
ABSTRACT
Histones are evolutionarily conserved proteins that together with DNA constitute eukaryotic chromatin in a defined stoichiometry. Core histones are dynamic scaffolding proteins that undergo a myriad of post-translational modifications, which selectively engage chromosome condensation, replication, transcription and DNA damage repair. Cullin4-RING ubiquitin E3 ligases are known to hold pivotal roles in a wide spectrum of chromatin biology ranging from chromatin remodeling and transcriptional repression, to sensing of cytotoxic DNA lesions. Our recent work uncovers an unexpected function of a CRL4 ligase upstream of these processes in promoting histone biogenesis. The CRL4WDR23 ligase directly controls the activity of the stem-loop binding protein (SLBP), which orchestrates elemental steps of canonical histone transcript metabolism. We demonstrate that non-proteolytic ubiquitination of SLBP ensures sufficient histone reservoirs during DNA replication and is vital for genome integrity and cellular fitness.
Introduction
Canonical histones are the core building blocks of eukaryotic chromosomes and de novo histone synthesis is essential for chromosome packaging. A surplus of histones is cytotoxic, and thus their expression is temporally and quantitatively restricted according to the length and rate of DNA replication.Citation33 In metazoans, histone transcripts originate from “replication-dependent” histone genes, that are usually clustered at one or multiple genomic loci.Citation22 Clustering may allow for a more precise transcriptional regulation, securing “en-masse” histone production exclusively during S-phase of the cell cycle.
Nascent histone transcripts lack poly(A)-tails but instead contain a highly conserved stem-loop structure within their 3′UTR that undergoes a series of unique modification steps primarily controlled by SLBP (for stem-loop-binding protein) (). SLBP recognizes and physically associates with this conserved RNA-element and remains tightly bound throughout mRNA cleavage and export to the cytoplasm, where it is eventually required for efficient translation of the mature transcript (Citation32 and references therein). 3’-end cleavage demands the recruitment of a ternary “core cleavage complex” (CCC) formed by the proteins Symplekin, CPSF100 and CPSF73. While the CCC participates in co-transcriptional 3’-end processing of all pre-mRNAs, its specific role during histone mRNA cleavage is dependent on the N-terminal region of Symplekin.Citation34 Adjacent to the cleavage site, the histone downstream element (HDE) is bound by the U7 snRNP via base pairing with the 5′-end of the U7 snRNA.Citation5,6,42,49 The U7 snRNP shares several components with spliceosomal snRNPs; however, the proteins Lsm10, Lsm11, and ZFP100, an extended protein that physically links the U7 snRNP to SLBP, are unique to this complex. Post-cleavage, SLBP-bound mature histone transcripts are exported into the cytoplasm, where SLBP again recruits several co-factors accelerating translation.Citation45 Histones fresh off the production line are then taken on by their chaperones coordinating DNA replication–coupled nucleosome assembly.
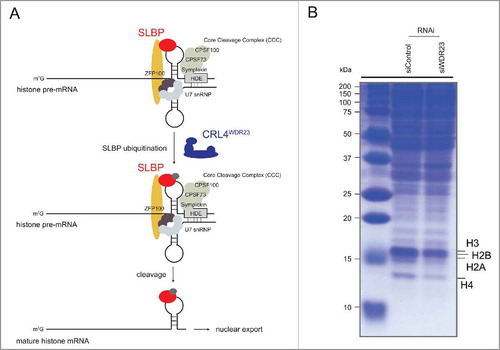
Figure 1. The CRL4WDR23 ligase activates histone mRNA processing by monoubiquitinating SLBP. (A) Schematic illustration of the nuclear histone mRNA processing pathway. SLBP binds the stem loop (SL) structure on the nascent transcript. Binding of SLBP to the hairpin is required for the subsequent recruitment of the ternary Core Cleavage Complex (CCC), which is composed of the scaffolding protein Symplekin and the endonucleases CPSF100/CPSF73. Together with the U7 snRNP complex, which basepairs with the Histone Downstream Element (HDE) and connects to SLBP via the Zn finger protein ZFP100, the CCC forms the heart of the histone pre-mRNA processing machinery. The endonucleolytic activity is further increased by additional co-factors, driving maturation of histone transcripts and subsequent SLBP-dependent export into the cytoplasm. The CRL4WDR23 E3 ligase monoubiquitinates SLBP within its RNA binding domain (RBD), and this regulation is critical for the efficiency of the histone mRNA cleavage process but not for nuclear export. (B) Downregulation of the CRL4 adaptor protein WDR23 by siRNA causes a drop in canonical histone levels due to a defect in histone transcript maturation. Extracts from U2OS cells treated with control or WDR23 siRNA were analyzed by SDS-PAGE. Total protein levels were visualized by Coomassie staining.
