
ABSTRACT
The facultative anaerobic Gram-positive bacterium Enterococcus faecium is a ubiquitous member of the human gut microbiota. However, it has gradually evolved into a pathogenic and multidrug resistant lineage that causes nosocomial infections. The establishment of high-level intestinal colonization by enterococci represents a critical step of infection. The majority of current research on Enterococcus has been conducted under aerobic conditions, while limited attention has been given to its physiological characteristics in anaerobic environments, which reflects its natural colonization niche in the gut. In this study, a high-density transposon mutant library containing 26,620 distinct insertion sites was constructed. Tn-seq analysis identified six genes that significantly contribute to growth under anaerobic conditions. Under anaerobic conditions, deletion of sufB (encoding Fe-S cluster assembly protein B) results in more extensive and significant impairments on carbohydrate metabolism compared to aerobic conditions. Consistently, the pathways involved in this utilization-restricted carbohydrates were mostly expressed at significantly lower levels in mutant compared to wild-type under anaerobic conditions. Moreover, deletion of sufB or pflA (encoding pyruvate formate lyase-activating protein A) led to failure of gastrointestinal colonization in mice. These findings contribute to our understanding of the mechanisms by which E. faecium maintains proliferation under anaerobic conditions and establishes colonization in the gut.
1. Introduction
E. faecium is a Gram-positive bacterium and a member of the normal intestinal flora of animals.Citation1 Additionally, there is substantiated evidence suggesting the presence of enterococci in plants, insects,Citation2,Citation3 water, soil, and mountainous regions.Citation4 Enterococcus species exhibit adaptability to both aerobic natural habitats and anaerobic intestinal environments. This bacterium exhibits an exceptional capacity to endure adverse environmental circumstances, such as elevated temperatures and heightened salt concentrations.Citation5 Moreover, Enterococcus species exhibit exceptional resistance to various chemical disinfectants, such as chlorine, glutaraldehyde, and alcohol.Citation6 These distinct characteristics contribute to the persistence of E. faecium within the modern hospital environment and highlight its ability to adapt during long-term co-evolutionary processes. Over the past three decades, E. faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumanii, Pseudomonas aeruginosa, and Enterobacter, which are collectively known as ESKAPE, have been identified by the American Society for Infectious Diseases as significant causes of treatment challenges and mortality worldwide.Citation7 E. faecium has emerged as a multidrug-resistant pathogen that is specifically associated with healthcare settings.Citation8
The establishment of a heightened level of enterococcal colonization within the intestines is a critical stage in a process that has the potential to result in nosocomial infections.Citation9 The capacity of bacteria to adapt to stress within the intestinal tract is crucial for survival and successful colonization. Throughout the gastrointestinal phase, intestinal bacteria confront the host’s formidable physiochemical defenses, which include the acidic pH of the stomach, heightened osmolarity and bile salts in the upper small intestine, as well as anaerobic conditions prevailing within the intestine.Citation10 The gut is characterized by anaerobic conditions, resulting in the predominance of anaerobic microorganisms in the intestinal microbiota of mammals.Citation11 The proportion of strict anaerobes in the gut increases progressively from the proximal to distal regions, constituting 99% of bacterial species within the colon.Citation12 The oxygen partial pressure in the colonic mucosa has been observed to be less than 25% of atmospheric oxygen concentration.Citation13 Despite being less abundant, facultative anaerobes such as Escherichia coli and Enterococcus are extensively present in the mammalian intestinal tract.Citation14 These organisms can survive under both anaerobic and aerobic conditions, and the concentration of oxygen plays a regulatory role in their growth and metabolism.Citation15 For instance, oxygen levels in the intestinal wall play a significant role in the virulence of pathogenic bacterium Shigella.Citation16 Despite the vital importance of oxygen levels in the colonization and virulence of intestinal microbes, limited research has been conducted in this particular field.
The microbiota could play a crucial role in modifying the gut environment, indirectly enhancing resistance against enteric pathogens and pathobionts.Citation17 A notable example of this phenomenon is the ability of symbiotic bacteria to establish and maintain intestinal hypoxia, which limits the proliferation of facultative anaerobic pathogens.Citation18 Consequently, facultative anaerobes in symbiosis can competitively acquire or sequester residual oxygen within the gut, further restricting pathogen colonization.Citation16 Furthermore, it has been observed that other symbionts can compete for anaerobic respiration substrates in the absence of oxygen.Citation19–21 Previous studies have investigated the impact of oxygen limitation on the abundance of Enterobacteriaceae in the gastrointestinal tract, revealing that a scarcity of respiratory electron acceptors limits the growth of Enterobacteriaceae within a balanced gut-associated microbial communityCitation22 and gastrointestinal colonization. As a facultative anaerobic bacterium, E. faecium grows most rapidly when respiring oxygen and switches to anaerobic respiration in the absence of oxygen.Citation23 To thrive, Enterococcus must undergo adaptations in an environment lacking oxygen. However, the specific genes involved in this adaptive process and their respective functions have not been extensively documented.
Transposon sequencing (Tn-seq) is a bacterial functional genomics research technique that combines transposon mutagenesis with high-throughput sequencing.Citation24 Tn-seq has emerged as a genome-wide technique that establishes a direct association between phenotype and genotype.Citation25 This technique facilitates the simultaneous pooling of millions of diverse mutant strains, enabling high-throughput identification and correlation analysis of gene function. Additionally, it allows for quantitative monitoring of variations in the relative abundance of mutants acquired during pre- and post-tests. Through Tn-seq, we can comprehensively identify conditionally essential genes in bacteria within specific growth environments and elucidate the functional roles of bacterial genes at a genome-wide scale.Citation26–29
In this study, a high-density transposon library was constructed for E. faecium E980, allowing for comprehensive identification of genes associated with anaerobic conditions through Tn-seq analysis. As a result, suf gene cluster was identified. This gene cluster is widely regarded as the oldest known pathway for the biogenesis of iron-sulfur (Fe-S) cluster and is supposed to be involved in early anaerobic life forms.Citation30 These Fe-S cluster play crucial roles as essential metal cofactors in various vital biological processes, including respiration, photosynthesis, nitrogen fixation, and DNA repair.Citation31,Citation32 The process of Fe-S biogenesis involves several steps: SufS facilitates mobilization of sulfide (in the form of persulfide) from L-cysteine, SufD acquires iron, and SufU transfers the obtained sulfur to the target scaffold proteins (SufB and SufC),Citation33 as shown in . Previous studies have also reported the mechanism of Fe-S cluster generation by Suf system in Enterococcus,Citation34–36 but the association of Fe-S cluster with bacterial growth characteristics and host interaction has not been described. In the present study, suf gene cluster of E. faecium was characterized, and Fe-S synthesis pathway encoded by suf genes was shown to play a significant role in anaerobic growth and gastrointestinal colonization of E. faecium.
2. Materials and methods
2.1. Bacterial strains, media, and growth conditions
Bacterial strains and plasmids used in this study are listed in Table S1. All strains were grown at 37°C in brain heart infusion (BHI) broth, and antibiotics (Solarbio) were used at the following concentrations: gentamicin, 200 µg/mL; spectinomycin, 100 µg/mL.
