitation; DISARM Defense island system associated with restrictionmodification; R-M system Restriction-modification system; BREX system Bacteriophage exclusion system; CRISPR Clustered regularly interspaced short palindromic repeats; Cas CRISPR-associated; PAMs Prospacer adjacent motifs; crRNA CRISPR RNA; SIE; OMPs; Superinfection exclusion; Outer membrane proteins; Abi Abortive infection; TA Toxin-antitoxin; TLR Toll-like receptor; APCs Antigen-presenting cells; DSS Dextran sulfate sodium; IELs Intraepithelial lymphocytes; FMT Fecal microbiota transfer; IFN-γ Interferon-gamma; IBD Inflammatory bowel disease; AgNPs Silver nanoparticles; MDSC Myeloid-derived suppressor cell; CRC Colorectal cancer; VLPs Virus-like particles; TMP Tape measure protein; PSMB4 Proteasome subunit beta type-4; ALD Alcohol-related liver disease; GVHD Graft-versus-host disease; ROS Reactive oxygen species; RA Rheumatoid arthritis; CCP Cyclic citrullinated protein; AMGs Accessory metabolic genes; T1DM Type 1 diabetes mellitus; T2DM Type 2 diabetes mellitus; SCFAs Short-chain fatty acids; GLP-1 Glucagon-like peptide-1; A. baumannii Acinetobacter baumannii; CpG Deoxycytidylinate-phosphodeoxyguanosine; PEG Polyethylene glycol; MetS Metabolic syndrome; OprM Outer membrane porin M.
1. Introduction
The human body is a complex symbiotic organism composed of its own cells and a large number of symbiotic microorganisms. Human microecology includes intestinal microecology, skin microecology, respiratory tract microecology, etc. However, intestinal microecology is the most important in the human microecosystem. Intestinal microecology imbalance may induce systemic diseases.Citation1 Eckburg PB et al. detected 13,355 prokaryotic ribosomal RNA gene sequences in multiple parts of the colonic mucosa and feces of healthy individuals and found that there were at least 1,000 known bacteria in the intestinal tract of each individual. Ninety-eight percent of intestinal bacteria can be classified as Bacteroidetes, Firmicutes, Proteobacteria and actinomycetes. Most of the remaining bacteria have yet to be identified, corresponding to uncultured species and unrecognized microorganisms.Citation2 In addition, there are thousands of viruses in the intestinal tract. By analyzing more than 28,000 intestinal microbiome samples and 2,898 reference genomes of cultured gut bacteria collected in various parts of the world, researchers from the Wellcome Sanger Institute and the European Bioinformatics Institute (EMBL-EBI) have identified more than 140,000 nonredundant gut phage genomes in the human intestinal tract.Citation3 Liang G et al. and Carding SR et al. suggested that phage‒bacterial ratios were basically maintained at 1:1 in the human gut.Citation4,Citation5
Phages are the most common viruses in the human intestine (up to 108 virus-like particles (VLPs) per milliliter in fecal filtrate).Citation6 A special attribute of phages is that they are hosted exclusively in bacteria. Phages were first discovered in Staphylococcus and Shigella in 1915, and the best-known among them today is phage T4, with Escherichia coli serving as its host.Citation7 Phages share similar characteristics with other types of viruses in the fact that they have small sizes (taking E. coli T4 phage as an example, its head size is approximately 95 × 65 nm, its tail is 95 ~ 125 nm long and its tubular structure has a 13 ~ 20 nm diameter), no complete cellular structures and a single nucleic acid segment. Based on their life cycle, phages can be classified into lysogenic phagesCitation8 and lytic phages.Citation9 Lysogenic phages, also known as temperate phages, integrate their viral genetic material into the genome of the host bacteria, where their DNA or RNA is replicated by bacterial chromosomes through normal cell division without cell lysis/cleavage.Citation10 In contrast, lytic phages, whose replication cycle is relatively complex, result in host bacteria lysing following four processes: adsorption, osmosis, synthesis, and release.Citation11 Based on the frequency of their presence in the human intestine, phages can be further classified into low-compatibility phages, conventional phages, and core phages, which are present in 2–19%, 20%-50%, and more than 50% of the human population, respectively.Citation11 The most widely distributed intestinal phage found in recent studies is a type that can infect Bacteroidetes, called “crAssphage”.Citation12
It is typical that phages interact specifically with a single strain of bacteria. Phage–bacterial interaction networks are nested and modular. These interactions are continuously evolving, although their evolution may be influenced by localization in organs and tissues and the complexity of the interaction network.Citation13 Phage activity affects the number and behavior of host bacteria and mediates gene transfer between bacteria during host inflammation. Phages are related to microecological balance and imbalance, and they can affect human health via predation of the bacterial ecological landscape or via more indirect routes, such as influencing metabolism and the immune system. Regulating the relationship between phages and bacteria can maintain the health of the body and even reverse diseases. In the 1930s, phages were used to fight infection in the United States, and later large-scale successful clinical trials were conducted on their use.Citation14 Subsequently, phages have been widely used for the prevention and treatment of bacillary dysentery and staphylococcal infection in South America.Citation15,Citation16 In recent years, phages have been widely used for ultrasensitive biomarker detection, enhanced biological imaging for disease diagnosis, targeted drugs and gene delivery, effective vaccination, replacement of antibiotics for sterilization, and more.
This paper discusses in detail the mode of phage action on bacteria, the mode of bacterial rejection of phages, and the coevolution of phages and bacteria in the human intestinal tract. The relationship between phages and host diseases and recent applications in the medical field are further discussed. Investigating the potential of phage therapy may provide a reference for phages as antimicrobial agents.
2. The relationship between phages and bacteria in the intestine
2.1 Effects of bacteriophage invasion on the intestinal flora
(I) phage-specific lysis-susceptible bacteria
The Escherichia virus PDX is a member of the strictly lytic Myoviridae family. It was reported that Myoviridae phage PDX killed a disease-associated enteroaggregative E. coli (EAEC) isolated from a child from rural Tennessee and an EAEC isolated from a child from Columbia in a dose-dependent manner. Cepko LCS et al. further found that EAEC reduced the β-diversity of the human microbiota, while Myoviridae phage PDX could kill EAEC without causing dysregulation of the human microbiome.Citation17 Lytic phages were injected into conventional mice colonized with a group of identified human symbiotic bacteria. Longitudinal tracking of each microbial response using high-throughput sequencing and quantitative PCR showed that phages T4, F1, B40-8, and VD13 lysed only their susceptible bacteria E. coli, Clostridium sporogenes, Bacteroides fragilis and Enterococcus faecalis, respectively. These phages showed no significant effect on other symbiotic bacteria.Citation18 An in vitro small intestine model was used to analyze the effects of a DSM 1058 phage preparation on preselected target E. coli strains and nontarget bacterial populations. It was found that the phage preparation of E. coli DSM 1058 affected only the population number of E. coli. However, other “symbiotic” bacterial species included in the intestinal model, such as Streptococcus salivarius, Streptococcus lutetiensis and E. faecalis, were not affected.Citation19
(II) phages affect the intestinal flora through horizontal gene transfer
Phages can significantly shape the ecosystem structure by strain-specific predation and mediate horizontal gene transfer by lysing host bacteria.Citation20,Citation21 Phages that can package the DNA of a bacterial host and transfer it to a new host are often called transduction phages.Citation22 Staphylococcus aureus’s highly mobile toxin-carrying pathogenic islands (SaPIs) are particularly suited for packaging and transfer by specific staphylococcal phages. SaPI-encoded toxic shock toxin 1 (TSST-1) and other superantigens are inserted into specific chromosomal sites, where they are excised and replicated by temperate phages. After replication, SaPI DNA is packaged into special small infectious particles that produce specific transfer.Citation23,Citation24 By monitoring the transduction of a sodCIII::neo cassette (a gene sequence) in the Fels-1 prophage from LT2 to a recipient Salmonella strain, Bearson BL et al. confirmed that carbadox induced phage-mediated gene transfer.Citation25 In multidrug-resistant strains DT104 and DT120, carbadox induced generalized transduction of phages, resulting in transfer of chromosome and plasmid DNA containing antibiotic resistance genes (ARGs).Citation25,Citation26 Modi et al. established mouse models of ciprofloxacin and ampicillin resistance and detected phage in feces. They found that phages in feces from ciprofloxacin-resistant mice carried genes encoding quinolone efflux pumps (e.g., NorM, mexD, and mexF), while phages in ampicillin-resistant mice carried genes encoding sensors and response regulators for cell wall synthesis inhibitors (e.g., VanRS) (3-fold increase in reads). The results indicated that phage could regulate the drug resistance of antibiotic-treated bacteria by encoding antibiotic resistance genes.Citation27 By reanalyzing data from Modi et al.’s paper, Enault et al. found that there was also a two- to threefold increase in the bacterial-only clusters of orthologous groups of proteins (COGs). All kinds of bacterial genes were detected more frequently in the microbial metagenome of mice treated with ciprofloxacin and ampicillin, and there was no special selection for ARGs.Citation28 This might be due to protophage induction by antibiotic treatment, with some subpopulations of protophage performing generalized transduction. Antibiotic treatment expanded the interaction between phages and bacterial species, leading to more tightly connected gene exchange networks between phages and bacteria.Citation27,Citation28
(III) phages encode virulence factors of the bacterial population
Phages can spread virulence factors between strains, including toxin-coding genes that cause many diseases, such as diphtheria, cholera, dysentery, and scarlet fever.Citation29 Vibrio cholerae, the pathogen of cholera, requires two coordinating regulators to achieve full virulence: cholera toxin and toxin-coregulated pilus. The structural genes of cholera toxin are encoded by the filamentous phage CTXφ, and the CTXφ genome acts as a plasmid for chromosome integration or replication. The El Tor mutation of the phage CTXφ destroys XerC and XerD, two bacterial-encoded tyrosine recombinases. These two enzymes usually play a role in the decomposition of chromosomal dimers. CTXφ phages integrate at the decomposition site DIF1 of the larger dimer of the two chromosomes of V. cholerae, leading to the genetic diversity of cholera epidemic strains and further affecting the release of cholera toxin by V. cholerae.Citation30,Citation31 The toxin in Shiga toxigenic E. coli (STEC) is encoded by resident temperate lambdoid bacteriophages. Temperate lambdoid bacteriophages might contain toxin structural genes or regulators of toxin structural genes transduced by host bacteria. The Shiga toxin (Stx) gene is expressed when the phage is induced to leave its dormant state and begin replication. Extensive phage replication results in the release of large amounts of Stx from E. coli.Citation32,Citation33 The Stx gene is located downstream of the phage PR promoter, and the transcription of the promoter and the expression of Stx are controlled by the Q antitermination protein. Q antitermination protein is expressed only during phage lysis-mediated growth, so phages carrying the Q21 subtype produce a lower amount of Stx.Citation32
(IV) phages involved in the regulation of bacterial metabolism
Specific predation by phages targeting bacterial species may eliminate the production of relevant bacterial metabolites. The relative abundance of different phages and bacterial metabolites in the intestinal tract of severely depressed patients and healthy persons has been monitored, and a symbiotic network has been thus constructed, showing that the abundance of Klebsiella phage (vB KpnP SU552A) was positively correlated with Bacteroidetes abundance and negatively correlated with proline, cysteine and tryptophan levels, affecting amino acid metabolism.Citation34 Tryptamine is a neurotransmitter produced by the decarboxylation of tryptophan in a few symbiotic intestinal bacteria. Tryptophan decarboxylase-encoding genes have been identified in species such as Ruminococcus gnavus and C. sporogenes.Citation35 Hsu BB et al. first used the Basic Local Alignment Search Tool (BLAST) to search tryptophan decarboxylase amino acid sequences from C. sporogenes (clospo_02083) against the other members of their consortium. The results showed that the amino acid sequence of the tryptophan decarboxylase from C. sporogenes (clospo_02083) had poor protein homology (top hit: 31% identity) with the other members of the consortium. The amino acid sequence of the tryptophan decarboxylase from C. sporogenes maintained its unique association with C. sporogenes. Then, the researchers collected fecal samples from germ-free mice colonized by the defined bacterial consortium and treated them with T4 and F1 phages. During treatment with the T4 and F1 phages, the researchers detected a 10-, 17-, and 2-fold reduction in tryptamine levels (at 0.3, 2, and 13 days, respectively).Citation18 The Lactobacillus species E. faecalis specifically produces the neurotransmitter tyramine through tyrosine decarboxylation.Citation36 No association of tyrosine decarboxylase with other members of the flora has been found, nor has any protein with significant homology to E. faecalis. E. faecalis has been treated with phage, and the tyrosine content was found to be reduced by 4 times at 0.3 days, 2.7 times at 2 days, and 4 times at 13 days.Citation18 The E. faecalis strain from Atp4aSl/Sl mouse feces was isolated and used to isolate E. faecalis phages. The E. faecalis phages Efmus1, Efmus2, Efmus3 and Efmus4 were isolated from untreated raw sewage water. Three or four different phages targeting cytolytic E. faecalis (1010 PFUs) were administered to mice via gavage. It was found that the phages could significantly reduce the number of Enterococcus and levels of lysocytin.Citation37 Phages may change the gene composition of their target hosts through the lysogenic pathway, affecting the expression of metabolite-related genes and later resulting in an increase or decrease in the levels of metabolites.