The cell division apparatus, and in particular timing and efficiency of DNA replication, DNA repair mechanisms as well as mitotic checkpoint control, rely on intricate cross-talk between numerous E3 ubiquitin ligases controlling the fate and/or half-life of their substrates.Citation47 Cullin4-RING E3 ubiquitin ligases (CRL4s) are critical genome caretakers, exerting prominent roles during DNA replication and repair pathways.Citation27,41 The remarkable pleiotropy of CRL4s can be explained by the modular design of these E3 ligases, which is shared among all CRL family members. Each Cullin (1, 2, 3, 4A, 4B and 5) targets multiple substrates by assembling into functional ligases with one of many interchangeable substrate recognition modules. For example, greater than 50 predicted substrate recognition proteins (DCAFs) are associated with the Cul4A/B-Rbx1-Ddb1 module comprising CRL4s, while roughly 70 substrate recognition (F-box) proteins join with Cul1-Rbx1-Skp1 to assemble SCF complexes.Citation31,38 Together, CRLs form roughly 200 distinct ubiquitin ligases, thus accounting for up to one third of the predicted human E3 pool.Citation20,28 The impact of CRL4s on cellular fitness has recently been highlighted in a human genome-wide CRISPR/Cas9 knockout screen in which the constitutive CRL4 component Ddb1 scored as an essential gene.Citation21 At present only a handful of CRL4s have been mechanistically and structurally analyzed (see below for more details on the function of selected complexes), and thus the bulk of forecasted DCAFs are still “orphan” receptors that await substrate identification.
Here we will highlight our recent findings showing that the CRL4WDR23 ligase regulates key steps of histone mRNA processing by targeting its master regulator SLBP. We speculate about the molecular consequences resulting from SLBP ubiquitination for the assembly and/or processivity of the cleavage complex. Moreover, we summarize available evidence that SLBP is targeted for proteasomal degradation in G2 phase of the cell cycle by another CRL, SCFcyclin F, thereby restricting SLBP function to S-phase. Finally, we briefly review the manifold roles of CRL4 in chromatin biology, in particular during DNA replication and repair.
A CRL4 ligase promotes histone mRNA processing by regulating SLBP
WDR23 was previously identified as a Ddb1-and-Cul4-associated factor (DCAF) that upon depletion provokes an S-phase delay accompanied by an increased frequency of DNA breaks.Citation37,39 Together, these results inferred a functional role of the protein during either DNA replication or repair. Consistent with these findings, WDR23 indeed assembles into an active, stoichiometric CRL4 complex in vitro and cellular phenotypes resulting from WDR23 depletion are caused by the lack of this particular E3 ligase activity.Citation7 Upon close examination, cells deprived of CRL4WDR23 typically presented with lagging replication forks along with an activated replication stress (intra-S) checkpoint. Limited stocks of nucleotides, histones, histone chaperones, and components of the replication machinery all adversely affect replication fork speed.Citation52 While CRL4WDR23 depletion has no obvious impact on nucleotides,Citation7 cells do gradually lose their canonical histones as a consequence of a shortage of mature histone transcripts ().
This phenotype is reminiscent of activated DNA damage pathways, which both trigger a reduction in the steady-state levels of histone mRNA by effectively repressing the transcription of core histonesCitation44 and activate the histone mRNA decay machinery to counteract cytotoxicity.Citation23,32 Likewise, ectopic stabilization of the CRL4CDT2 target Set8b, a histone methyltransferase, results in an analogous drop in histone levels due to suspended histone mRNA expression.Citation3 In the case of CRL4WDR23 however, initial transcription of histone mRNA is unaffected. On the contrary, unprocessed histone transcripts accumulate upon WDR23 depletion, pointing toward a specific block or slowdown of histone mRNA 3’-end processing in the absence of this specific E3 ligase. This hypothesis was confirmed by ablating the histone mRNA-processing factor SLBP, which binds and audits nascent histone transcripts before their cleavage until their translation in the cytoplasm. Remarkably, lack of SLBP in cells phenocopies hallmarks of WDR23 depletion, including a decline in mature histone mRNA and protein levels, a slowdown of DNA replication as well as increased checkpoint activity with none of these being further aggravated upon simultaneous depletion of both factors. Together, these findings imply a tight functional relationship between SLBP and CRL4WDR23 and raise the question of whether SLBP might be a candidate substrate for this particular ubiquitin ligase.