2.2. Construction of a mariner transposon mutant library in E. faecium
To create a high-density transposon mutant library, the temperature-sensitive plasmid pGPA2Citation37 was electroporated into E. faecium E980. The mutant library was generated according to previously described methods.Citation38 In brief, E980 containing plasmid were grown in BHI broth supplemented with chloramphenicol at 30°C overnight, after which cultures (200 μL) were added to BHI broth supplemented with gentamicin (25 μg/mL) and nisin (25 ng/mL) overnight. Then, cultures were incubated in BHI broth without antibiotics at 37°C for two successive passages. Subsequently, cultures were stored at −80°C in 50% (v/v) glycerol.
2.3. Tn-seq analysis of conditionally essential genes involved in anaerobic conditions
Tn-seq, a high-throughput tool for the functional genomic study of pathogens, has previously been described in detail.Citation39 In this study, we used this technique to perform genome-wide identification of conditionally essential genes of E. faecium under anaerobic treatment. The procedure was similar to that described previously.Citation40 E. faecium mariner-based transposon mutant library was grown in BHI broth supplemented with 25 µg/mL gentamycin to the stationary growth phase.
Subsequently, cultures were inoculated anaerobically at a concentration of 1 × 107 CFUs/mL in BHI broth at 37°C for 10–12 h in an anaerobic incubator. Similarly, the control group was placed in a warm air incubator at 37°C for 10–12 h. Then, bacteria were collected immediately by centrifugation, and a genomic DNA kit (Qiagen) was used for genomic DNA extraction. Sample libraries were prepared and analyzed by barcoded Tn-seq as described previously.Citation41 Library preparation and sequencing were performed on the Illumina HiSeq PE150 platform (Personalbio, Shanghai, China), generating an average of 1 G high-quality sequencing reads per sample. This experiment was performed in triplicate.
2.4. Bioinformatics analysis of Tn-seq data
Tn-seq data analysis was performed as described previously.Citation42 After Illumina sequencing, the raw reads were split into groups based on their barcodes using Fastp.Citation43 Sixteen nucleotide fragments of each read, corresponding to E980 genomic sequence flanking transposon, were mapped to E980 genome using Bowtie 2.Citation44 Then, Integrative Genomics Viewer (IGV) was used to sort and count results of the alignment. Next, the read 6 counts of each gene were normalized by the following formula: RPTAM = (number of reads mapped to a gene × 106/(total mapped input reads in the sample × number of TA sites in this gene). Cyber-TCitation45 (http://cybert.microarray.ics.uci.edu/) was used for statistical analysis of RPTAM values between different groups. A p value < .05 was considered to indicate statistical significance. Only genes with a significant and unique association with either sensitivity or tolerance were identified.
2.5. Phylogenetic and in silico analysis of suf gene cluster in E. faecium E980
The BLAST tool of the NCBI (https://www.ncbi.nlm.nih.gov/) was used for homology analysis of suf gene cluster.
The amino acid sequences of SufB (Fe-S cluster assembly protein B) and PflA (pyruvate formate lyase-activating protein A) in E. faecium E980 were submitted to structural prediction using the AlphaFold2 server.Citation46 Structural alignment of SufB (or PflA) in E. coli and E980 was performed using the TM-align serverCitation47 for structural analysis. Sequence alignment was performed using the CLUSTALW server,Citation48 and visualization was achieved using the ESPript 3.0 server.Citation49
To explore which Fe-S cluster-containing proteins downstream were affected by impaired Suf system, two methods were performed. Firstly, the homologous Fe-S cluster-containing protein in E. faecium was analyzed based on the literature report of Fe-S cluster-containing protein in E. coli.Citation50 Moreover, the UniProt database was used to directly search for Fe-S cluster-containing proteins in E. faecium. The identified proteins by above methods were subsequently mapped to E. faecium E980 genome. We employed these two methods to identify Fe-S cluster-containing proteins potentially implicated in E. faecium E980 strain.
2.6. Construction and complementation of E. faecium E980 mutant strains
Deletion of sufB was performed in E980 strain through the recombination system mediated by plasmid pWS3.Citation51 A schematic diagram of the recombination system for the construction of gene knockout mutant in E. faecium is provided in Supplementary Materials (Figure S1).The primers used are listed in Table S1. Plasmid pWS3 was digested with Sma I, and PCR fragments (gene_up, gene_dn, gm) were ligated by using NovoRec plus One step PCR Cloning Kit (Novoprotein Scientific, Inc., Shanghai, China). Plasmid pWS3_gene_gm was transformed into E980 competent cells using the electroporation method. As shown in Figure S2, a transformant containing plasmid was grown overnight in BHI broth at 30°C supplemented with gentamicin. Cell cultures were then diluted 10,000-fold in prewarmed BHI broth and grown at 37°C overnight without antibiotics. Cells were then plated on BHI agar plates with gentamicin and incubated at 37°C. Colonies were then restreaked on BHI agar plates with spectinomycin and BHI agar plates with gentamicin, respectively. The gentamicin-resistant but spectinomycin-susceptible colonies were supposed to be marked deletion mutants. Thus, marked deletion mutants were obtained. Deletion of pflA was performed via the same method. All plasmid constructs and gene deletions were confirmed by DNA sequencing.
Genetic complementation of mutants was performed as previously described.Citation52 To complement ΔsufB::gm, sufB gene with its promoter was cloned and inserted into pMSP3535 shuttle plasmid. PCR products were cloned and inserted into BamH I site of pMSP3535 vector using NovoRec® plus One-step PCR Cloning Kit (Novoprotein), generating plasmid pMSP3535-sufB. Positive transformants were obtained as previously described. Recombinant plasmids were subsequently introduced into ΔsufB::gm host strains by electroporation, after which transformants were selected at 37°C on BHI plates supplemented with erythromycin and lincomycin. Other complement strains were constructed via the same method.
2.7. Determination of growth curves
A growth curve was generated for wild-type (WT), isogenic mutant, and complemented strains under both aerobic and anaerobic conditions. Anaerobic operation was conducted within a specialized anaerobic incubator. Strains were grown overnight in BHI. 50 μL of overnight culture was inoculated into 50 mL of solution. Cultures were incubated in an incubator at 37°C, and absorbance at 600 nm (OD600) was recorded every hour for 9 hours. Each experiment was carried out in triplicate.
2.8. Transcriptome comparison between WT and ΔsufB::gm strains under aerobic and anaerobic conditions
WT and ΔsufB::gm were incubated in BHI broth under aerobic and anaerobic conditions, respectively, for 6 hours. Bacterial cells were centrifuged at 10,000 rpm for 1 min and snap-frozen in liquid nitrogen. RNA extraction and quantification were performed as described in previous studies.Citation53 Then, amplified and purified RNA-Seq libraries were sequenced on the NovaSeq 6000 platform (Illumina, San Diego, CA, USA), and 150-bp paired-end reads were generated. Differentially expressed gene (DEG) analysis and enrichment analysis (Gene Ontology [GO] analysis) were performed according to previous methods.Citation53
2.9. qPCR validation of RNA-Seq experiments
cDNA was synthesized according to instructions of the PrimeScript RT Reagent Kit with gDNA Eraser (Takara, Beijing, China). Then, qPCR was conducted using TB Green® Premix Ex Taq™ (Tli RNaseH Plus) Kit (Thermo Fisher Scientific, Waltham, MA, USA) and Applied Biosystems™ QuantStudio™ 3 instrument (Thermo Fisher Scientific, Waltham, Massachusetts, USA). Transcript levels of assayed genes relative to divIVA were calculated using QuantStudioTM Design & Analysis Software 1.3.1 (Thermo Fisher Scientific, Waltham, MA, USA). Data analysis was performed using the 2 −ΔΔCt method. Three biological replicates were performed for each group.