(V) phages have a cascade of interactions with other bacteria
After reducing the number of their target bacteria, phages will also affect nontarget bacterial species in the symbiotic bacterial community of the intestinal tract through a cascade effect with the result that the overall number of intestinal bacteria is basically stable. For example, E. coli promotes the growth of B. fragilis and inhibits the growth of Bacteroides vulgatus through an interaction network. E. coli depletion by phage T4 resulted in growth inhibition of B. fragilis and enhanced growth of B. vulgatus.Citation18 In addition, after continuous application of phages to mice, the degree of intestinal microbiome dissimilarity between each mouse was gradually reduced. This finding indirectly suggests that phage predation may contribute to the stability of bacterial communities.
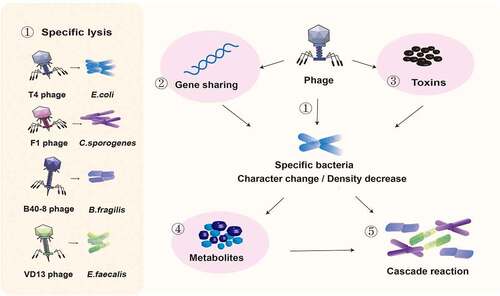
The effects of bacteriophage inoculation on the intestinal flora are shown in .
Figure 1. Effects of bacteriophage inoculation on the intestinal flora. ① Phages T4, F1, B40-8, and VD13 lysed only their susceptible bacteria E. coli, C. sporogenes, B. fragilis and E. faecalis. Phages directly affect susceptible bacteria through ① specific lysis, ② horizontal gene transfer and ③ encoding virulence genes, resulting in changes in intestinal bacteria and a decrease in their number. ④ The type of metabolites secreted may change due to the change in bacterial characteristics, and the total amount of metabolites secreted may decrease due to the decrease in bacterial number. ⑤ Changes in the abundance of bacteria and their metabolites can affect the intestinal environment and the growth of surrounding bacteria.
2.2 Antagonistic mechanisms of intestinal bacteria against phage invasion
Humans are an important part of the Earth’s ecological environment. The intestine is a human organ directly connected with the external environment. Microorganisms that live in the environment also live in human intestines and perform similar functions. Bacteria and phages coexist in a complex and structured interaction network in the intestine, as in the environment.Citation38,Citation39
(I) bacteria prevent phage adsorption by changing or hiding receptors
Phages infect their target bacteria with receptor-binding proteins (RBPs) on the surface of the bacteria. RBPs belong to different biochemical families, mainly represented by surface proteins, polysaccharides and lipopolysaccharides (LPSs). Bacteria prevent phages from binding to receptors by regulating receptor expression, mutating receptors and hiding receptors. For example, V. cholerae reduces O1 antigen expression by regulating the expression of manA and wbeL, two variable genes required for O1 antigen biosynthesis, which helps it avoid phage adsorption.Citation40 The E. coli F strain can produce the outer membrane protein (OMP) TraT and prevents phage adsorption by masking or modifying the OmpA conformation.Citation41 The capsule of the Staphylococcus simulans strain inhibits phage U16 from binding to its receptor, thus inhibiting phage U16 adsorption. Phages bind to Pseudomonas by polysaccharides.Citation42 To prevent infection, Pseudomonas selects for mutations at many common sites associated with mucoid transformation, including mucA and algU, and inhibiting mucoidy.Citation43 However, phages are also not static and can change their structure to bind to new receptors. The RBP of λ phage is encoded by the J gene and can bind to the host surface receptor LamB. When the expression of the LamB gene is suppressed, phages complete subsequent infection by changing the terminal structure of protein J and binding to the new receptor protein OmpF.Citation44 Small modifications can also disguise receptors from phages. The E. coli K1 capsule can block phage T7 infection,Citation45 and Pseudomonas aeruginosa O antigen modification and type IV pilus glycosylation can block phage infection.Citation46,Citation47 However, when E. coli produces a capsule to mask its LPS receptor, phage H4489A often encodes an extracellular hyaluronic acid lyase to degrade the capsule, thereby aiding adsorption.Citation48 In addition, bacteria can also prevent phage adsorption by producing an extracellular matrix and through competitive inhibition. E. coli and V. cholerae reduce phage infection by providing phage-sensitive receptors on outer membrane vesicles (OMVs).Citation49,Citation50 E. coli FhuA is an iron transporter and an entry port for T5 phage. The antimicrobial molecule Microcin J25 uses FhuA as its receptor and competes with phage T5 for binding sites,Citation51 resulting in a reduced chance of infection success.
(II) bacteria destroy phage DNA through restrictive modification, inhibiting the integration and replication of phage genes
Restrictive modification in bacteria means that bacteria mark their genetic material by methylation at specific sites in the genome, and thus, unlabeled DNA is recognized, cleaved and degraded by a host of endonucleases. Type I, II and III restriction-modification (R-M) systems have methyltransferases and restriction endonucleases, which can protect the host by cutting DNA that is not recognized by its distinct code recognition.Citation52 The defense island system associated with restriction-modification (DISARM) methylase, a widespread bacterial defense system, modifies the host CCWGG motif (W = A or T) as a self-DNA marker.Citation53 The SspABCD-SspE phosphorothioate (PT) system in Vibrio cyclitrophicus, E. coli and Streptomyces yokosukanensis constitutes a defensive barrier against a diverse array of phages. SspABCD provides single-chain and high-frequency PTs. SspB in SspABCD binds to SspE as a nickel enzyme. SspE senses sequence-specific PTs by virtue of its PT-stimulated NTPase activity to exert its antiphage activity, and SspE inhibits phage propagation by introducing nicking damage to impair phage DNA replication.Citation54 BREX (for bacteriophage exclusion) is a superfamily of common defense systems of bacteria, such as Bacillus subtilis. Bacteria differentiate self DNA from nonself DNA by methylating specific asymmetric sites using the BrxX (PglX) methyltransferase.Citation55,Citation56 Phage DNA that is not methylated will be unable to replicate and integrate with bacterial DNA. In the case of type I R–M systems, phages encode proteins that bind to the restriction complex and prevent its binding to unmodified recognition sites in phage DNA. A prototypical protein of this kind is OCR (overcome classical restriction) the product of the lytic bacteriophage T7 gene 0.3. As a DNA mimic, OCR binds many host proteins that interact with nucleic acids (e.g., DNA-dependent RNA polymerase). Goldfarb T et al. found that OCR overcomes E. coli BREX defense by specifically binding BrxX methyltransferase.Citation56
(III) bacteria degrade phage DNA by the CRISPR-cas system
The natural clustered regularly interspaced short palindromic repeats (CRISPR) system is divided into three phases: adaptation, expression and interference. During the adaptation phase, foreign DNA fragments (approximately 30–45 nucleotides, also known as prospacers) from invading plasmids or viruses are incorporated into the CRISPR sequence as new spacers. Selection of native spacers from foreign DNA is based on native prospacer adjacent motifs (PAMs). The new spacer provides memory for specific sequences to defend against the corresponding invading plasmid or virus. At the expression stage, the CRISPR array is transcribed into pre-CRISPR RNA, which is further processed into mature CRISPR RNA (crRNA). Each crRNA contains a conserved repeat sequence and a transcription interval that is complementary to the foreign DNA. A crRNA library can target multiple genetic elements because each crRNA corresponds to an invading sequence. In the interference phase, crRNA acts as a guide for specifically targeting the PAM, and Cas9 cleaves the matched DNA. Phage DNA is inserted into the CRISPR gene cluster to form a new spacer sequence, which is transcribed into crRNA. When the phage invades again, crRNA acts as a recognition marker to guide the Cas protein to the phage DNA and then acts as an endonuclease to degrade the phage DNA. For example, the Serrella III-A CRISPR-Cas complex can block phage infection, and Cas13 from the leptin type VI system can shear phage MS2 RNA in E. coli.Citation57 However, five different “CRISPR-resistant” genes have been found in the genomes of phages infecting P. aeruginosa. Mutations in the CRISPR resistance gene of phages prevent them from infecting bacteria carrying a functional CRISPR/Cas system.Citation58 V. cholerae serogroup O1 phage can encode part of the gene sequence of its own functional CRISPR-Cas system, which can target the functional CRISPR-Cas system at the initial stage of infection and destroy the host antiphage defense system. ICP1_2011_A phage targeted the V. cholerae O1 El Tor strain (harboring phage inducible chromosomal islands-like element 1 (PLE1)) by ICP1. The ICP1 phage CRISPR/Cas system consists of six Cas genes and two CRISPR loci (CR1 and CR2). Genomic organization of PLE1 targeted the CRISPR/Cas system of ICP1-related phage. Eleven ICP1-related phages from stools of cholera patients were isolated, five of which encode a CRISPR/Cas system located between ORF 87 and ORF 88 of the ancestral ICP1 genome14. The GC content of this CRISPR/Cas system is the same (~37%) as that of the rest of the ICP1 genome.Citation59
(IV) bacteria prevent the reinjection of phage DNA through a hyperinfection immune system
Superinfection exclusion (SIE) is a process in which proteins that may be anchored to the membrane or associated with elements on the membrane come into play when host bacteria are infected by one phage and another similar phage is adsorbed on the surface of the host cell, thus preventing secondary infections caused by similar phages. Arguijo-Hernández ES et al. presented genetic and biochemical evidence that the E. coli mEp167 Cor protein is an OMP. Cor interacts with OMPs, including OmpA, OmpC, OmpF and OmpW. Cor excluded fHUa-dependent phages in the lysogenic phage strain of E. coli mEp167.Citation60 The superinfection exclusion A (SieA) system of Salmonella typhimurium carrying lysogenic phage P22 can prevent infection by phage LMG178.Citation61 When a bacterial cell is infected with a T-even phage, the phage-encoded proteins Imm and Sp emerge rapidly. Imm prevents DNA transfer across plasma membranes and partially inhibits the release of DNA from reinfected virions, while Sp inhibits the local degradation of bacterial wall proteins by phage-associated lysozyme.Citation62
(V) bacterial infection through the abi system leads to failure of phage lysis
The abortive infection (Abi) system is a mechanism by which cell death is induced after infection but before phage reproduction has completed, thereby protecting uninfected neighbors in a population. One of the Abi systems is the Rex system. E. coli phage λ of RexAB is a typical Abi system. The Rex system can be activated by the phage’s protein‒DNA complex. When the Rex system is triggered, the RexA protein activates the RexB protein, forming membrane channels that result in ATP leakage, loss of membrane potential, and obstruction of phage excretion.Citation63,Citation64 The Abi system can lead to the death of infected cells and often does not produce mature virus particles. Although this system has been studied for decades, we still do not fully understand its mode of action because of the complexity and diversity of the Abi system. There is also another Abi system called the toxin-antitoxin (TA) mechanism. This mechanism is based on the interaction between a toxin and an antitoxin. Endoribonuclease toxin-N was activated in E. coli after infection with T4 and other bacteriophages. Endoribonuclease toxin-N blocks phage development by cleaving viral mRNA and inhibiting its translation.Citation65 The toxin targets essential cellular processes, causing the bacteria to hibernate or die at the same time. For example, for the lncL plasmid of Klebsiella pneumoniae carrying the PemIK (PemK/PemI) Type II TA System, overexpression of PemK toxin results in bacterial dormancy.Citation66 To bypass the toxin, phages can encode antitoxins.Citation67,Citation68 Antitoxins can compete for the same binding site of toxins to disable their function. Alternatively, antitoxins can be chosen to neutralize the toxicity of toxins. Phages can generate a variety of escape methods by recombining to acquire host genetic material, and recombination can also facilitate phage acquisition of antitoxins.