SLBP was previously found to be ubiquitinated and degraded right after DNA replication by an unknown E3 ligaseCitation24,56 (see also below for more details on SLBP degradation). Notably, a recent report suggested that CRL4WDR23 may also target SLBP for proteasomal degradation.Citation11 Our results, however, demonstrate that CRL4WDR23 directly governs SLBP function rather than protein levels. First, there is no evidence that cells lacking WDR23 stockpile excess SLBP. Moreover, the primary phenotype of WDR23 depletion lies in the accumulation of unprocessed histone mRNA transcripts, emphasizing that the ligase acts upstream of SLBP proteolysis. In line with this, reconstituted CRL4WDR23 complexes rapidly monoubiquitinate recombinant and cell-extract-derived SLBP. Furthermore, mass-spectrometry based ubiquitin-remnant-profiling in cells revealed that SLBP was modified at multiple lysine residues, and at least one of them is regulated by WDR23 in vivo. The functional relevance of CRL4WDR23-mediated monoubiquitination of SLBP was further underscored by the expression of a triple SLBPKR mutant that mimics the depletion phenotypes observed for WDR23 and SLBP with respect to impaired histone mRNA processing.Citation7 Collectively, these data identify SLBP, the central regulator of histone mRNA metabolism, as a novel substrate of CRL4WDR23. Monoubiquitination of SLBP is critical for the conversion from premature to translation-competent histone transcripts. If defective, unprocessed histone mRNA accumulates and histone protein levels drop, ultimately leading to a roadblock for DNA replication and elevated DNA damage.
Mechanistic consequences of non-proteolytic SLBP monoubiquitination
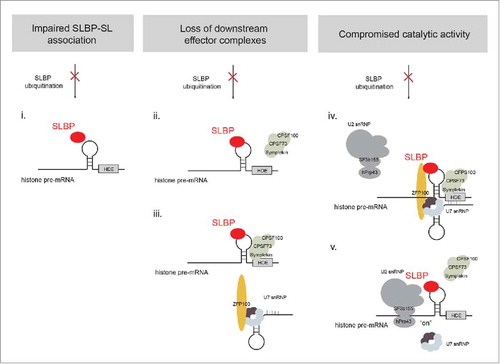
Substrate monoubiquitination typically results in functional consequences for the modified protein such as modulating its intra- or intermolecular architecture, interaction partner, localization or activity.Citation51 While our work established SLBP as a bona fide substrate of CRL4WDR23 required for core histone expression during S-phase, the underlying mechanism of when and how ubiquitination is required to activate histone pre-mRNA processing remains elusive. The first step in the pathway involves the recruitment of SLBP itself to the characteristic stem-loop (SL) within the 3’-end of histone transcripts. Considering the severe nature of WDR23 depletion on histone homeostasis it is conceivable that monoubiquitination of SLBP in cells is critical to initially liberate and/or significantly enhance its RNA-binding properties (). Indeed, this step is subject to post-translational regulation as demonstrated in Drosophila, where C-terminal phosphorylation of SLBP significantly increases its affinity for the SL.Citation54 Likewise, phosphorylation of T171 within the RNA-binding domain (RBP) of hSLBP stabilizes the RNA:protein complex.Citation12,13,55 Since CRL4WDR23 in vivo targets the exposed lysine residue K156 right in the center of the RBD,Citation7 ubiquitin moieties possibly configure SLBP to enhance SL-association. While the single non-ubiquitinatable K156R point mutant partially rescued the siRNA-depletion phenotype of SLBP, a triple mutant, which in addition to K156R harbors arginine substitutions at the neighboring lysine residues (K146R, K170R), is significantly impaired in histone mRNA processing. However, the precise number of CRL4WDR23-dependent ubiquitin sites within SLBP and their relative contribution to SL-binding remain to be determined. This becomes particularly important in light of the available crystal structure of phosphorylated SLBP bound to the SL, in which the positively charged K146 along with phosphorylated T171 and other residues contribute to the stabilization of the complex by bringing together 3 α-helices and a loop of the RBD.Citation54 Although the SLBP-K146R mutant used in our study preserves its positive charge and full RNA-binding activity,Citation12 the accessibility of this site in vivo clearly requires experimental validation. Notably, in vitro, cell extract-derived SLBP-3KR mutant protein is still a substrate for recombinant CRL4WDR23, suggesting that there are additional ubiquitin acceptor sites within SLBP (unpublished results). Nevertheless, it is unlikely that SLBP ubiquitination merely regulates RNA-binding, as unmodified hSLBP has a basal ability to bind the SL in the co-crystal structure.Citation46 Interestingly, abrogation of WDR23 function in cells results in subcellular redistribution of SLBP from the nucleus to the cytoplasm