2.10. Carbohydrates metabolism test
To assess the metabolic capacities of WT strain, its isogenic mutants, and complemented strain under both aerobic and anaerobic conditions in relation to various sugars, namely, glucose, trehalose, sucrose, raffinose, arabinose, mannitol, cellobiose, galactose, lactose, xylose, xylitol, fructose, L -rhamnose, maltose, sorbitol, inulin and mannose, 17 microbiochemical identification tubes (Haibo, China) were employed. A total of 1 × 108 CFUs of WT strain, its isogenic mutants, and complemented strains were inoculated with 300 μL of microbiochemical identification reagent, BHI broth was used for overnight culture of all strains. The results are recorded at two-hour intervals within a 24-hour period, and at twelve-hour intervals between 24 and 48 hours.
2.11. Intestinal colonization in mice
Colonization of intestines of mice was conducted using E. faecium strains following a previously described method with certain modifications.Citation38,Citation54,Citation55 Specifically, before antibiotic treatment, 10 BALB/c mice were subjected to environmental adaptation for one week. seven-week SPF mice were subjected to a 3-day decolonization period via the oral administration of four antibiotics once daily.Citation56 Mice were allowed to drink water supplemented with four antibiotics (1 g/L) ad libitum for 7 days. Enterococcus in fecal pellets was not detected after antibiotic therapy. Mice were then inoculated with 1 × 108 CFUs in 300 μL of phosphate-buffered saline (PBS), which was an equal ratio of WT and ΔsufB::gm; both were cultured separately overnight in BHI broth. Feeding was performed as previously described with contaminated food.Citation38,Citation54,Citation55 Pfizer Enterococcus Selective Agar (PESA; These plates support the growth of both WT and mutants) or PESA supplemented with 5 μg/mL gentamicin was used to plate the diluted inoculums. The viable counts for mutants and WT were measured in feces samples at 1, 3, 5, and 7 days after colonization. We euthanized mice after they had colonized for 7 days. Samples were collected, weighed, and homogenized in 10 volumes of PBS after being detached from cecum and colon, as shown in . To determine viable counts, serial dilutions of homogenates were performed, and subsequent plating on PESA or PESA supplemented with 5 μg/mL gentamicin was performed. Cells were incubated at 37°C for 2 days; subsequently, colonies were counted, and identity of strains were confirmed via PCR. ΔpflA::gm was treated similarly. The enteric competition test of WT and ΔsufB::gm + sufB was the same as enteric competition test of WT and mutant strains in the preliminary treatment. Mice were then inoculated with 1 × 108 CFUs in 300 μL of PBS, which was an equal ratio of WT and ΔsufB::gm + sufB; both were cultured separately overnight in BHI broth. The viable counts for ΔsufB::gm + sufB and WT were measured in feces samples at 3, 5, and 7 days post colonization. ΔpflA::gm was treated similarly. We conducted a paired two-tailed Student’s t test to determine whether differences in the log-transformed data between WT and mutant strains were statistically significant. A p value < .05 was considered to indicate statistical significance. The Animal Care and Use Committee of Linyi University (Linyi, China) approved all the experiments.
2.12. Statistical analysis
All analyses of significance were performed using one-way analysis of variance (ANOVA) and Duncan’s multiple range test (DMRT). In all the analysis, only when p < .05 the data were considered to be statistically significant. All tests were conducted in triplicate.
3. Results
3.1. Construction and evaluation of a high-density transposon mutation library in E. faecium
We constructed a mariner-based transposon insertion library in E. faecium E980, and the quality of this mutation library was evaluated by Tn-seq analysis. The key steps involved in Tn-seq are illustrated in . The sequencing results revealed that each sample exhibited a minimum of 15 million Tn-seq readings, with no discernible presence of biased transposon insertion sites. The transposon distribution exhibited a uniform and high-density pattern, as shown in , with a total of 26,620 distinct insertion sites identified across the entire genome (Table S2). Subsequently, a comprehensive analysis of Tn-seq data was conducted to discern the genetic determinants associated with anaerobic conditions.
Figure 1. Tn-seq analysis for identification of functional genes under anaerobic conditions in E. faecium.
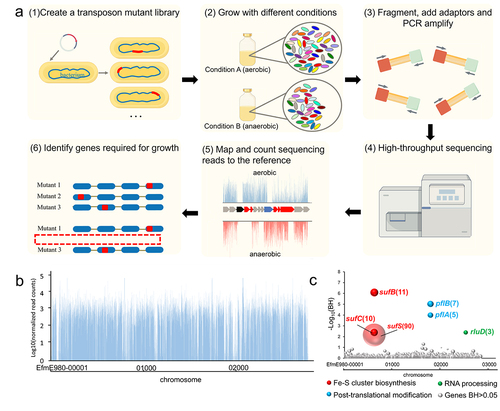
3.2. Identification of genetic determinants involved in anaerobic growth by Tn-seq
To identify genes required for growth under anaerobic conditions in E. faecium, Tn-seq was performed to determine which mutants were selectively lost during anaerobic growth. The involvement of six genes (Benjamini – Hochberg corrected p value (BH) < .05) under anaerobic conditions was identified ( and Table S2). Genes that had the most pronounced effect on the growth of E. faecium under anaerobic conditions included genes involved in Fe-S cluster biosynthesis (sufB, sufC, sufS), genes associated with post-translational modification (rluD) and genes involved in RNA processing (pflA, pflB) ().
3.3. Revealing essential genes of E. faecium E980 through Tn-seq analysis
By employing the EL-ARTISIT approach for identification of essential genes in E980 cells cultivated in BHI medium, a total of 631 essential genes were identified in Table S3, constituting approximately 21.25% of the entire gene repertoire. Essential genes identified in this study were submitted to GO and KEGG functional enrichment analysis, which provided insights into biological processes that are crucial for survival of E. faecium.
According to GO enrichment analysis (), a Padj (corrected p value) < .05 indicated statistically significant enrichment in the GO functional terms. The set of essential genes exhibited significant enrichment in 88 biological process terms, 135 molecular function terms, and 13 cell component terms. The functions of essential genes are predominantly involved in translation, cell metabolism, and biosynthesis. These genes encompass conserved multi-component pathways essential for fundamental metabolic and structural functions in bacteria.
Figure 2. Essential gene of GO (a) and KEGG (b) enrichment analysis.
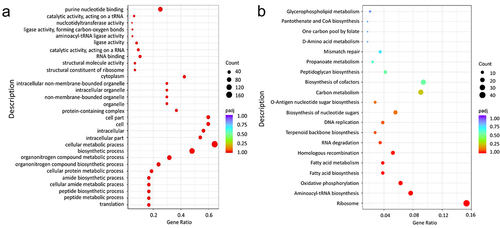
The KEGG pathway analysis is illustrated in . Essential genes exhibited enrichment in 48 biological metabolic pathways, with significant enrichment observed in 8 specific pathways, namely, ribosome pathway, aminoacyl-tRNA biosynthesis pathway, oxidative phosphorylation pathway, fatty acid synthesis pathway, fatty acid metabolism pathway, homologous recombination pathway, RNA degradation pathway, and terpenoid skeleton biosynthesis pathway. The enrichment of ribosome pathway was the most significant, with a notable abundance of enriched genes.