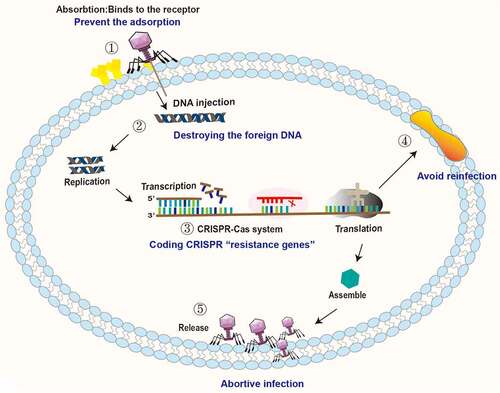
The antagonistic mechanisms of intestinal bacteria against phage invasion are shown in .
Figure 2. Antagonistic mechanisms of intestinal bacteria against phage invasion. ① Bacteria can prevent phage adsorption by changing and hiding receptors. ② Bacteria destroy the DNA of the invading phage through the restrictive modification-methylation pathway and inhibit the replication and integration of phage genes. ③ Bacteria degrade phage DNA by the CRISPR-Cas system. ④ Bacteria prevent reinfection by encoding superinfection immune system proteins. ⑤ Bacteria interfere with phage adsorption, injection, replication, assembly and release through the Abi system, leading to failure of phage lysis.
2.3 Phages and bacteria coexist continuously in the gut
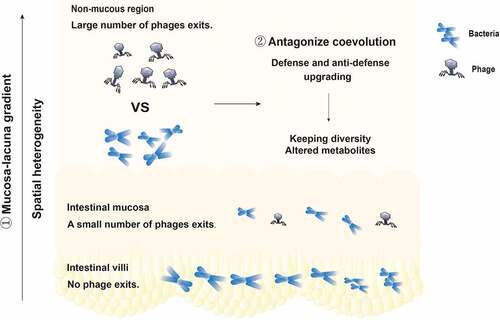
Phages are core members and potential regulators of the intestinal microbiota and play a vital role in maintaining the structure and function of the intestinal microbial community.Citation11 Recent studies have shown that administration of virulent phages can strongly affect intestinal colonization of their target bacteria but still support long-term coexistence.Citation69 The long-term coexistence of phages and gut bacteria is worth considering. Phages drive and maintain the stability and diversity of intestinal microecology by maintaining coevolutionary interactions with their microbial prey. Survival competition,Citation70–73 gene transfer,Citation74 and other factors are involved in the formation and turnover of microbial communities. Specific interactions between phages and bacteria, competition between defense and antidefense, and the mechanisms through which genes are shared to form new mutations and new species combinatorially promote coevolution. The replication and survival of bacteriophages rely on the bacterial host. A high density of bacteria is conducive to the survival of bacteriophages.Citation75 Based on this, the “density dependence” hypothesis states that the more bacteria there are, the more phages there are. Two competing theories about the survival of viruses and bacteria are interesting. One is the kill-the-winner model, and the other is the piggyback-the-winner model.Citation76 When bacterial density increases in an ecosystem, bacteria are called winners. As bacterial density increases in an ecosystem, so does the number of phages that infect those bacteria. It is widely believed that this growing population of phages then kills an increasing number of bacteria, limiting the population size. This is the model called “kill-the-winner”. The “piggyback-the-winner” model refers to the hypothesis that as potential host bacteria become increasingly numerous, some viruses forgo rapid multiplication and instead choose to remain steady in their host. These viruses multiply more slowly, avoid competing with other viruses, and avoid coming into contact with the host’s own immune defenses. These two seemingly contradictory theories are in fact how phages survive. Moreover, the latest research proposes a theory called bacterial spatial distribution, which suggests that the structured environment – the intestinal villi – can function as a spatial refuge for bacteria, allowing them to escape phage predation. This leads to the coexistence of bacteria and phages.Citation77 Bacteriophages have specificity in killing intestinal symbiotic bacteria.Citation75 In the intestinal microecosystem, bacteriophages and symbiotic bacteria have killing specificity, and there are also some bacteriophages and symbiotic bacteria that cannot interact with each other. The cause of the persistent coexistence of phages and bacteria in the intestinal tract is shown in .
Figure 3. The cause of the persistent coexistence of phages and bacteria in the intestinal tract. ① The distribution of bacteriophages and bacteria is spatially heterogeneous. There are no phages in the intestinal villi, few phages in the intestinal mucosa, and a large number of phages in the intestinal lumen. The phage density in the intestinal tract presents a mucosal-lumenal gradient, providing a place for bacteria to avoid phage attack. ② The specific interaction between phages and bacteria, the competition between defense and anti-defense systems, and the way in which genes are shared to form new mutations or new species promote coevolution.
3. Intestinal phages induce diseases by regulating specific bacterial metabolism and initiating the immune response
(I) phages regulate colitis via the immune response
Intestinal microbes are closely related to intestinal diseases such as colitis. Norman JM et al. (2015) observed a decrease in intestinal bacterial population diversity in patients with Crohn’s disease and ulcerative colitis (UC) and found that the occurrence of disease was associated with significant expansion of Caudovirales phages.Citation78 Phage-lysed bacteria release bacterial surface molecules and intercellular contents to activate Toll-like receptor (TLR) signaling of the immune system or influence immune responses by regulating the bacterial community and inducing intestinal inflammation.Citation79 Liu L et al. (2019) found that symbiotic viruses, such as bacteriophages, were recognized by RIG-I receptors expressed in intestinal antigen-presenting cells (APCs). RIG-I receptors promoted the production of IL-15 through the MAVS-IRF-1 signaling pathway and enhanced the activity and function of CD8ααα+TCR-αβ+ and CD8αβ+TCR-αβ+ intraepithelial lymphocytes (IELs). Mice lacking common viruses or MAVS are more likely to develop dextran sulfate sodium (DSS)-induced colitis, and restoring IELs in these mice through IL-15 supplementation reduces susceptibility to DSS.Citation80 Gogokhia L et al. (2019) showed that Lactobacillus phage, E. coli phage, and Bacteroidetes phages stimulated interferon-gamma (IFN-γ) production through the nucleotide sensing receptor TLR9. Internalized phages triggered TLR-9 signaling with phage epitopes presented to CD4 + T cells, leading to the production of IFN-γ and the activation of inflammatory responses in the gut. Phages exacerbate colitis through TLR9 and IFN-γ. Before fecal microbiota transfer (FMT) and 4 weeks after FMT, Gogokhia L et al. performed total nucleic acid sequencing of 20 individuals with active UC. They found that patients who had a clinical response to FMT had a lower relative abundance of Caudovirales bacteriophages than patients who did not respond to therapy. Mucosal IFN-γ levels were positively correlated with phage levels, and phages in patients with active UC induced more IFN-γ than in healthy individuals.Citation69 Adiliaghdam F et al. found that bacteriophages enriched from non-inflammatory bowel disease (IBD) individuals could actively elicit atypical anti-inflammatory innate immune programs. Bacteriophages enriched from IBD individuals divergently provoke proinflammatory macrophage responses. Increasing the ratio of non-IBD-associated bacteriophages in feces could partially restore the ability of macrophages to produce anti-inflammatory cytokines such as IL-22. Harnessing bacteriophages might offer therapeutic potential for IBD.Citation81
(II) Phages affect colorectal cancer by changing the bacterial community structure and regulating the immune microenvironment
Nakatsu G et al. (2018) performed a shotgun metagenomic analysis of stool samples from 74 colorectal cancer (CRC) patients and 92 non-CRC patients in Hong Kong and found that enterovirus population disorder was associated with early and advanced CRC.Citation82 Zheng DW et al. (2019) observed a harmful overpopulation of Fusobacterium nucleatum in mice and patients with CRC, inhibiting the proliferation of beneficial Clostridium butyricum.Citation83 Phages infecting gram-negative bacterial hosts, such as enterotoxigenic B. fragilis, E. coli and Clostridium nucleobacter, are associated with CRC.Citation84–87
Hannigan et al. (2018) analyzed 16S rRNA gene, whole shotgun metagenomic, and purified virus metagenomic sequencing of fecal samples from 30 healthy people, 30 patients with adenoma and 30 patients with CRC. The results showed that the cancer-associated virome consisted primarily of temperate bacteriophages that can indirectly induce cancer development by regulating the bacterial community composition.Citation88 Moreover, phages can spread throughout sterile regions of bodies, including the blood, lymph, organs, and even the brain. Nguyen S et al. reported the rapid and directional transcytosis of diverse bacteriophages across confluent cell layers originating from the gut by incubating phage T4 with T84 (colon epithelial) cells and CaCo2 (colon epithelial) cells. Bacteriophages can access both the vesicular and cytosolic compartments of the eukaryotic cell, and transcytosed phages can traffic through the Golgi apparatus via the endomembrane system.Citation89,Citation90
There are cancer-promoting bacteria and cancer-inhibiting bacteria in the intestinal tract. Similarly, there may be cancer-promoting phages and cancer-inhibiting phages in the intestinal tract. The host adaptive immune response is active against phages and the gut microbiota, with certain microbial antigens able to stimulate memory T cells. Some intestinal microbial antigens can stimulate memory T cells. At the same time, intestinal microorganisms that cross-react with tumor-associated antigens can activate CD4+ and CD8 + T cells that specifically secrete IFN-γ, contributing to the antitumor immune response. Flukiger et al. found that the symbiotic bacterium Enterococcus hirae contained a prephage, which encoded an MHC class I restricted antigen in its tape measure protein (TMP). Mice colonized by E. coli containing a prephage can induce a TMP-specific CD8 + T-cell reaction after cyclophosphamide treatment. Furthermore, the expression of the mimic carcinogenic peptide PSMB4 in phage TMP1 can effectively inhibit the growth of tumors to improve the effect of immunotherapy.Citation91,Citation92 Murgas P et al. (2018) demonstrated that single-chain DNA containing M13 phage had high immunogenicity and could specifically target tumor cell surfaces, trigger inflammation and invasion of activated innate immune cells, overcome tumor-related immunosuppression, and promote antitumor immunity.Citation91 Dong X et al. (2020) achieved the specific elimination of Clostridium symbiosis by electrostatic assembly of silver nanoparticles (AgNPs) on the surface capsid protein (M13@Ag) of M13 phage that specifically binds Clostridium. M13 phage activated the host immune system and inhibited CRC development by activating APCs to alter the tumor immune microenvironment and reduce myeloid-derived suppressor cell (MDSC) amplification at the tumor site.Citation93
(III) phages induce alcoholic liver disease development by increasing bacterial exotoxin release