indicating that SLBP is exported along with the unprocessed histone transcripts.Citation7 This is consistent with previous observations indicating that in contrast to the SLBP-SL interaction, histone pre-mRNA metabolism per se contributes little to the consecutive export step.Citation15,45 Alternative to RNA-association, lack of SLBP ubiquitination might compromise the catalytic activity of the endonucleolytic cleavage machinery either due to the poor recruitment of the downstream effector complexes or forestalled catalysis. To probe the processing efficiency, several assays were developed in the past that allow monitoring the maturation of labeled histone pre-mRNA substrates in nuclear extracts depleted for SLBP. Restoration of pathway activity can then be probed using different versions of recombinant SLBP. In such an experimental setup our current model would make a strong prediction: if cell extracts lacking endogenous SLBP are supplemented with recombinant pre-ubiquitinated SLBP, substrate cleavage should bypass the requirement for the adaptor protein WDR23. Furthermore, to disentangle defects in downstream effector complex assembly versus enzymatic activity, immunoprecipitated S-phase SLBP- and non-ubiquitinatable SLBPKR mutant complexes might reveal the relative presence of individual CCC and U7 snRNP components (). In wild-type SLBP complexes, we readily detected the U2 snRNP spliceosome components SF3b155 along with hPrp43/DHX15, a DEAH-box helicase.Citation7 Both proteins have been previously reported to bind to an RNA element upstream of the SL, suggesting that intronless histone transcripts borrow parts of the spliceosome machinery to boost U7-snRNP-dependent catalysis.Citation17 Finally, parallels between histone mRNA cleavage and general pre-mRNA splicing allow for venturing into a more mechanistic model of how (reversible) ubiquitination might drive histone metabolism (). Spliceosome dynamics integrate a series of inputs derived from non-proteolytic ubiquitination events that trigger subunit assembly, reordering, disassembly as well as liberation of various catalytic cores.Citation8 Thus, it is tempting to speculate that in the final step of SLBP-initiated complex assembly, an ubiquitination event creates or exposes a binding-site for U2 snRNP to fuel U7-snRNP-dependent cleavage. This is consistent with our observation that immature histone transcripts accumulate upon WDR23 depletion. Alternatively, SLBP ubiquitination obstructs precocious hPrp43 activity, which is believed to release the majority of protein-RNA interactions after cleavage.Citation17 In this scenario, a sudden imbalance in complex association and dissociation rates would likewise result in a poor histone mRNA maturation process.
Figure 2. Different models of how loss of SLBP monoubiquitination by CRL4WDR23 may lower histone mRNA processing activity. Given that CRL4WDR23 targets the RBD of SLBP, it is conceivable that ubiquitination positively influences SL binding by lowering the dissociation rate of SLBP for the SL (i.). Although SLBP has an intrinsic affinity for the SL, reinforcement of this interaction might stabilize SLBP-mRNA association until the histone transcripts are translated in the cytoplasm. Alternatively, exposed ubiquitin moieties on SLBP might create binding sites for the components of the endonucleotytic machinery, in particular the CCC (ii.) and/or U7 snRNP (iii.). In each case, poor recruitment of one or both complexes would cause a severe block to mRNA cleavage. It is also conceivable that the lack of SLBP monoubiquitination leaves the initial assembly of the endonucleolytic machinery unperturbed, but has effects on its processivity. In cells, the cleavage of histone pre-mRNA is greatly enhanced by the recruitment of the spliceosomal U2 snRNP complex upstream of the SL. Ubiquitinated SLBP might thus contribute to this step, either by exposing an additional binding site for U2 snRNP or by together configuring an overall architecture that favors CCC/U7 snRNP-dependent histone mRNA cleavage (iv.). Lastly, the U2 snRNP complex contains the DEAH box helicase hPrp43, and its RNA helicase activity was previously suggested to facilitate the disassembly of the cleavage machinery before the export of the mature histone transcript. Ubiquitinated SLBP might block the catalytic activity of hPrp43 to allow completion of the cleavage reaction. Hence, a defect in SLBP ubiquitination would lead to constitutive hPrp43 helicase activity and the premature release of individual subcomplexes from the mRNA (v.).