3.4. Phylogenetic and in silico analysis of suf gene cluster in E. faecium
A genome sequence analysis of E. faecium strain E980 was performed to identify suf gene cluster. The putative suf gene cluster included sufC, sufD, sufS, sufU, and sufB in E980 strain. To identify suf homologues in E980 genome, we searched for these homologues in 10 other species, namely, Enterococcus faecalis, Listeria monocytogenes, Bacillus subtilis, Salmonella typhimurium, E. coli, Lactococcus lactis, Streptococcus pyogenes, Prochlorococcus marinus, Methanocaldococcus vulcanius, and Metallosphaera cuprina. Among these bacteria, Prochlorococcus marinus is a bacterium with a simple structure; Methanocaldococcus vulcanius and Metallosphaera cuprina are archaea. The findings are illustrated in . The proteins encoded by this gene cluster are homologous to Suf proteins of E. faecalis V583 (amino acid identity: 76%–95%), B. subtilis 168 (45%–74%), L. monocytogenes EGD-e (48%–76%), S. typh-imurium LT2 (27%–54%), E. coli K12 (25%–53%), L. lactis lac460 (56%–83%) and S. pyogenes M1 (30%–87%). P. marinus MIT 9515 (24%–52%), M. cuprina Ar-4 (40%–46%), and M. vulcanius M7 (29%–39%) were used. These results indicate that suf gene cluster are widely distributed across a diverse range of species.
Figure 3. Phylogenetic and in silico analysis of Suf pathway.
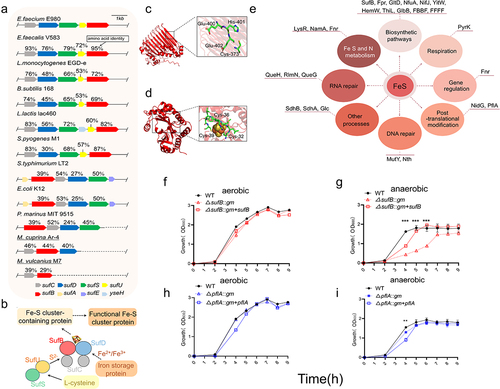
The predicted structure of E980 SufB protein exhibited a topological arrangement highly similar to that of SufB protein in E. coli (Figure S3a; TM-Score = 0.92,RMSD = 2.01). Additionally, the crucial functional sites 373C, 400E, 401 H, and 402E exhibited significant conservation (Figure S3c). These sites are predicted to be associated with Fe-S cluster biosynthetic function of SufB in E. coli.Citation57 Therefore, we hypothesized that these sites also play similar roles in Fe-S biosynthesis of E980 SufB. These sites are shown in E980 SufB structure (). Similarly, structural and sequence alignment analyses of PflA in E980 and E. coli were performed (Figure S3b, d; TM-Score = 0.95;RMSD = 1.73), and Fe-S cluster modification siteCitation58 of PflA was identified in E. coli. The potential Fe-S cluster modification sites of PflA in E980 were identified at positions 32C, 36C, and 39C (Figure S3d; ). These findings support a role for SufB and PflA in E. faecium E980 Fe-S cluster biosynthesis and modification.
To explore which possible pathways downstream were affected by impaired Suf system, Fe-S cluster-containing proteins in E980 strain were identified by bioinformatical analysis. A total of 45 Fe-S cluster-containing proteins in E. faecium were identified through homology comparison with E. coli Fe-S cluster proteins and retrieval from UniProt (Table S4). Subsequently, these 45 downstream proteins were further compared to E980 genome, resulting in a final set of 26 proteins. These 26 proteins are involved in various pathways, including biosynthesis; DNA repair; Fe, S, and N metabolism; gene regulation, post-translational modification; respiration; RNA modification; and other crucial cellular processes, as depicted in . We note that many known essential Fe-S cluster-modified proteins in E. coli have not been identified in E980, such as aconitate hydratase (AcnA) and fumarate hydratase (FumAB). Due to the limitations of this methodology, Fe-S cluster-containing proteins in E980 strain were not fully identified. Even so, these findings suggest that biosynthesis pathway of Fe-S cluster plays a pivotal role in diverse bacterial physiological processes.
The functional analysis of suf gene cluster in E. faecium support a role for SufB and PflA in E. faecium E980 Fe-S cluster biosynthesis and modification, as well as diverse bacterial physiological processes.
3.5. Growth of E. faecium under aerobic and anaerobic conditions
A growth curve was generated for WT, ΔsufB::gm, and ΔsufB::gm + sufB strains under both aerobic () and anaerobic () conditions. Similarly, growth curves of WT, ΔpflA::gm, and ΔpflA::gm + pflA strains were generated under both aerobic () and anaerobic () conditions. Both WT and isogenic mutants exhibited indistinguishable growth patterns under aerobic conditions, suggesting that the introduced mutations did not exert any discernible impact on aerobic growth. However, deletion of sufB and pflA genes had a significant impact on anaerobic growth of E. faecium E980. The difference between ΔpflA::gm strain and WT strain was only statistically significant at the fourth hour. The results indicate that sufB and pflA genes of E. faecium play crucial roles in E980 anaerobic growth.
3.6. Transcriptome comparison between WT and ΔsufB::gm strains under aerobic and anaerobic conditions
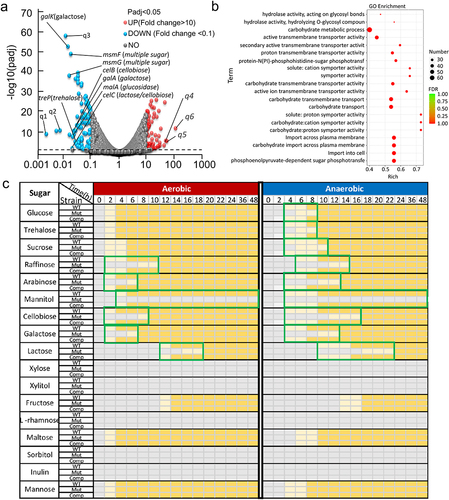
Transcriptomic analysis was also conducted to investigate the differences in gene expression between WT and ΔsufB::gm strains during anaerobic growth. Significant disparities in the heatmap (Figure S4) between two groups indicate notable variations in expression patterns between WT and ΔsufB::gm under anaerobic conditions. Further analysis of the data revealed 1099 DEGs for genes with a Padj value < .05 and a fold change > 2. Among the 1099 genes, 552 genes were upregulated, whereas 547 genes were downregulated (Table S6). Moreover, when we further increased the fold change requirement (FC > 10 or FC < 0.1), there were still 54 genes were upregulated, whereas 101 genes were downregulated (). Deletion of sufB resulted in a large proportion of DEGs under anaerobic conditions. GO enrichment analysis was subsequently performed to determine the functions of DEGs. The most significant top 20 GO terms of downregulated DEGs are shown in . Out of the total of 20 terms, 7 terms were associated with carbohydrate metabolism. The 7 terms included carbohydrate metabolic process, carbohydrate transmembrane transporter activity, carbohydrate transmembrane transport, carbohydrate transport, carbohydrate: cation symporter activity, carbohydrate: proton symporter activity, and carbohydrate import across plasma membrane. This result may indicate that deletion of sufB gene in E. faecium affects carbohydrate metabolism. Furthermore, the real-time PCR results confirmed reliability of RNA-Seq analysis, as shown in Figure S6.