Alcoholic hepatitis is the most serious form of alcohol-related liver disease (ALD), with a mortality rate of up to 40%. Lu Jiang et al. (2020) extracted VLPs from the feces of 89 patients with alcoholic hepatitis included in a multicenter observational study and conducted metagenomic sequencing to characterize enteroviruses. The authors observed that the diversity of viral groups in fecal samples of patients with alcoholic hepatitis was most significantly higher. The abundances of Escherichia, Enterobacteria and Enterococcus phage were abnormally high, and the number of mammalian viruses, such as Parvoviridae and Herpesviridae, was significantly increased. Duan Y et al. (2019) compared patients with alcoholic hepatitis, subjects with alcohol use disorders, and healthy controls and found a more than 2,700-fold increase in the number of E. faecalis in the gut microflora of the alcoholic hepatitis group.Citation37 Thirty percent of E. faecalis strains have genes that encode an exotoxin called cytolysin. The presence of lysin-positive (lysozyme) E. faecalis was associated with liver disease severity and mortality in patients with alcoholic hepatitis: 89% of lysin-positive patients died within 180 days of admission, compared with 3.8% of those who were lysin-negative. Next, the researchers used a mouse model of ALD to demonstrate that cytolysin directly contributes to disease progression. Cytolysins directly induce liver cell death in vitro and promote liver damage in ethanol-induced liver disease (chronic alcohol diet) mice. Mice colonized with E. faecalis and then fed ethanol had more severe liver damage and hepatic steatosis than the control group. The same trend in ALD severity was observed in germ-free mice receiving fecal transplants from cytolysin-positive patients with alcoholic hepatitis. Researchers have developed a novel treatment that uses phages to specifically target E. faecalis. Four phages capable of lysing cytolytic E. faecalis from Atp4aSl/Sl mouse feces were isolated from sewage. Intragastrically administered with these phages, Atp4aSl/Sl mice had less severe liver damage, steatosis, and inflammation following chronic intragastric administration of ethanol than those administered control phages or vectors. Importantly, according to 16S rRNA sequencing analysis, the fecal abundance of E. faecalis was reduced after the phage intervention. According to qPCR analysis, cytolysin levels were reduced.Citation94–96
(IV) phages may alleviate graft-versus-host disease by reducing inflammation and regulating immunity
Graft-versus-host disease (GVHD) is the most common serious complication after hematopoietic cell transplantation. It is caused by an immune attack by T cells contained in the graft, which recognize the recipient’s foreign tissue, activate lymphocytes, and then develop an immune response to the graft cells. Recent data indicate a correlation between the degree of intestinal microbiome imbalance and outcome and mortality. Enterobacter, Staphylococcaceae, Actinomycetes and Firmicutes are related to the occurrence and severity of GVHD.Citation97–99 Promising preliminary results were obtained for the treatment of GVHD using FMT, in which bacterial-deficient fecal extracts effectively mediated the beneficial effects of FMT; successful FMT is associated with an increase in the number of cauda viruses, suggesting that phages may be a key part of the microbiota responsible for the efficacy of FMT.Citation100,Citation101 Studies have shown that phages can protect intestinal epithelial cells from bacterial invasion and directly inhibit inflammation by interacting with epithelial cells. For example, phages can inhibit reactive oxygen species (ROS) production induced by E. coli in human neutrophils and monocytes and endotoxin-induced ROS production.Citation102,Citation103 Phage tail filament proteins bind to host cell surface-specific receptors and attach to the cell surface. Different phages have different receptors on the host cell surface. LPS is a common receptor for phages such as T3, T4 and T7. LPS is an important bacterial factor inducing the production of inflammatory cytokines, disrupting the internal environment. Phages can interfere with LPS release and induce mononuclear cells to produce IL-10 and IL-1 receptor antagonists to reduce inflammationCitation104–106 and can regulate the functions of dendritic cells, monocytes and granulocytes, inhibit the NF-kappaB pathway stimulated by LPS, and regulate immune function.Citation107 Miernikiewicz P et al. found that the infiltration of leukocytes into the lungs, liver and spleen was markedly increased in mice treated with LPS compared to that in mice treated with PBS. The LPS structure was associated with its ability to induce an inflammatory response. Binding of phage protein gp12 to the hydrophilic core disturbed its function in the formation of TLR4-MD-2-LPS complexes that could lead to immune stimulation.Citation108 Phages reduce the production of inflammatory mediators, which may reduce the development of GVHD and have a protective effect against GVHD in the clinic.
(V) phages and specific bacteria affect rheumatoid arthritis through the immune pathway
Rheumatoid arthritis (RA) is a highly inherited multifactorial autoimmune disease. Studies have shown that RA may be related to changes in the intestinal flora, and intestinal microbes may be involved in immune regulation in RA development as inflammatory mediators.Citation109,Citation110 RA is characterized by the presence of antibodies against cyclic citrullinated protein (CCP) in serum antibody-positive individuals. The heritability of RA is estimated at 40–60%. Familial risk was significantly increased in first-degree relatives (FDRs) of patients diagnosed with RA. Mangalea et al. performed intestinal phage analysis of FDRs of RA patients (with and without anti-CCP antibodies) and found that the intestinal phages of the FDR (high RA risk) group were enriched in those targeting Bacteroidaceae. The number of Streptococcaceae phage and Lachnospiraceae phage in the CCP+FDR group was increased.Citation111 Phages use bacteria such as Lachnospiraceae as their target hosts, and their proliferation is related to an increased abundance of bacteria such as Lachnospiraceae in the CCP+ group.Citation112 Phages enriched in the CCP+FDR group carry the accessory metabolic gene (AMG) PHNP, which encodes a phosphodiesterase and regulates phosphate degradation.Citation108 Mangalea also found that phages enriched in the CCP+FDR group carried clusters of transferases such as mannose-phosphotransferase (algA, xanB, rfbA, wbpW, and PSLB), mannose-heptanose transferase (gmhC, hldE, waaE, and rfaE), GALE (epimerase) and glm transaminase.Citation111 Phages of the CCP+FDR group may influence the formation of bacterial cell wall polysaccharides and biofilms through transferases,Citation113 thus participating in immune evasion. Phages also have immunomodulatory effects through their inherent anti-inflammatory properties and can directly regulate lymphocytes by translocation to multiple tissues.Citation104 The intestinal phage community composition of patients with RA can fluctuate with changes in immune system function and disease, and this community composition has potential as a biomarker for early disease detection.Citation114
(VI) phages affect blood glucose homeostasis by altering intestinal bacterial composition and metabolism
Type 1 diabetes mellitus (T1DM) is a disease in which the autoimmune system attacks Langerhans insulin-producing cells. Type 2 diabetes mellitus (T2DM) is a chronic metabolic disease with high blood sugar in the context of insulin resistance and impaired insulin secretion. The development of both T1DM and T2DM is associated with intestinal microbiome disorders. The abundance of Bacteroidetes increased in T1DM, while the abundance of butyric-producing bacteria decreased.Citation115 In T2DM, the abundance of sulfate-reducing bacteria and Enterobacteriaceae increased, while the abundance of Firmicutes and butyric-producing bacteria decreased.Citation116–118 Changes in the intestinal microbiome composition can affect the functions of the microbiome, such as increased membrane transport of sugars or branched amino acids, increased enzyme activities involved in exogenous or carbohydrate metabolism, and decreased functions involved in cell motility and butyric acid synthesis.Citation116,Citation117 Such changes also affect the metabolic functions of the microbiome, such as short-chain fatty acids (SCFA) production and vitamin metabolism, which are negatively correlated with insulin resistance.Citation119 The binding of SCFAs to GPR43 and GPR41 increases plasma glucagon-like peptide-1 (GLP-1) and peptide YY levels and improves glucose homeostasis.Citation120 Phages may be involved in the occurrence and development of diabetes by affecting bacterial hosts.Citation121 For example, prophage activation significantly induced E. coli biofilms to release amyloid. Amyloid-derived curly fibers may trigger T1DM progression through the TLR2-MyD88-NF-kB signaling pathway.Citation122 Enhancement of intestinal gram-negative bacterial lysis by phages releases LPS, leading to systemic subclinical inflammation and affecting insulin sensitivity.Citation123,Citation124 Zhao G et al. observed changes in enteric phages over time, changes in enteric viruses preceding autoimmunity, and disease-related viral phages associated with specific components of the bacterial microbiome.Citation125 This finding suggests that phages may cause dysglycemia by altering the immune regulation of the gut microbiome. In addition, the biological properties of phages give them the ability to regulate host abundance, thereby influencing bacterial community structure through a cascade of positive and negative interactions between bacterial components of the intestinal microbial community. When the cecal virus population was transferred from lean mice to obese mice, transgraft-induced transgenic changes in the fecal virus group resulted in weight fluctuations in obese mice, and blood glucose parameters returned to normal.Citation126 Transplanting the fecal microbiota of a healthy lean donor to metabolic disease subjects has been shown to improve peripheral insulin sensitivity, with 65% of subjects experiencing a 10% increase in glucose loss during the first 6 weeks after treatment, and bacteriophage groups that could explain these differences were identified and examined.Citation127
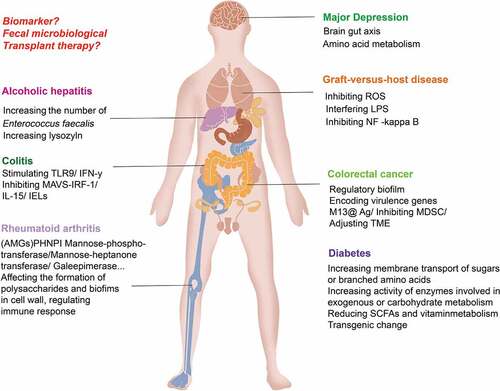
The mechanism of intestinal phage-induced disease is shown in .
Figure 4. Intestinal phages induce the development of diseases by regulating and increasing the release of bacterial exotoxins, regulating bacterial community metabolism, and initiating immune responses. Changes in intestinal phages, bacteria and their metabolites through fecal microbiota transfer may help alleviate diseases. Changes in bacterial metabolites closely related to disease may be able to be used as markers for disease surveillance.
4. Application of phage in the treatment of intestinal diseases
With the emergence of bacteria resistant to many antibiotics, there is increasing interest in phage therapy. Phages are host-specific and can target specific pathogenic bacteria without directly affecting the normal flora of the host. Phages are easy to grow and purify; phages are “live drugs” that increase in number as the target bacterial population spreads and can be administered in small doses; phages are not toxic, and they attack only bacteria, not human cells; and phages are self-limiting, and once the target bacterial population is eliminated, the number of phages will greatly decrease. Therefore, the use of phage therapy is currently a hot topic.