Clearly, additional experimentation will be required, ideally using a fully reconstituted system and single-molecule analysis, to obtain a comprehensive view of how CRL4WDR23-mediated SLBP monoubiquitination activates downstream histone mRNA processing events either by enhancing the affinity of SLBP for the histone pre-mRNA stem-loop, gathering crucial downstream factors, or by tweaking the enzymatic activity of these effector proteins. Moreover, it would be interesting to test whether WDR23-mediated monoubiquitination on SLBP persists, or if a deubiquitination step is critical to facilitate release of complex components before the export of the mature transcript. If so, is there a specific deubiquitinase (DUB) involved? Alternatively, removal of ubiquitin from SLBP may liberate the protein itself from translation-competent histone transcripts, thereby closing the regulatory circuit of histone mRNA processing by decommissioning SLBP.
Coordinated histone biogenesis is likely mediated by cross-regulation of SLBP by multiple CRL complexes
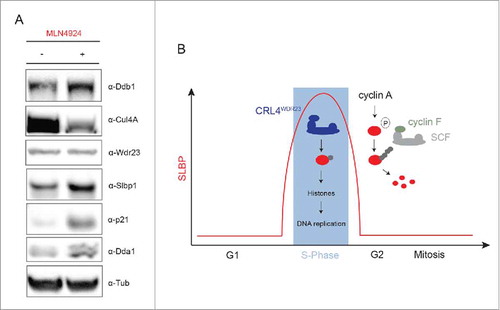
As described earlier, the rate and completion of DNA synthesis is strictly dependent on sufficient stocks of canonical histones during S-phase. Thus, not surprisingly, swift histone production is synchronized with a drastic increase in SLBP protein levels during late G1/S-phase. Shortly after completion of DNA replication, SLBP is degraded during early G2 phase in mammalian cells (). Two previous studies mapped SLBP phospho-sites that in combination trigger destruction of the protein.Citation24,56 In addition, the identification of an adjacent consensus cyclin A/F binding motif and subsequent biochemical reconstitution of SLBP phosphorylation suggested that priming of SLBP degradation requires cyclin A/Cdk1. However, until recently the relevant E3 ligase targeting SLBP for proteasomal degradation remained elusive, despite a growing body of evidence strongly pointed toward another CRL. For example, SLBP has been isolated repeatedly in proteomic screens evaluating changes in protein abundance in the presence and absence of the general CRL-activity inhibitor MLN4924.Citation14,30 Consistently, we also found SLBP stabilized upon MLN4924 treatment along with the CRL4 core components Ddb1, Dda1 and the characterized proteolytic CRL substrate p21 by western blotting, whereas we did not observe an increase in total SLBP levels upon WDR23 depletionCitation7 (). Furthermore, SLBP is stabilized upon overexpression of dominant-negative cullin1 but not cullin4,Citation14 suggesting that SLBP is a SCF substrate. Indeed, recent work identified the F-box-containing substrate adaptor cyclin F to efficiently target SLBP for degradation after S-phaseCitation10 (). In this elegant study a series of experiments were conducted to first identify and then substantiate a physical interaction between cyclin F, core SCF components and SLBP in cells. Subsequently, the authors probed the dependence of SLBP degradation in G2 on the presence and relative levels of cyclin F, and demonstrated increased in vivo ubiquitination of SLBP upon ectopic expression of the adaptor protein. Conversely, the study defined a specific SLBP “degron” mutant (RL97/99AA), which was no longer degraded in G2, in agreement with the inability of this mutant to bind cyclin F. Importantly, while impaired proteolytic destruction of SLBP had no obvious effect on core histone mRNA and protein levels, it is critical for balanced cellular H2A.X levels and genome maintenance upon genotoxic stress. Notably, proteasomal degradation of SLBP in G2 is restricted to vertebrates and evolutionary correlates with the occurrence of SCFcyclin F complexes.Citation10 As mentioned earlier, SLBP degradation in G2 was also attributed to the activity of CRL4WDR23.Citation11 The discrepancy between these 2 studies might arise from the fact that Djakbarova and colleagues used a bacterially produced short GST-SLBP fragment to isolate interaction partners in cell