Figure 4. Transcriptome comparison between WT and sufB mutant strains under anaerobic conditions.
In order to study the influence of sufB deletion in E. faecium, we conducted an RNA-Seq experiments to illustrate the transcriptional effect of sufB deletion on suf gene cluster in different conditions (Table S5-S8). Under aerobic conditions, O2 can damage the homeostatic state of Fe-S cluster, so the demand for Fe-S cluster is higher, which is also verified by the RNA-Seq results in this study. Under aerobic conditions, the impact of sufB deletion on Fe-S cluster generation may be small (), which is also illustrated by RNA-Seq: even if there is a high demand for Fe-S cluster under aerobic conditions, sufB deletion does not up-regulate the expression of sufCDSU gene.
3.7. Deletion of sufB results in extensive and significant impairments on carbohydrate metabolism
We have conducted a metabolic experiment on 17 different sugars. In general, all strains (WT, sufB mutant, and complement strain) grow faster in aerobic conditions compared to anaerobic conditions (). In aerobic conditions, although deletion of sufB does not affect the growth on glucose and several other sugars, the growth on raffinose, etc. especially mannitol is significantly suppressed, indicating that the essentiality of sufB depends on the particular carbon source. Under anaerobic conditions, the utilization of those unaffected sugars (including glucose, trehalose and sucrose) in aerobic conditions are impaired, suggesting that deletion of sufB under anaerobic conditions results in more extensive and significant impairments on carbohydrate metabolism compared to aerobic conditions. Interestingly, the type of “slow-growth carbohydrates” of sufB mutant were highly correlated with the function of down-regulated genes in transcriptome analysis as shown . The results established a link between phenotype and gene regulation, further revealing the mechanism by which sufB affects bacterial growth under anaerobic conditions.
3.8. sufB and pflA are required for colonization of intestinal tract by E. faecium
To further investigate the impact of sufB and pflA of E. faecium on intestinal colonization in mice, cell suspensions of E980 strain and ΔsufB::gm were mixed in a 1:1 ratio (108 CFUs of each strain) and administered to mice via oral infection (). Similarly, the other mutant, ΔpflA::gm, underwent identical treatment procedures. As shown in . E. faecium colonized intestinal tract of mice at high levels. There was a constant colonization rate, as the concentration in feces samples was always >109 CFUs/g. In competitive colonization assays, based on all stool samples collected across all time points, results showed that mutant isolate had significantly fewer viable cells compared to WT strain, as shown in . Mutant became almost undetectable by 3 days after infection. A completely lower level of colonization in cecum and colon was observed with mutant, as shown in , compared to parental strain. Cell suspensions of E980 strain and ΔsufB::gm + sufB were mixed in a 1:1 ratio (108 CFUs of each strain) and administered to mice via oral infection. As shown in , it can also be proved E. faecium colonized intestinal tract of mice at high levels. There was a constant colonization rate, as the concentration in feces samples was always >109 CFUs/g. Although the number of ΔsufB::gm + sufB is not as much as WT, it is far more than the number of ΔsufB::gm. Results showed that although it could not be fully complemented, complementary strain could rescue colonization to a fairly level which is significantly higher than mutants. Incomplete complementarity may be due to instability of plasmid in vivo environment or excessive copy number of plasmid, gene expression disorder and other predictable and common reasons. This result further confirms our previous findings.
Figure 5. Colonization of intestine by E. faecium in a mouse model.
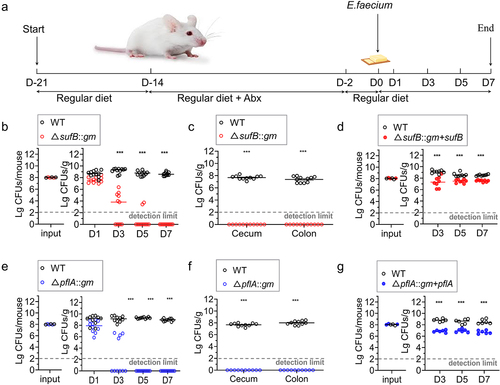
Based on these results, we concluded that deleting sufB and pflA in E. faecium E980 resulted in diminished intestinal colonization in mice.
4. Discussion
The majority of current research on Enterococcus has been conducted under aerobic conditions, with limited attention given to its physiological responses in anaerobic environments. The mechanisms by which Enterococcus maintain proliferation under anaerobic conditions are poorly understood, resulting in a significant knowledge gap. Therefore, it is important to explore the growth mechanism of E. faecium under anaerobic conditions. This study provided a comprehensive genome-wide identification of essential genetic determinants under anaerobic conditions in E. faecium.
In this study, Tn-seq methodology was used to elucidate genetic determinants involved in anaerobic growth. E. faecium, as a gut commensal, must have developed mechanisms to sense, respond to, and adapt to anaerobic environments.Citation52 Deletion of sufB and pflA genes had a significant impact on anaerobic growth of E. faecium E980 but had no discernible influence on its aerobic growth (), which verified the results of Tn-seq analysis. The role of sufB and pflA in gut colonization was demonstrated through competition experiments with WT and isogenic mutants. Our study emphasizes the significance of Fe-S cluster modification system in facilitating the proliferation of E. faecium under anaerobic conditions and gastrointestinal colonization.
The suf gene cluster was found to be widely spread across diverse bacterial species (). Fe – S cluster exhibited a nearly ubiquitous presence in both aerobic and anaerobic Archaea, Bacteria, and Eukarya.Citation50 Fe – S cluster are highly conserved cofactors in both prokaryotes and eukaryotes.Citation59 Suf system, an upstream pathway of Fe-S cluster biosynthesis, regulates numerous functional Fe – S cluster proteins and has prominent roles in multiple important cellular processes,Citation60 including respiration, central metabolism, gene regulation, RNA modification, DNA repair and replication. E. coli contains approximately 140 Fe-S proteins.Citation61 In this study, a total of 26 putative functional Fe – S cluster proteins were identified in E. faecium E980. Further research on E. faecium is anticipated to reveal additional functional Fe – S cluster proteins, particularly those associated with sugar metabolism, which could provide novel insights into carbohydrate-dependent mechanisms that contribute to intestinal colonization of gut microbes.
In order to find out whether E. faecium encodes any other Fe-S cluster biogenesis systems than Suf pathway, we performed bioinformatical analyses of Isc, Nif and recently identified MIS (minimal iron-sulfur) and SMS (SUF-like minimal system) based on protein sequence (blastp) and structure (HHpred), and confirmed that none of systems is present in E. faecium.Citation62 Therefore, results show that Suf system is only known Fe-S cluster biogenesis system in E. faecium. Our results of essential gene analysis showed that sufB is not essential while sufU is essential for E. faecium. The similar results were also observed in B. subtilis and S. aureus, in which the disruption of sufB, sufC, sufD or sufS was not lethal.Citation63–65 Therefore, we believe that although Suf system is necessary for bacterial viability, not every suf gene is required. The results suggest that the dependence of Fe-S cluster on sufB is different under aerobic and anaerobic conditions. Compared with aerobic conditions, the formation of effective Fe-S cluster under anaerobic conditions is more dependent on an intact (all genes of sufCDSUB cluster) suf gene cluster.