(I) phage cocktails as antimicrobial therapy
The problem of rising antibiotic resistance in recent years has revived researchers’ interest in phage therapy, as phage are the natural enemies of bacteria. The most common bacterial infections of the gastrointestinal tract are diarrhea caused by Clostridioides difficile, dysentery caused by Shigella, gastroenteritis caused by strains of E. coli, cholera caused by V. cholerae, and salmonellosis caused by Salmonella enteritidis. Many studies have assessed the potential of phages for treating gastrointestinal infections caused by E. coli, Enterobacter,Citation128 V. cholerae,Citation129 and C. difficile.Citation130 Phage therapy has successfully provided support for the treatment of intestinal infection in diabetic foot infection,Citation131 lung infection,Citation132 corneal abscessCitation133 and other conditions. For example, the Phagoburn (phase I–II clinical trial) project funded by the European Union under the 7th R&D Framework Programme has conducted a large-scale clinical trial of lysophages for the treatment of infected burn wounds.Citation134 Nestle (Switzerland), in collaboration with The International Centre for Diarrheal Disease Research Dhaka Hospital, Bangladesh, has conducted a phase I/II trial to evaluate the safety and efficacy of oral T4-like phage cocktails in hospitalized children with acute bacterial diarrhea,Citation135 further facilitating the study of phages in enteric infectious diseases. Because of the synergy between individual phages, a phage cocktail may be more effective than a single phage. A 68-year-old man with necrotizing pancreatitis with multidrug-resistant Acinetobacter baumannii infection failed to improve after multiple rounds of antibiotic therapy. The researchers isolated A. baumannii from the patient and screened phages in two different laboratories, mixing the phages targeting the bacteria to produce a phage cocktail, and rapid improvement was achieved with the administration of the phage cocktail by intraperitoneal catheter and intravenous injection.Citation136 Phage injection can be an effective method for controlling antibiotic-resistant bacteria in treating bacterial infections, by using phages that target specific bacteria.
(II) phage vaccines enhance specific immune responses
Phages are composed mainly of nucleic acids and proteins wrapped in a capsid that protects them from nuclease degradation. Phages are excellent carriers of DNA owing to their ability to maintain stability over a range of pH values and resist nuclease degradation. In eukaryotic hosts, phages are inert granular antigens that cannot trigger pathogenesis. Therefore, in recent years, many studies have explored the use of phages as nanodrug platforms to develop vaccines. At present, most phage vaccines are phage display vaccines and phage DNA vaccines.Citation137 Phage display vaccines insert the DNA sequence of the reference foreign protein or polypeptide into the appropriate location of the phage coat protein structural gene so that the foreign gene is expressed along with the coat protein expression. The phage then captures specific molecules such as replicases, pathogen viral factors, or bacterial cell antigens using antigen-binding peptides displayed on the surface of coat proteins. Phage display technology can be used to prepare anti-intestinal pathogenic microorganism vaccines, which can assist in the diagnosis and treatment of intestinal infectious diseases. Phage DNA vaccines are entire phage particles that are used as vectors for genes encoding protective antigenic peptides that are carried to target cells to produce antigens. The proportion of deoxycytidylinate-phosphodeoxyguanosine (CpG) sequences in the phage genome is relatively high, and TLR-9 can recognize CpG sequences and initiate an immune response.Citation138,Citation139 DNA nanodevice vaccines, precisely assembled with two types of molecular adjuvants and an antigenic peptide, have been identified to induce a powerful antigen-specific T-cell response to yield tumor regression, as well as a long-term T-cell response to protect the body from tumor recurrence.Citation140 Phage DNA vaccines targeting intestinal tumors to initiate adaptive immunity while improving the intestinal microenvironment are worthy of further exploration as potential treatments.
(III) phage module exchange or gene editing
Bacteria are central to human health and disease conditions, but existing tools for editing microbial consortia are limited. For example, broad-spectrum antibiotics do not offer precise control over bacterial communities. Synthetic biology is beginning to solve this problem; microbial synthetic biology refers to a customized biological application system that modifies the internal structure of microorganisms to mimic the functions of specific engineering systems.Citation141 Using synthetic biology to edit phages can enlarge their host range, enhance the ability of phages to penetrate biofilms, and improve the targeted use of phages in antibacterial therapy. Ando H et al. directed E. coli phage scaffolds to pathogenic Yersinia and Klebsiella; in contrast, Klebsiella phage scaffolds were directed to E. coli through a module exchange of phage tail components. Synthetic phages can effectively kill target bacteria and selectively remove specific bacteria from multispecies bacterial communities using mixtures based on common viral scaffolds.Citation142 Phages modified by the CRISPR/Cas system can sensitize resistant bacteria by eliminating drug-resistant plasmids. Synthetic biologist Timothy Lu of Massachusetts Institute of Technology and his team used DNA programming CRISPR technology to build engineered phages that specifically infect and kill drug-resistant bacteria. The phage targets bacteria with drug-resistant DNA sequences via fragments of RNA. If the bacterium contains a drug-resistant DNA sequence, the RNA can bind to this sequence and bind the Cas9 enzyme to cleave the bacterial DNA and kill the bacterium.Citation143 Jalasvuori et al. showed that the use of plasmid-dependent phages on E. coli and Salmonella enterica resulted in a significant reduction in the number of resistant bacteria.Citation144 Bacteriophages are potential candidates for replacing antibiotics in the control of infectious bacteria. However, host bacterial resistance to phages is inevitable. Interestingly, gut bacteria that respond to phage resistance recover their sensitivity to certain antibiotics. P. aeruginosa has a receptor-binding site, the outer membrane porin M (OprM), of the multidrug efflux systems MexAB and MexXY. Chan BK et al. found that the evolution of P. aeruginosa resistance to phage attack changed the efflux pump mechanism, causing increased sensitivity to antibiotic drugs from ciprofloxacin and tetracycline.Citation145 Fong K et al. found that sensitivity to tetracycline was increased, while S. enterica developed resistance to Bacteriophage SI1 via mutated genes involved in type VI secretion that contributes to LPS production.Citation146
(IV) phage-targeted delivery of therapeutic drugs for intestinal diseases
Phages can be engineered to form self-assembled nanomaterials with affinity properties. Drugs attached to the surface of the phage can be carried by the phage to a specific location in the body. Drug-carrying phages can then improve a disease environment by specifically lysing bacteria, releasing drugs or binding to antibody receptors on the membranes of specific cancer cells, participating in endocytosis, or treating intracellular degradation. For example, dextran nanoparticles loaded with the cancer chemotherapeutic irinotecan covalently attached to azide-modified phages have been used to treat the overgrowth of F. nucleatum, which was significantly inhibited in mice with in situ-induced CRC or naturally formed CRC after oral or intravenous administration of the modified phages, and CRC was significantly ameliorated under the therapeutic effect of phage-delivered irinotecan. It was also found that oral phage-guided irinotecan nanoparticles did not cause significant changes in blood cell count, immunoglobulin and histamine levels, or liver and kidney function in piglets.Citation83 The emergence of this type of technology for covalent modification of viral particles lays a foundation for the transformation of viral capsids into targeted drug carrier systems.Citation147 Kovacs et al. were able to coat genome-free MS2 capsids with polyethylene glycol (PEG) chains and incorporate 50–70 copies of fluorescent drug mimics into the capsids. Studies have shown that despite extensive modification, the capsid remains assembled, making it an effective delivery vessel for drugs.Citation148 Drugs can also attach to the phage surface without damaging the cell target. For example, drug-carrying liposomes can deliver drugs to cells via phage-liposome complexes.Citation149 Phage-carried drugs naturally existing in the intestinal tract have the characteristics of excellent targeting, strong therapeutic effects and few toxic side effects, holding the potential for serving as ideal drug delivery tools.
The application of phages in the treatment of intestinal diseases is shown in .
Figure 5. Application of phage in the treatment of intestinal diseases. ① Using the nature of phage, a phage cocktail is made to target bacterial infection. ② The structure of phages can be modified by switching phage modules or by applying CRISPR gene editing technology to change the intestinal bacterial host or enhance its antagonism to the intestinal bacterial host. ③ Preparation of phage display vaccines and phage DNA vaccines to enhance specific immune responses. ④Using phage to treat specific intestinal or other bacteria with targeted delivery of disease treatment drugs. For example, glucan nanoparticles are covalently linked to azide-modified phages, polyethylene glycol capsid-modified phages, phage-liposome complexes and other anti-colorectal cancer drugs.
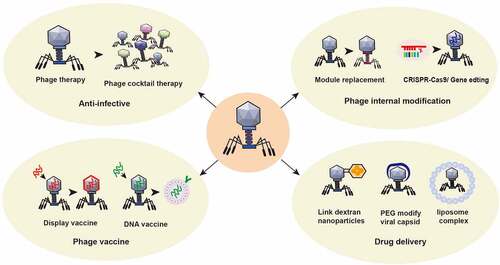
(V) potential limitations of phage therapy
Not all phages are suitable for therapeutic use. Lytic phages are commonly used for clinical treatment. It is required to have specific fracture characteristics and stable fracture effects (independent of temperature and environment) and to ensure safety and efficiency (no toxin protein gene in the genome). Bacteriophage preparations may contain endotoxic proteins of host bacteria, and the bacterial endotoxins released after lysing the host may affect the normal function of the body. Phages that do not lyse bacteria quickly are less effective against them. Phages usually act only on a certain genus or species of bacteria, and some even act only on a limited number of strains of a species. Phage preparations mainly include phage nucleic acids, capsid proteins and so on in the clinic. These preparations are more complex than common clinical pharmaceutical preparations with a single chemical structure. It is difficult to evaluate the activity and purity of drugs, and it is impossible to accurately define the method of administration, dosage form, dose, concentration and administration time of phage preparations. The pharmacokinetics of phage preparations are not clear, and the safety of phage after entering the body cannot be evaluated. Phages are proliferating, evolving, and gene-editing organisms that interact with the body’s immune system.Citation150–154 It is not clear whether the use of phages can also adversely affect the human immune system.
Author contributions
All authors participated in the conception and design of the study. Han Shuwen wrote the manuscript. Ding Kefeng reviewed and sorted the literature. Han Shuwen designed and drew the figures. All authors read and approved the paper.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Kåhrström CT, Pariente N, Weiss U. Intestinal microbiota in health and disease. Nature. 2016;535(7610):47.
- Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science. 2005;308(5728):1635–23.
- Camarillo-Guerrero LF, Almeida A, Rangel-Pineros G, Finn RD, Lawley TD. Massive expansion of human gut bacteriophage diversity. Cell. 2021;184(4):1098–1109.e1099.
- Liang G, Zhao C, Zhang H, Mattei L, Sherrill-Mix S, Bittinger K, Kessler LR, Wu GD, Baldassano RN, DeRusso P, et al. The stepwise assembly of the neonatal virome is modulated by breastfeeding. Nature. 2020;581(7809):470–474.
- Reyes A, Haynes M, Hanson N, Angly FE, Heath AC, Rohwer F, Gordon JI. Viruses in the faecal microbiota of monozygotic twins and their mothers. Nature. 2010;466(7304):334–338.
- Shkoporov AN, Clooney AG, Sutton TDS, Ryan FJ, Daly KM, Nolan JA, McDonnell SA, Khokhlova EV, Draper LA, Forde A, et al. The human gut virome is highly diverse, stable, and individual specific. Cell Host Microbe. 2019;26(4):527–541.e525.
- James TD, Cardozo T, Abell LE, Hsieh ML, Jenkins LM, Jha SS, Hinton DM. Visualizing the phage T4 activated transcription complex of DNA and E. coli RNA polymerase. Nucleic Acids Res. 2016;44(16):7974–7988.
- Erez Z, Steinberger-Levy I, Shamir M, Doron S, Stokar-Avihail A, Peleg Y, Melamed S, Leavitt A, Savidor A, Albeck S, et al. Communication between viruses guides lysis-lysogeny decisions. Nature. 2017;541(7638):488–493.
- Yosef I, Manor M, Kiro R, Qimron U. Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria. Proc Natl Acad Sci U S A. 2015;112(23):7267–7272.
- Al-Anany AM, Fatima R, Hynes AP. Temperate phage-antibiotic synergy eradicates bacteria through depletion of lysogens. Cell Rep. 2021;35(8):109172.