A transcriptome comparison between WT and sufB mutant strains demonstrated that suf deficiency resulted in a large proportion of DEGs under anaerobic conditions (), and there was notable enrichment in pathways related to sugar metabolism (). The suf-encoded Fe-S cluster biosynthesis pathway is an important means of protein modification, in which Fe-S cluster are modified into numerous proteins to regulate their function.Citation57 We hypothesized that proteins encoded by these genes involved in glycometabolism may exhibit impaired functionality due to deficient Fe-S cluster modification, thereby resulting in attenuated growth of suf-deficient mutant under anaerobic conditions ().
Bacteria use different energy metabolism pathways under aerobic and anaerobic conditions.Citation66 Under anaerobic conditions, bacteria do not have exogenous electron acceptors and cannot achieve complete tricarboxylic acid cycle (TCA cycle), and mainly perform fermentation to release energy lower compared to aerobic conditions. The formula was: glucose→2 pyruvate→2 acetate + ethanol + formate + lactate + 2 CO2. During this process, pyruvate is converted into acetyl-CoA and formate by pyruvate formate lyase (PflA), which is then metabolized to CO2 and H2 by formate dehydrogenase (Fdh).Citation67 Both PflA and Fdh are Fe-S cluster modified enzymes.Citation68,Citation69 Under anaerobic conditions, the restriction of Fe-S cluster generation caused by sufB deletion prevents PflA and Fdh from being modified by Fe-S cluster and thus cannot exert catalytic activity, which reduces the efficiency of process. In this study, the importance of pflA gene on anaerobic growth and colonization was also screened by Tn-seq, and verified by in vitro and in vivo experiments (), which further confirmed the important role of Fe-S cluster modification on anaerobic respiration. We performed a further RNA-Seq experiment to check gene regulation of WT E. faecium under aerobic and anaerobic conditions. As expected, the pathways relevant to pyruvate metabolism and TCA cycle were significantly down-regulated (Figure S5 and Table S7).
Deletion of sufB results in more extensive and significant impairments on carbohydrate metabolism under anaerobic conditions compared to aerobic conditions. Results are consistent with the growth experiment on BHI () in which the main carbon source is glucose. Mice were fed with commercial mice feed in which starch is the main carbon source. We attempted to grow E. faecium on the medium made with mice feed, however, probably because E. faecium cannot utilize starch directly as carbon source, Cell density could only reach to approximately 107, which is too low to perform growth experiment. However, in the gut starch could eventually be digested to glucose by the host and gut microbes. Therefore, the relative inefficiency of glucose metabolism of sufB mutant under anaerobic conditions could be one of factors that leads to the failure of intestinal colonization.
Our study revealed the pivotal role of Fe-S cluster biosynthesis system in intestinal colonization of E. faecium, thereby providing compelling evidence for investigating the key determinants of gut microbial colonization.
Supplemental Material
Download Zip (11.3 MB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The datasets generated during this study are available in the National Center for Biotechnology Information (NCBI) BioProject Repository https://www.ncbi.nlm.nih.gov/bioproject under BioProject PRJNA1061835.
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/19490976.2024.2359665
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
References
- Pöntinen AK, Top J, Arredondo-Alonso S, Tonkin-Hill G, Freitas AR, Novais C, Gladstone RA, Pesonen M, Meneses R, Pesonen H, et al. Apparent nosocomial adaptation of Enterococcus faecalis predates the modern hospital era. Nat Commun. 2021;12(1):1523. doi:10.1038/s41467-021-21749-5.
- García-Solache M, Rice LB. The Enterococcus: a model of adaptability to its environment. Clin Microbiol Rev. 2019;32(2):e00058–17. doi:10.1128/cmr.00058-18.
- Krawczyk B, Wityk P, Galecka M, Michalik M. The many faces of Enterococcus spp — commensal, probiotic, and opportunistic pathogen. Microorganisms. 2021;9(9):1900. doi:10.3390/microorganisms9091900.
- Gilmore MS, Willems RJ, Lebreton F. Enterococcus diversity, origins in nature, and gut colonization. Enterococci: from commensals to leading causes of drug-resistant infection. 2014. [Internet]. Available online. https://www.ncbi.nlm.nih.gov/books/NBK190427/.
- Facklam R, Collins M. Identification of Enterococcus species isolated from human infections by a conventional test scheme. J Clin Microbiol. 1989;27(4):731–734. doi:10.1128/jcm.27.4.731-734.1989.
- Bradley C, Fraise A. Heat and chemical resistance of enterococci. J Hosp Infect. 1996;34(3):191–196. doi:10.1016/S0195-6701(96)90065-1.
- Boucher HW, Talbot GH, Bradley JS, Edwards JE, Gilbert D, Rice LB, Scheld M, Spellberg B, Bartlett J. Bad bugs, no drugs: no ESKAPE! An update from the infectious diseases society of America. Clin Infect Dis. 2009;48(1):1–12. doi:10.1086/595011.
- Arias CA, Murray BE. The rise of the Enterococcus: beyond vancomycin resistance. Nat Rev Microbiol. 2012;10(4):266–278. doi:10.1038/nrmicro2761.
- Zhang XL, Bierschenk D, Top J, Anastasiou I, Bonten MJM, Willems RJL, van Schaik W, Dyer TD, Comuzzie A, Curran JE. Functional genomic analysis of bile salt resistance in Enterococcus faecium. BMC Genomics. 2013;6(1):14. doi:10.1186/1755-8794-6-14.
- Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science. 2005;307(5717):1915–1920. doi:10.1126/science.1104816.
- Jones SA, Gibson T, Maltby RC, Chowdhury FZ, Stewart V, Cohen PS, Conway T. Anaerobic respiration of Escherichia coli in the mouse intestine. Infect Immun. 2011;79(10):4218–4226. doi:10.1128/iai.05395-11.
- Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI. Molecular analysis of commensal host-microbial relationships in the intestine. Science. 2001;291(5505):881–884. doi:10.1126/science.291.5505.881.
- Wilson M. Microbial inhabitants of humans: their ecology and role in health and disease. Cambridge (UK): Cambridge University Press; 2005. p. 1–50. ISBN 9780511735080.
- Savage DC. Microbial ecology of the gastrointestinal tract. Annu Rev Microbiol. 1977;3(12):107–133. doi:10.1146/annurev.mi.31.100177.000543.
- Jones SA, Chowdhury FZ, Fabich AJ, Anderson A, Schreiner DM, House AL, Autieri SM, Leatham MP, Lins JJ, Jorgensen M, et al. Respiration of Escherichia coli in the mouse intestine. Infect Immun. 2007;75(10):4891–4899. doi:10.1128/iai.00484-07.
- Marteyn B, West NP, Browning DF, Cole JA, Shaw JG, Palm F, Mounier J, Prévost M-C, Sansonetti P, Tang CM, et al. Modulation of Shigella virulence in response to available oxygen in vivo. Nature. 2010;465(7296):355–U113. doi:10.1038/nature08970.
- Caballero-Flores G, Pickard JM, Núñez G. Microbiota-mediated colonization resistance: mechanisms and regulation. Nat Rev Microbiol. 2023;21(6):347–360. doi:10.1038/s41579-022-00833-7.
- Litvak Y, Byndloss MX, Tsolis RM, Bäumler AJ. Dysbiotic Proteobacteria expansion: a microbial signature of epithelial dysfunction. Curr Opin Microbiol. 2017;39:1–6. doi:10.1016/j.mib.2017.07.003.
- Winter SE, Winter MG, Xavier MN, Thiennimitr P, Poon V, Keestra AM, Laughlin RC, Gomez G, Wu J, Lawhon SD, et al. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science. 2013;339(6120):708–711. doi:10.1126/science.1232467.
- Spees AM, Wangdi T, Lopez CA, Kingsbury DD, Xavier MN, Winter SE, Tsolis RM, Bäumler AJ. Streptomycin-induced inflammation enhances Escherichia coli gut colonization through nitrate respiration. Mbio. 2013;4(4):e00430–13. doi:10.1128/mbio.00430-13.
- Herp S, Brugiroux S, Garzetti D, Ring D, Jochum LM, Beutler M, Eberl C, Hussain S, Walter S, Gerlach RG, et al. Mucispirillum schaedleri antagonizes salmonella virulence to protect mice against colitis. Cell Host Microbe. 2019;25(5):681–694.e8. doi:10.1016/j.chom.2019.03.004.
- Rivera-Chávez F, Lopez CA, Bäumler AJ. Oxygen as a driver of gut dysbiosis. Freeradicalbiomed. 2017;105:93–101. doi:10.1016/j.freeradbiomed.2016.09.022.
- Kelly CJ, Colgan SP. Breathless in the gut: implications of luminal O2 for microbial pathogenicity. Cell Host Microbe. 2016;19(4):427–428. doi:10.1016/j.chom.2016.03.014.
- van Opijnen T, Bodi KL, Camilli A. Tn-seq: high-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms. Nat Methods. 2009;6(10):767–U21. doi:10.1038/NMETH.1377.
- Cain AK, Barquist L, Goodman AL, Paulsen IT, Parkhill J, van Opijnen T. A decade of advances in transposon-insertion sequencing. Nat Rev Genet. 2020;21(9):526–540. doi:10.1038/s41576-020-0244-x.
- Powell JE, Leonard SP, Kwong WK, Engel P, Moran NA. Genome-wide screen identifies host colonization determinants in a bacterial gut symbiont. P Natl Acad Sci USA. 2016;113(48):13887–13892. doi:10.1073/pnas.1610856113.
- Caballero-Flores G, Pickard JM, Fukuda S, Inohara N, Núñez G. An enteric pathogen subverts colonization resistance by evading competition for amino acids in the gut. Cell Host Microbe. 2020;28(4):526±. doi:10.1016/j.chom.2020.06.018.
- Wang H, Xing XL, Wang JP, Pang B, Liu M, Larios-Valencia J, Liu T, Liu G, Xie S, Hao G, et al. Hypermutation-induced in vivo oxidative stress resistance enhances Vibrio cholerae host adaptation. PLOS Pathog. 2018;14(48):e1007413. doi:10.1371/journal.ppat.1007413.
- Stacy A, Andrade-Oliveira V, McCulloch JA, Hild B, Oh JH, Perez-Chaparro PJ, Sim CK, Lim AI, Link VM, Enamorado M, et al. Infection trains the host for microbiota-enhanced resistance to pathogens. Cell. 2021;184(3):615–627.e17. doi:10.1016/j.cell.2020.12.011.
- Boyd ES, Thomas KM, Dai YY, Boyd JM, Outten FW. Interplay between oxygen and Fe-S cluster biogenesis: insights from the Suf Pathway. Biochemistry. 2014;53(37):5834–5847. doi:10.1021/bi500488r.
- Przybyla-Toscano J, Roland M, Gaymard F, Couturier J, Rouhier N. Roles and maturation of iron-sulfur proteins in plastids. J Biol Inorg Chem. 2018;23(4):545–566. doi:10.1007/s00775-018-1532-1.
- Rouault TA. The indispensable role of mammalian iron sulfur proteins in function and regulation of multiple diverse metabolic pathways. Biometals. 2019;32(3):343–353. doi:10.1007/s10534-019-00191-7.
- Ayala-Castro C, Saini A, Outten FW. Fe-S cluster assembly pathways in bacteria. Microbiol Mol Biol R. 2008;72(1):110–125. doi:10.1128/mmbr.00034-07.
- Riboldi GP, Verli H, Frazzon J. Structural studies of the Enterococcus faecalis SufU [Fe-S] cluster protein. BMC Biochem. 2009;10(1):10. doi:10.1186/1471-2091-10-3.
- Riboldi GP, Larson TJ, Frazzon J. Enterococcus faecalis sufCDSUB complements Escherichia coli sufABCDSE. FEMS Microbiol Lett. 2011;320(1):15–24. doi:10.1111/j.1574-6968.2011.02284.x.
- Riboldi GP, de Oliveira JS, Frazzon J. Enterococcus faecalis SufU scaffold protein enhances SufS desulfurase activity by acquiring sulfur from its cysteine-153. Bba-Proteins Proteom. 2011;1814(12):1910–1918. doi:10.1016/j.bbapap.2011.06.016.
- Zhang XL, de Maat V, Prieto AMG, Prajsnar TK, Bayjanov JR, de Been M, Rogers MRC, Bonten MJM, Mesnage S, Willems RJL, et al. RNA-seq and Tn-seq reveal fitness determinants of vancomycin-resistant Enterococcus faecium during growth in human serum. BMC Genomics. 2017;18(1):18. doi:10.1186/s12864-017-4299-9.
- Zhang XL, Paganelli FL, Bierschenk D, Kuipers A, Bonten MJM, Willems RJL, van Schaik W, Hughes D. Genome-wide identification of ampicillin resistance determinants in Enterococcus faecium. PLOS Genet. 2012;8(6):e1002804. doi:10.1371/journal.pgen.1002804.
- Liu XY, Liu GY, Wu YS, Pang X, Wu YQ, Niu J, Chen Q, Zhang X. Transposon sequencing: A powerful tool for the functional genomic study of food-borne pathogens. Trends Food Sci Tech. 2021;118:679–687. doi:10.1016/j.tifs.2021.06.032.
- Zhang XL, Rogers M, Bierschenk D, Bonten MJM, Willems RJL, van Schaik W, Manganelli R. A lacI-family regulator activates maltodextrin metabolism of Enterococcus faecium. PLOS ONE. 2013;8(8):e72285. doi:10.1371/journal.pone.0072285.
- Wu YS, Pang XX, Liu XY, Wu YJ, Zhang XL, Cocolin L. Functional genomics identified novel genes involved in growth at low temperatures in Listeria monocytogenes. Microbiol Spectr. 2022;10(4). doi:10.1128/spectrum.00710-22.
- Pang XX, Wu YS, Liu XY, Wu YJ, Shu Q, Niu JR, Chen QH, Zhang XL, Power P. The lipoteichoic acid-related proteins YqgS and LafA contribute to the resistance of listeria monocytogenes to nisin. Microbiol Spectr. 2022;10(1). doi:10.1128/spectrum.02095-21.
- Chen SF, Zhou YQ, Chen YR, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34(17):884–890. doi:10.1093/bioinformatics/bty560.
- Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9(4):357–U54. doi:10.1038/NMETH.1923.