- Manrique P, Bolduc B, Walk ST, van der Oost J, de Vos WM, Young MJ. Healthy human gut phageome. Proc Natl Acad Sci U S A. 2016;113(37):10400–10405.
- Edwards RA, Vega AA, Norman HM, Ohaeri M, Levi K, Dinsdale EA, Cinek O, Aziz RK, McNair K, Barr JJ, et al. Global phylogeography and ancient evolution of the widespread human gut virus crAssphage. Nat Microbiol. 2019;4(10):1727–1736.
- Flores CO, Meyer JR, Valverde S, Farr L, Weitz JS. Statistical structure of host-phage interactions. Proc Natl Acad Sci U S A. 2011;108(28):E288–297.
- Petrovic Fabijan A, Lin RCY, Ho J, Maddocks S, Ben Zakour NL, Iredell JR, Khalid A, Venturini C, Chard R, Morales S. Safety of bacteriophage therapy in severe Staphylococcus aureus infection. Nat Microbiol. 2020;5(3):465–472.
- Doore SM, Schrad JR, Dean WF, Dover JA, Parent KN. Shigella phages isolated during a dysentery outbreak reveal uncommon structures and broad species diversity. J Virol. 2018;92(8)
- Quiles-Puchalt N, Carpena N, Alonso JC, Novick RP, Marina A, Penadés JR. Staphylococcal pathogenicity island DNA packaging system involving cos-site packaging and phage-encoded HNH endonucleases. Proc Natl Acad Sci U S A. 2014;111(16):6016–6021.
- Cepko LCS, Garling EE, Dinsdale MJ, Scott WP, Bandy L, Nice T, Faber-Hammond J, Mellies JL. Myoviridae phage PDX kills enteroaggregative Escherichia coli without human microbiome dysbiosis. J Med Microbiol. 2020;69(2):309–323.
- Hsu BB, Gibson TE, Yeliseyev V, Liu Q, Lyon L, Bry L, Silver PA, Gerber GK. Dynamic modulation of the gut microbiota and metabolome by bacteriophages in a mouse model. Cell Host Microbe. 2019;25(6):803–814.e805.
- Cieplak T, Soffer N, Sulakvelidze A, Nielsen DS. A bacteriophage cocktail targeting Escherichia coli reduces E. coli in simulated gut conditions, while preserving a non-targeted representative commensal normal microbiota. Gut Microbes. 2018;9(5):391–399.
- Tzipilevich E, Habusha M, Ben-Yehuda S. Acquisition of phage sensitivity by bacteria through exchange of phage receptors. Cell. 2017;168(1–2):186–199.e112.
- Andam CP, Williams D, Gogarten JP. Biased gene transfer mimics patterns created through shared ancestry. Proc Natl Acad Sci U S A. 2010;107(23):10679–10684.
- Chen J, Novick RP. Phage-mediated intergeneric transfer of toxin genes. Science. 2009;323(5910):139–141.
- Lindsay JA, Ruzin A, Ross HF, Kurepina N, Novick RP. The gene for toxic shock toxin is carried by a family of mobile pathogenicity islands in Staphylococcus aureus. Mol Microbiol. 1998;29(2):527–543.
- Ruzin A, Lindsay J, Novick RP. Molecular genetics of SaPI1–a mobile pathogenicity island in Staphylococcus aureus. Mol Microbiol. 2001;41(2):365–377.
- Bearson BL, Allen HK, Brunelle BW, Lee IS, Casjens SR, Stanton TB. The agricultural antibiotic carbadox induces phage-mediated gene transfer in Salmonella. Front Microbiol. 2014;5:52.
- Johnson TA, Looft T, Severin AJ, Bayles DO, Nasko DJ, Wommack KE, Howe A, Allen HK. The in-feed antibiotic carbadox induces phage gene transcription in the swine gut microbiome. mBio. 2017;8(4)
- Modi SR, Lee HH, Spina CS, Collins JJ. Antibiotic treatment expands the resistance reservoir and ecological network of the phage metagenome. Nature. 2013;499(7457):219–222.
- Enault F, Briet A, Bouteille L, Roux S, Sullivan MB, Petit MA. Phages rarely encode antibiotic resistance genes: a cautionary tale for virome analyses. Isme j. 2017;11(1):237–247.
- Meinel DM, Margos G, Konrad R, Krebs S, Blum H, Sing A. Next generation sequencing analysis of nine Corynebacterium ulcerans isolates reveals zoonotic transmission and a novel putative diphtheria toxin-encoding pathogenicity island. Genome Med. 2014;6(11):113.
- Das B, Bischerour J, Val ME, Barre FX. Molecular keys of the tropism of integration of the cholera toxin phage. Proc Natl Acad Sci U S A. 2010;107(9):4377–4382.
- Waldor MK, Mekalanos JJ. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science. 1996;272:1910–1914.
- Koudelka GB, Arnold JW, Chakraborty D. Evolution of STEC virulence: insights from the antipredator activities of Shiga toxin producing E. coli. Int J Med Microbiol. 2018;308(7):956–961.
- O’Brien AD, Newland JW, Miller SF, Holmes RK, Smith HW, Formal SB. Shiga-like toxin-converting phages from Escherichia coli strains that cause hemorrhagic colitis or infantile diarrhea. Science. 1984;226(4675):694–696.
- Yang J, Zheng P, Li Y, Wu J, Tan X, Zhou J, Sun Z, Chen X, Zhang G, Zhang H, et al. Landscapes of bacterial and metabolic signatures and their interaction in major depressive disorders. Sci Adv. 2020;6(49)
- Williams BB, Van Benschoten AH, Cimermancic P, Donia MS, Zimmermann M, Taketani M, Ishihara A, Kashyap PC, Fraser JS, Fischbach MA. Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine. Cell Host Microbe. 2014;16(4):495–503.
- Connil N, Le Breton Y, Dousset X, Auffray Y, Rincé A, Prévost H. Identification of the Enterococcus faecalis tyrosine decarboxylase operon involved in tyramine production. Appl Environ Microbiol. 2002;68(7):3537–3544.
- Duan Y, Llorente C, Lang S, Brandl K, Chu H, Jiang L, White RC, Clarke TH, Nguyen K, Torralba M, et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature. 2019;575(7783):505–511.
- De Sordi L, Lourenço M, Debarbieux L. “I will survive”: a tale of bacteriophage-bacteria coevolution in the gut. Gut Microbes. 2019;10(1):92–99.
- De Sordi L, Lourenço M, Debarbieux L. The battle within: interactions of bacteriophages and bacteria in the gastrointestinal tract. Cell Host Microbe. 2019 Feb 13;25(2):210–218.
- Seed KD, Faruque SM, Mekalanos JJ, Calderwood SB, Qadri F, Camilli A. Phase variable O antigen biosynthetic genes control expression of the major protective antigen and bacteriophage receptor in vibrio cholerae O1. PLoS Pathog. 2012;8(9):e1002917.
- Riede I, Eschbach ML. Evidence that TraT interacts with OmpA of Escherichia coli. FEBS Lett. 1986;205(2):241–245.
- Ohshima Y, Schumacher-Perdreau F, Peters G, Pulverer G. The role of capsule as a barrier to bacteriophage adsorption in an encapsulated Staphylococcus simulans strain. Med Microbiol Immunol. 1988;177(4):229–233.
- Scanlan PD, Buckling A. Co-evolution with lytic phage selects for the mucoid phenotype of Pseudomonas fluorescens SBW25. Isme j. 2012;6(6):1148–1158.
- Petrie KL, Palmer ND, Johnson DT, Medina SJ, Yan SJ, Li V, Burmeister AR, Meyer JR. Destabilizing mutations encode nongenetic variation that drives evolutionary innovation. Science. 2018;359(6383):1542–1545.
- Scholl D, Adhya S, Merril C. Escherichia coli K1ʹs capsule is a barrier to bacteriophage T7. Appl Environ Microbiol. 2005;71(8):4872–4874.
- Harvey H, Bondy-Denomy J, Marquis H, Sztanko KM, Davidson AR, Burrows LL. Pseudomonas aeruginosa defends against phages through type IV pilus glycosylation. Nat Microbiol. 2018;3(1):47–52.
- Bondy-Denomy J, Qian J, Westra ER, Buckling A, Guttman DS, Davidson AR, Maxwell KL. Prophages mediate defense against phage infection through diverse mechanisms. Isme j. 2016;10(12):2854–2866.
- Singh SK, Malhotra S, Akhtar MS. Characterization of hyaluronic acid specific hyaluronate lyase (HylP) from Streptococcus pyogenes. Biochimie. 2014;102:203–210.
- Manning AJ, Kuehn MJ. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011;11(1):258.
- Reyes-Robles T, Dillard RS, Cairns LS, Silva-Valenzuela CA, Housman M, Ali A, Wright ER, Camilli A. Vibrio cholerae outer membrane vesicles inhibit bacteriophage infection. J Bacteriol. 2018;200(15)
- Roier S, Zingl FG, Cakar F, Durakovic S, Kohl P, Eichmann TO, Klug L, Gadermaier B, Weinzerl K, Prassl R, et al. A novel mechanism for the biogenesis of outer membrane vesicles in gram-negative bacteria. Nat Commun. 2016;7(1):10515.
- Huang X, Wang J, Li J, Liu Y, Liu X, Li Z, Kurniyati K, Deng Y, Wang G, Ralph JD, et al. Prevalence of phase variable epigenetic invertons among host-associated bacteria. Nucleic Acids Res. 2020;48(20):11468–11485.
- Ofir G, Melamed S, Sberro H, Mukamel Z, Silverman S, Yaakov G, Doron S, Sorek R. DISARM is a widespread bacterial defence system with broad anti-phage activities. Nat Microbiol. 2018;3(1):90–98.
- Xiong X, Wu G, Wei Y, Liu L, Zhang Y, Su R, Jiang X, Li M, Gao H, Tian X, et al. SspABCD-SspE is a phosphorothioation-sensing bacterial defence system with broad anti-phage activities. Nat Microbiol. 2020;5(7):917–928.
- Goldfarb T, Sberro H, Weinstock E, Cohen O, Doron S, Charpak-Amikam Y, Afik S, Ofir G, Sorek R. BREX is a novel phage resistance system widespread in microbial genomes. Embo J. 2015;34(2):169–183.
- Isaev A, Drobiazko A, Sierro N, Gordeeva J, Yosef I, Qimron U, Ivanov NV, Severinov K. Phage T7 DNA mimic protein Ocr is a potent inhibitor of BREX defence. Nucleic Acids Res. 2020;48(10):5397–5406.
- Malone LM, Warring SL, Jackson SA, Warnecke C, Gardner PP, Gumy LF, Fineran PC. A jumbo phage that forms a nucleus-like structure evades CRISPR-Cas DNA targeting but is vulnerable to type III RNA-based immunity. Nat Microbiol. 2020;5(1):48–55.
- Bondy-Denomy J, Pawluk A, Maxwell KL, Davidson AR. Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system. Nature. 2013;493(7432):429–432.
- Seed KD, Lazinski DW, Calderwood SB, Camilli A. A bacteriophage encodes its own CRISPR/Cas adaptive response to evade host innate immunity. Nature. 2013;494(7438):489–491.
- Arguijo-Hernández ES, Hernandez-Sanchez J, Briones-Peña SJ, Oviedo N, Mendoza-Hernández G, Guarneros G, Kameyama L. Cor interacts with outer membrane proteins to exclude FhuA-dependent phages. Arch Virol. 2018;163(11):2959–2969.
- Susskind MM, Botstein D, Wright A. Superinfection exclusion by P22 prophage in lysogens of Salmonella typhimurium. III. Failure of superinfecting phage DNA to enter sieA+ lysogens. Virology. 1974;62(2):350–366.
- Lu MJ, Henning U. Superinfection exclusion by T-even-type coliphages. Trends Microbiol. 1994;2(4):137–139.
- Parma DH, Snyder M, Sobolevski S, Nawroz M, Brody E, Gold L. The Rex system of bacteriophage lambda: tolerance and altruistic cell death. Genes Dev. 1992 Mar;6(3):497–510.
- Alattas H, Wong S, Slavcev RA. Identification of Escherichia coli host genes that influence the bacteriophage lambda (λ) T4rII exclusion (Rex) phenotype. Genetics. 2020 Dec;216(4):1087–1102.
- Guegler CK, Laub MT. Shutoff of host transcription triggers a toxin-antitoxin system to cleave phage RNA and abort infection. Mol Cell. 2021;81(11):2361–2373.e2369.
- Bleriot I, Blasco L, Pacios O, Fernández-García L, Ambroa A, López M, Ortiz-Cartagena C, Cuenca FF, Oteo-Iglesias J, Á P, et al. The role of PemIK (PemK/PemI) type II TA system from Klebsiella pneumoniae clinical strains in lytic phage infection. Sci Rep. 2022;12(1):4488.
- Kurata T, Saha CK, Buttress JA, Mets T, Brodiazhenko T, Turnbull KJ, Awoyomi OF, Oliveira SRA, Jimmy S, Ernits K, et al. A hyperpromiscuous antitoxin protein domain for the neutralization of diverse toxin domains. Proc Natl Acad Sci U S A. 2022;119(6)
- Chattopadhyay G, Bhasin M, Ahmed S, Gosain TP, Ganesan S, Das S, Thakur C, Chandra N, Singh R, Varadarajan R. Functional and biochemical characterization of the MazEF6 toxin-antitoxin system of mycobacterium tuberculosis. J Bacteriol. 2022;204(4):e0005822.
- Gogokhia L, Buhrke K, Bell R, Hoffman B, Brown DG, Hanke-Gogokhia C, Ajami NJ, Wong MC, Ghazaryan A, Valentine JF, et al. Expansion of bacteriophages is linked to aggravated intestinal inflammation and colitis. Cell Host Microbe. 2019;25(2):285–299.e288.
- Williams HT. Phage-induced diversification improves host evolvability. BMC Evol Biol. 2013;13(1):17.
- Meaden S, Biswas A, Arkhipova K, Morales SE, Dutilh BE, Westra ER, Fineran PC. High viral abundance and low diversity are associated with increased CRISPR-Cas prevalence across microbial ecosystems. Curr Biol. 2022;32(1):220–227.e225.
- Andersson AF, Banfield JF. Virus population dynamics and acquired virus resistance in natural microbial communities. Science. 2008;320(5879):1047–1050.
- Liu Y, Dai L, Dong J, Chen C, Zhu J, Rao VB, Tao P. Covalent modifications of the bacteriophage genome confer a degree of resistance to bacterial CRISPR systems. J Virol. 2020;94(23)
- Venkataraman A, Bassis CM, Beck JM, Young VB, Curtis JL, Huffnagle GB, Schmidt TM. Application of a neutral community model to assess structuring of the human lung microbiome. mBio. 2015;6(1)
- de Jonge Pa, Nobrega FL, Brouns SJJ, Dutilh BE, de Jonge PA. Molecular and evolutionary determinants of bacteriophage host range. Trends Microbiol. 2019;27(1):51–63.
- Knowles B, Silveira CB, Bailey BA, Barott K, Cantu VA, Cobián-Güemes AG, Coutinho FH, Dinsdale EA, Felts B, Furby KA, et al. Lytic to temperate switching of viral communities. Nature. 2016;531(7595):466–470.
- Lourenço M, Chaffringeon L, Lamy-Besnier Q, Pédron T, Campagne P, Eberl C, Bérard M, Stecher B, Debarbieux L, De Sordi L. The spatial heterogeneity of the gut limits predation and fosters coexistence of bacteria and bacteriophages. Cell Host Microbe. 2020;28(3):390–401.e395.
- Norman JM, Handley SA, Baldridge MT, Droit L, Liu CY, Keller BC, Kambal A, Monaco CL, Zhao G, Fleshner P, et al. Disease-specific alterations in the enteric virome in inflammatory bowel disease. Cell. 2015;160(3):447–460.
- Gogokhia L, Round JL. Immune-bacteriophage interactions in inflammatory bowel diseases. Curr Opin Virol. 2021;49:30–35.
- Liu L, Gong T, Tao W, Lin B, Li C, Zheng X, Zhu S, Jiang W, Zhou R. Commensal viruses maintain intestinal intraepithelial lymphocytes via noncanonical RIG-I signaling. Nat Immunol. 2019;20(12):1681–1691.
- Adiliaghdam F, Amatullah H, Digumarthi S, Saunders TL, Rahman RU, Wong LP, Sadreyev R, Droit L, Paquette J, Goyette P, et al. Human enteric viruses autonomously shape inflammatory bowel disease phenotype through divergent innate immunomodulation. Sci Immunol. 2022;7(70):eabn6660.
- Nakatsu G, Zhou H, Wkk W, Wong SH, Coker OO, Dai Z, Li X, Szeto CH, Sugimura N, Lam TY, et al. Alterations in enteric virome are associated with colorectal cancer and survival outcomes. Gastroenterology. 2018;155(2):529–541.e525.
- Zheng DW, Dong X, Pan P, Chen KW, Fan JX, Cheng SX, Zhang XZ. Phage-guided modulation of the gut microbiota of mouse models of colorectal cancer augments their responses to chemotherapy. Nat Biomed Eng. 2019;3(9):717–728.
- Boleij A, Hechenbleikner EM, Goodwin AC, Badani R, Stein EM, Lazarev MG, Ellis B, Carroll KC, Albesiano E, Wick EC, et al. The Bacteroides fragilis toxin gene is prevalent in the colon mucosa of colorectal cancer patients. Clin Infect Dis. 2015;60(2):208–215.
- Castellarin M, Warren RL, Freeman JD, Dreolini L, Krzywinski M, Strauss J, Barnes R, Watson P, Allen-Vercoe E, Moore RA, et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012;22(2):299–306.
- Cuevas-Ramos G, Petit CR, Marcq I, Boury M, Oswald E, Nougayrède JP. Escherichia coli induces DNA damage in vivo and triggers genomic instability in mammalian cells. Proc Natl Acad Sci U S A. 2010;107(25):11537–11542.
- Niu YD, McAllister TA, Nash JH, Kropinski AM, Stanford K. Four Escherichia coli O157:H7 phages: a new bacteriophage genus and taxonomic classification of T1-like phages. PLoS One. 2014;9(6):e100426.
- Hannigan GD, Duhaime MB, MTt R, Koumpouras CC, Schloss PD. Diagnostic potential and interactive dynamics of the colorectal cancer virome. mBio. 2018;9(6)
- Nguyen S, Baker K, Padman BS, Patwa R, Dunstan RA, Weston TA, Schlosser K, Bailey B, Lithgow T, Lazarou M, et al. Bacteriophage transcytosis provides a mechanism to cross epithelial cell layers. mBio. 2017;8(6)
- Lehti TA, Pajunen MI, Skog MS, Finne J. Internalization of a polysialic acid-binding Escherichia coli bacteriophage into eukaryotic neuroblastoma cells. Nat Commun. 2017;8(1):1915.
- Murgas P, Bustamante N, Araya N, Cruz-Gómez S, Durán E, Gaete D, Oyarce C, López E, Herrada AA, Ferreira N, et al. A filamentous bacteriophage targeted to carcinoembryonic antigen induces tumor regression in mouse models of colorectal cancer. Cancer Immunol Immunother. 2018;67(2):183–193.
- Fluckiger A, Daillère R, Sassi M, Sixt BS, Liu P, Loos F, Richard C, Rabu C, Alou MT, Goubet AG, et al. Cross-reactivity between tumor MHC class I-restricted antigens and an enterococcal bacteriophage. Science. 2020;369(6506):936–942.
- Dong X, Pan P, Zheng DW, Bao P, Zeng X, Zhang XZ. Bioinorganic hybrid bacteriophage for modulation of intestinal microbiota to remodel tumor-immune microenvironment against colorectal cancer. Sci Adv. 2020;6(20):eaba1590.
- Jiang L, Lang S, Duan Y, Zhang X, Gao B, Chopyk J, Schwanemann LK, Ventura-Cots M, Bataller R, Bosques-Padilla F, et al. Intestinal virome in patients with alcoholic hepatitis. Hepatology. 2020;72(6):2182–2196.
- Frederick E. Bacterial toxin linked to severe alcoholic liver disease. Science. 2019;366(6467):784.
- Ray K. Manipulating the gut microbiota to combat alcoholic hepatitis. Nat Rev Gastroenterol Hepatol. 2020;17(1):3.
- Mancini N, Greco R, Pasciuta R, Barbanti MC, Pini G, Morrow OB, Morelli M, Vago L, Clementi N, Giglio F, et al. Enteric microbiome markers as early predictors of clinical outcome in allogeneic hematopoietic stem cell transplant: results of a prospective study in adult patients. Open Forum Infect Dis. 2017;4(4):ofx215.
- Golob JL, Pergam SA, Srinivasan S, Fiedler TL, Liu C, Garcia K, Mielcarek M, Ko D, Aker S, Marquis S, et al. Stool microbiota at neutrophil recovery is predictive for severe acute graft vs host disease after hematopoietic cell transplantation. Clin Infect Dis. 2017;65(12):1984–1991.
- Taur Y, Jenq RR, Perales MA, Littmann ER, Morjaria S, Ling L, No D, Gobourne A, Viale A, Dahi PB, et al. The effects of intestinal tract bacterial diversity on mortality following allogeneic hematopoietic stem cell transplantation. Blood. 2014;124(7):1174–1182.
- Kakihana K, Fujioka Y, Suda W, Najima Y, Kuwata G, Sasajima S, Mimura I, Morita H, Sugiyama D, Nishikawa H, et al. Fecal microbiota transplantation for patients with steroid-resistant acute graft-versus-host disease of the gut. Blood. 2016;128(16):2083–2088.
- Zuo T, Wong SH, Lam K, Lui R, Cheung K, Tang W, Ching JYL, Chan PKS, Chan MCW, Wu JCY, et al. Bacteriophage transfer during faecal microbiota transplantation in Clostridium difficile infection is associated with treatment outcome. Gut. 2018;67(4):634–643.
- Przerwa A, Zimecki M, Switała-Jeleń K, Dabrowska K, Krawczyk E, Łuczak M, Weber-Dabrowska B, Syper D, Miedzybrodzki R, Górski A. Effects of bacteriophages on free radical production and phagocytic functions. Med Microbiol Immunol. 2006;195(3):143–150.
- Miedzybrodzki R, Switala-Jelen K, Fortuna W, Weber-Dabrowska B, Przerwa A, Lusiak-Szelachowska M, Dabrowska K, Kurzepa A, Boratynski J, Syper D, et al. Bacteriophage preparation inhibition of reactive oxygen species generation by endotoxin-stimulated polymorphonuclear leukocytes. Virus Res. 2008;131(2):233–242.
- Górski A, Dąbrowska K, Międzybrodzki R, Weber-Dąbrowska B, Łusiak-Szelachowska M, Jończyk-Matysiak E, Borysowski J. Phages and immunomodulation. Future Microbiol. 2017;12(10):905–914.
- Sun Y, Li Y, Wu B, Wang J, Lu X, Qu S, Weng J, Feng B. Biological responses to M13 bacteriophage modified titanium surfaces in vitro. Acta Biomater. 2017;58:527–538.
- Liang Y, Ma S, Zhang Y, Wang Y, Cheng Q, Wu Y, Jin Y, Zheng D, Wu D, Liu H. IL-1β and TLR4 signaling are involved in the aggravated murine acute graft-versus-host disease caused by delayed bortezomib administration. J Immunol. 2014;192(3):1277–1285.