- Kayala MA, Baldi P. Cyber-T web server: differential analysis of high-throughput data. Nucleic Acids Res. 2012;40(W1). doi:10.1093/nar/gks420.
- Bryant P, Pozzati G, Elofsson A. Improved prediction of protein-protein interactions using AlphaFold2. Nat Commun. 2022;13(1). doi:10.1038/s41467-022-28865-w.
- Zhang Y, Skolnick J. TM-align: a protein structure alignment algorithm based on the TM-score. Nucleic Acids Res. 2005;33(7):2302–2309. doi:10.1093/nar/gki524. Print 2005.
- Hung CL, Lin YS, Lin CY, Chung YC, Chung YF. CUDA ClustalW: An efficient parallel algorithm for progressive multiple sequence alignment on Multi-GPUs. Comput Biol Chem. 2015;58:62–68. doi:10.1016/j.compbiolchem.2015.05.004.
- Singh R, Gurao A, Rajesh C, Mishra S, Rani S, Behl A, Kumar V, Kataria RS. Comparative modeling and mutual docking of structurally uncharacterized heat shock protein 70 and heat shock factor-1 proteins in water buffalo. Vet World. 2019;12(12):2036–2045. doi:10.14202/vetworld.2019.2036-2045.
- Rice L. Antibiotics and gastrointestinal colonization by vancomycin-resistant enterococci. Eur J Clin Microbiol Infect Dis. 2005;24(12):804–814. doi:10.1007/s10096-005-0057-z.
- Zhang XL, Vrijenhoek JEP, Bonten MJM, Willems RJL, van Schaik W. A genetic element present on megaplasmids allows Enterococcus faecium to use raffinose as carbon source. Environ Microbiol. 2011;13(2):518–528. doi:10.1111/j.1462-2920.2010.02355.x.
- Zhang XL, Top J, de Been M, Bierschenk D, Rogers M, Leendertse M, Bonten MJM, van der Poll T, Willems RJL, van Schaik W. Identification of a genetic determinant in clinical Enterococcus faecium strains that contributes to intestinal colonization during antibiotic treatment. J Infect Dis. 2013;207(11):1780–1786. doi:10.1093/infdis/jit076.
- Liu XY, Pang XX, Wu YS, Wu YJ, Xu LA, Chen QH, Niu J, Zhang X. New insights into the lactic acid resistance determinants of listeria monocytogenes based on transposon sequencing and transcriptome sequencing analyses. Microbiol Spectr. 2023;11(1). doi:10.1128/spectrum.02750-22.
- Krueger WA, Krueger-Rameck S, Koch S, Carey V, Pier GB, Huebner J. Assessment of the role of antibiotics and enterococcal virulence factors in a mouse model of extraintestinal translocation. Crit Care Med. 2004;32(2):467–471. doi:10.1097/01.CCM.0000109447.04893.48.
- Kuss SK, Best GT, Etheredge CA, Pruijssers AJ, Frierson JM, Hooper LV, Dermody TS, Pfeiffer JK. Intestinal microbiota promotes enteric Virus replication and systemic pathogenesis. Science. 2011;334(6053):249–252. doi:10.1126/science.1211057.
- Blahut M, Sanchez E, Fisher CE, Outten FW. Fe-S cluster biogenesis by the bacterial Suf pathway. Bba-Mol Cell Res. 2020;1867(11):118829. doi:10.1016/j.bbamcr.2020.118829.
- Vey JL, Yang J, Li M, Broderick WE, Broderick JB, Drennan CL. Structural basis for glycyl radical formation by pyruvate formate-lyase activating enzyme. P Natl A Sci India B. 2008;105(42):16137–16141. doi:10.1073/pnas.0806640105.
- Py B, Barras F. Building Fe-S proteins: bacterial strategies. Nat Rev Microbiol. 2010;8(6):436–446. doi:10.1038/nrmicro2356.
- Pérard J, de Choudens SO. Iron-sulfur clusters biogenesis by the SUF machinery: close to the molecular mechanism understanding. J Biol Inorg Chem. 2018;23(4):581–596. doi:10.1007/s00775-017-1527-3.
- Andrew AJ, Song J-Y, Schilke B, Craig EA, Fox TD. Posttranslational regulation of the scaffold for Fe-S cluster biogenesis, Isu. Mol Biol Cell. 2008;19(12):5259–5266. doi:10.1091/mbc.e08-06-0622.
- Blanc B, Gerez C, de Choudens SO. Assembly of Fe/S proteins in bacterial systems: biochemistry of the bacterial ISC system. Bba-Mol Cell Res. 2015;1853(6):1436–1447. doi:10.1016/j.bbamcr.2014.12.009.
- Garcia PS, D’Angelo F, de Choudens SO, Dussouchaud M, Bouveret E, Gribaldo S, Barras F. An early origin of iron-sulfur cluster biosynthesis machineries before Earth oxygenation. Nat Ecol Evol. 2022;6(10):1564–1572. doi:10.1038/s41559-022-01857-1.
- Yokoyama N, Nonaka C, Ohashi Y, Shioda M, Terahata T, Chen W, Sakamoto K, Maruyama C, Saito T, Yuda E, et al. Distinct roles for U-type proteins in iron–sulfur cluster biosynthesis revealed by genetic analysis of the Bacillus subtilis sufCDSUB operon. Mol Microbiol. 2018;107(6):688–703. doi:10.1111/mmi.13907.
- Albrecht AG, Netz DJA, Miethke M, Pierik AJ, Burghaus O, Peuckert F, Lill R, Marahiel MA. SufU is an essential iron-sulfur cluster scaffold protein in Bacillus subtilis. J Bacteriol. 2010;192(6):1643–1651. doi:10.1128/jb.01536-09.
- Roberts CA, Al-Tameemi HM, Mashruwala AA, Rosario-Cruz Z, Chauhan U, Sause WE, Torres VJ, Belden WJ, Boyd JM. The Suf Iron-Sulfur cluster biosynthetic system is essential in staphylococcus aureus, and decreased suf function results in global metabolic defects and reduced survival in human neutrophils. Infect Immun. 2017;85(6). doi:10.1128/iai.00100-17.
- Richardson DJ. Bacterial respiration: a flexible process for a changing environment 1999 Fleming Lecture (Delivered at the 144th meeting of the Society for General Microbiology, 8 September 1999). Microbiology. 2000;146(3):551–571. doi:10.1099/00221287-146-3-551.
- Tang YW. Molecular medical microbiology: chapter 11 - bacterial energy metabolism. Diagn Microbiol Infect Dis. 2002;43(2):173–174. doi:10.1016/S0732-8893(02)00377-2.
- Shisler KA, Hutcheson RU, Horitani M, Duschene KS, Crain AV, Byer AS, Shepard EM, Rasmussen A, Yang J, Broderick WE, et al. Monovalent cation activation of the radical SAM enzyme pyruvate formate-lyase activating enzyme. J Am Chem Soc. 2017;139(34):11803–11813. doi:10.1021/jacs.7b04883.
- Boyington JC, Gladyshev VN, Khangulov SV, Stadtman TC, Sun PD. Crystal structure of formate dehydrogenase H: catalysis involving Mo, molybdopterin, selenocysteine, and an Fe4S4 cluster. Science. 1997;275(5304):1305–1308. doi:10.1126/science.275.5304.1305.