- Park S, Shin HJ, Shah M, Cho HY, Anwar MA, Achek A, Kwon HK, Lee B, Yoo TH, Choi S. TLR4/MD2 specific peptides stalled in vivo LPS-induced immune exacerbation. Biomaterials. 2017;126:49–60.
- Miernikiewicz P, Kłopot A, Soluch R, Szkuta P, Kęska W, Hodyra-Stefaniak K, Konopka A, Nowak M, Lecion D, Kaźmierczak Z, et al. T4 phage tail adhesin Gp12 Counteracts LPS-induced inflammation in vivo. Front Microbiol. 2016;7:1112.
- Jubair WK, Hendrickson JD, Severs EL, Schulz HM, Adhikari S, Ir D, Pagan JD, Anthony RM, Robertson CE, Frank DN, et al. Modulation of inflammatory arthritis in mice by gut microbiota through mucosal inflammation and autoantibody generation. Arthritis Rheumatol. 2018;70(8):1220–1233.
- Chen J, Wright K, Davis JM, Jeraldo P, Marietta EV, Murray J, Nelson H, Matteson EL, Taneja V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016;8(1):43.
- Mangalea MR, Paez-Espino D, Kieft K, Chatterjee A, Chriswell ME, Seifert JA, Feser ML, Demoruelle MK, Sakatos A, Anantharaman K, et al. Individuals at risk for rheumatoid arthritis harbor differential intestinal bacteriophage communities with distinct metabolic potential. Cell Host Microbe. 2021;29(5):726–739.e725.
- Liu X, Zeng B, Zhang J, Li W, Mou F, Wang H, Zou Q, Zhong B, Wu L, Wei H, et al. Role of the gut microbiome in modulating arthritis progression in mice. Sci Rep. 2016;6(1):30594.
- Nakao R, Senpuku H, Watanabe H. Porphyromonas gingivalis galE is involved in lipopolysaccharide O-antigen synthesis and biofilm formation. Infect Immun. 2006;74(11):6145–6153.
- Sinha A, Maurice CF. A phingerprint for rheumatoid arthritis development? Cell Host Microbe. 2021;29(5):678–680.
- de Goffau MC, Luopajärvi K, Knip M, Ilonen J, Ruohtula T, Härkönen T, Orivuori L, Hakala S, Welling GW, Harmsen HJ, et al. Fecal microbiota composition differs between children with β-cell autoimmunity and those without. Diabetes. 2013;62(4):1238–1244.
- Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, Liang S, Zhang W, Guan Y, Shen D, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. 2012;490(7418):55–60.
- Karlsson FH, Tremaroli V, Nookaew I, Bergström G, Behre CJ, Fagerberg B, Nielsen J, Bäckhed F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature. 2013;498(7452):99–103.
- Sato J, Kanazawa A, Ikeda F, Yoshihara T, Goto H, Abe H, Komiya K, Kawaguchi M, Shimizu T, Ogihara T, et al. Gut dysbiosis and detection of “live gut bacteria” in blood of Japanese patients with type 2 diabetes. Diabetes Care. 2014;37(8):2343–2350.
- Tolhurst G, Heffron H, Lam YS, Parker HE, Habib AM, Diakogiannaki E, Cameron J, Grosse J, Reimann F, Gribble FM. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes. 2012;61(2):364–371.
- Everard A, Cani PD. Gut microbiota and GLP-1. Rev Endocr Metab Disord. 2014;15(3):189–196.
- Ma Y, You X, Mai G, Tokuyasu T, Liu C. A human gut phage catalog correlates the gut phageome with type 2 diabetes. Microbiome. 2018;6(1):24.
- Tetz G, Brown SM, Hao Y, Tetz V. Type 1 diabetes: an association between autoimmunity, the dynamics of gut amyloid-producing E. coli and their phages. Sci Rep. 2019;9(1):9685.
- Tetz G, Tetz V. Bacteriophages as new human viral pathogens. Microorganisms. 2018;6(2):54.
- Chen Q, Ma X, Li C, Shen Y, Zhu W, Zhang Y, Guo X, Zhou J, Liu C. Enteric phageome alterations in patients with type 2 diabetes. Front Cell Infect Microbiol. 2020;10:575084.
- Zhao G, Vatanen T, Droit L, Park A, Kostic AD, Poon TW, Vlamakis H, Siljander H, Härkönen T, Hämäläinen AM, et al. Intestinal virome changes precede autoimmunity in type I diabetes-susceptible children. Proc Natl Acad Sci U S A. 2017;114(30):E6166–e6175.
- Rasmussen TS, Mentzel CMJ, Kot W, Castro-Mejía JL, Zuffa S, Swann JR, Hansen LH, Vogensen FK, Hansen AK, Nielsen DS. Faecal virome transplantation decreases symptoms of type 2 diabetes and obesity in a murine model. Gut. 2020;69(12):2122–2130.
- Manrique P, Zhu Y, van der Oost J, Herrema H, Nieuwdorp M, de Vos WM, Young M. Gut bacteriophage dynamics during fecal microbial transplantation in subjects with metabolic syndrome. Gut Microbes. 2021;13(1):1–15.
- Manohar P, Tamhankar AJ, Lundborg CS, Nachimuthu R. Therapeutic characterization and efficacy of bacteriophage cocktails infecting Escherichia coli, klebsiella pneumoniae, and enterobacter species. Front Microbiol. 2019;10:574.
- Yen M, Cairns LS, Camilli A. A cocktail of three virulent bacteriophages prevents Vibrio cholerae infection in animal models. Nat Commun. 2017;8(1):14187.
- Selle K, Fletcher JR, Tuson H, Schmitt DS, McMillan L, Vridhambal GS, Rivera AJ, Montgomery SA, Fortier LC, Barrangou R, et al. In vivo targeting of clostridioides difficile using phage-delivered CRISPR-Cas3 antimicrobials. mBio. 2020;11(2)
- Taha OA, Connerton PL, Connerton IF, El-Shibiny A. Bacteriophage ZCKP1: a potential treatment for klebsiella pneumoniae isolated from diabetic foot patients. Front Microbiol. 2018;9:2127.
- Roach DR, Leung CY, Henry M, Morello E, Singh D, Di Santo JP, Weitz JS, Debarbieux L. Synergy between the host immune system and bacteriophage is essential for successful phage therapy against an acute respiratory pathogen. Cell Host Microbe. 2017;22(1):38–47.e34.
- Liu X, Sui J, Li C, Peng X, Wang Q, Jiang N, Xu Q, Wang L, Lin J, Zhao G. Preparation of a nanobody specific to dectin 1 and its anti-inflammatory effects on fungal keratitis. Int J Nanomedicine. 2022;17:537–551.
- Jault P, Leclerc T, Jennes S, Pirnay JP, Que YA, Resch G, Rousseau AF, Ravat F, Carsin H, Le Floch R, et al. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): a randomised, controlled, double-blind phase 1/2 trial. Lancet Infect Dis. 2019;19(1):35–45.
- Sarker SA, Sultana S, Reuteler G, Moine D, Descombes P, Charton F, Bourdin G, McCallin S, Ngom-Bru C, Neville T, et al. Oral phage therapy of acute bacterial diarrhea with two coliphage preparations: a randomized trial in children from Bangladesh. EBioMedicine. 2016;4:124–137.
- Schooley RT, Biswas B, Gill JJ, Hernandez-Morales A, Lancaster J, Lessor L, Barr JJ, Reed SL, Rohwer F, Benler S, et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant acinetobacter baumannii infection. Antimicrob Agents Chemother. 2017;61(10)
- González-Cano P, Gamage LNA, Marciniuk K, Hayes C, Napper S, Hayes S, Griebel PJ. Lambda display phage as a mucosal vaccine delivery vehicle for peptide antigens. Vaccine. 2017;35(52):7256–7263.
- Gomes-Neto JF, Sartorius R, Canto FB, Almeida TS, Dias AA, Barbosa CD, Melo GA, Oliveira AC, Aguiar PN, Machado CR, et al. Vaccination with recombinant filamentous fd phages against parasite infection requires TLR9 expression. Front Immunol. 2018;9:1173.
- Hess KL, Andorko JI, Tostanoski LH, Jewell CM. Polyplexes assembled from self-peptides and regulatory nucleic acids blunt toll-like receptor signaling to combat autoimmunity. Biomaterials. 2017;118:51–62.
- Liu S, Jiang Q, Zhao X, Zhao R, Wang Y, Wang Y, Liu J, Shang Y, Zhao S, Wu T, et al. A DNA nanodevice-based vaccine for cancer immunotherapy. Nat Mater. 2021;20(3):421–430.
- White RA. 3rd: the future of virology is synthetic. mSystems. 2021;6(4):e0077021.
- Ando H, Lemire S, Pires DP, Lu TK. Engineering modular viral scaffolds for targeted bacterial population editing. Cell Syst. 2015;1(3):187–196.
- Citorik RJ, Mimee M, Lu TK. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat Biotechnol. 2014;32(11):1141–1145.
- Jalasvuori M, Koskinen K. Extending the hosts of Tectiviridae into four additional genera of gram-positive bacteria and more diverse bacillus species. Virology. 2018;518:136–142.
- Chan BK, Sistrom M, Wertz JE, Kortright KE, Narayan D, Turner PE. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci Rep. 2016;6(1):26717.
- Fong K, Mu K, Rheault JG, Levesque RC, Kitts DD, Delaquis P, Goodridge L, Wang S. Bacteriophage-insensitive mutants of antimicrobial-resistant salmonella enterica are altered in their tetracycline resistance and virulence in Caco-2 intestinal cells. Int J Mol Sci. 2020;21(5):1883.
- Sunderland KS, Yang M, Mao C. Phage-enabled nanomedicine: from probes to therapeutics in precision medicine. Angew Chem Int Ed Engl. 2017;56(8):1964–1992.
- Kovacs EW, Hooker JM, Romanini DW, Holder PG, Berry KE, Francis MB. Dual-surface-modified bacteriophage MS2 as an ideal scaffold for a viral capsid-based drug delivery system. Bioconjug Chem. 2007;18(4):1140–1147.
- Ngweniform P, Abbineni G, Cao B, Mao C. Self-assembly of drug-loaded liposomes on genetically engineered target-recognizing M13 phage: a novel nanocarrier for targeted drug delivery. Small. 2009;5(17):1963–1969.
- Pinto AM, Faustino A, Pastrana LM, Bañobre-López M, Sillankorva S. Pseudomonas aeruginosa PAO 1 in vitro time-kill kinetics using single phages and phage formulations-modulating death, adaptation, and resistance. Antibiotics (Basel). 2021;10(7)
- Göller PC, Elsener T, Lorgé D, Radulovic N, Bernardi V, Naumann A, Amri N, Khatchatourova E, Coutinho FH, Loessner MJ, et al. Multi-species host range of staphylococcal phages isolated from wastewater. Nat Commun. 2021;12(1):6965.
- Gill JJ, Pacan JC, Carson ME, Leslie KE, Griffiths MW, Sabour PM. Efficacy and pharmacokinetics of bacteriophage therapy in treatment of subclinical Staphylococcus aureus mastitis in lactating dairy cattle. Antimicrob Agents Chemother. 2006;50(9):2912–2918.
- Tetz GV, Ruggles KV, Zhou H, Heguy A, Tsirigos A, Tetz V. Bacteriophages as potential new mammalian pathogens. Sci Rep. 2017;7(1):7043.
- Luong T, Salabarria AC, Edwards RA, Roach DR. Standardized bacteriophage purification for personalized phage therapy. Nat Protoc. 2020;15(9):2867–2890.