extracts. While they recover the CRL4 adaptor protein WDR23 as a specific binding partner, they do not find cyclin F or other SCF components. This is somewhat surprising since the SLBP truncation (aa 51–108) used in this study encompasses the minimal cyclin F binding motif (aa R97/L99). However, SLBP lacking a large proportion of its N-terminus including the NLS might not be correctly folded or too short for cyclin F binding whereas a SLBP construct spanning aa 1–99 preserves cyclin F binding.Citation10 Further, the authors demonstrate that SLBP levels increase or decline depending on the siRNA-mediated depletion or transient overexpression of WDR23.Citation11 However, given that SLBP levels fluctuate throughout the cell cycle, cell synchronization experiments in combination with inducible cell lines and siRNA resistant constructs are necessary to clarify whether WDR23 may contribute to SLBP degradation. Considering our own results and the mechanism described in Dankert et al., we currently advocate a model of sequential interaction and ubiquitination of SLBP by CRL4WDR23 and SCFcyclin F. It is also possible that CRL4WDR23 -mediated monoubiquitination during S-phase may serve as a priming step for subsequent chain formation by SCFcyclin F during G2. Intriguingly, substrate “sharing” between CRL4 and SCF complexes is an emerging conceptCitation2,50 and here used to temporally regulate the activity of SLBP through the cell cycle.
Figure 3. SLBP is targeted by the SCFcyclin Fcomplex for proteasomal degradation in G2. (A) Western blot analysis of asynchronous HeLa Kyoto cells treated with inhibitor MLN4924, which interferes with the conjugation of Nedd8 to the cullin protein. As expected, MLN4924 treatment leads to a loss of CRL activity, which is reflected in the accumulation of the CRL4 core components Ddb1 and Dda1 as well as the shared CRL4/SCF substrate p21. While downregulation of WDR23 in cells does not affect SLBP protein levels, MLN4924 treatment leads to SLBP stabilization, implying that another CRL targets SLBP for proteasomal degradation. (B) Model depicting the sequential regulation of SLBP by CRL4WDR23 and SCFcyclin F through the cell cycle.
Concluding remarks
The dynamics of chromatin, starting from its coordinated assembly to its active part in DNA replication, transcription and DNA repair, demand elaborate and synchronized regulatory mechanisms. A growing body of evidence suggests that the highly polymorphic family of CRL4 E3 ligases is critically involved in cellular DNA homeostasis by controlling the fate and lifetime of histones, chromatin remodelers, transcription factors, and repair proteins. Indeed, based on our findings CRL4WDR23 joins the ranks of several CRL4 ligases that watch over cell cycle progression and genome stability by selectively moderating chromatin-related processes in a proteolytic and regulatory fashion. While CRL4WDR23 ensures histone supply during S-phase, in S. cerevisiae CRL4 in complex with an unknown adaptor is required to convey newly synthesized H3-H4 dimers bound to the histone chaperone Asf1 to downstream chaperones for nucleosome assembly.Citation19 Likewise, CRL4RBBP7 was shown to aid timely deposition of the centromere-specific histone variant CENP-A during S-phase.Citation35 Furthermore, CRL4s at multiple steps coordinate timely cell cycle progression with DNA damage repair. Most prominently, CRL4Cdt2 manages the coordinated destruction of its targets Cdt1, p21 and Set8b upon entry into S-phase, thereby thwarting re-initiation of DNA replication. CRL4Cdt2 also participates in the UV-induced degradation of Cdt1, p21 and Set8 irrespective of the cell cycle stage.Citation1,3,4, 48 Downstream of DNA replication, CRL4DDB2 and CRL4CSA are among the first responders that track down DNA lesions.Citation16,29,43 Ddb2 ubiquitinates histones to weaken DNA-protein interactions thereby creating space for effector proteins of the global nucleotide excision repair (NER) pathway. CSA is involved in transcription-coupled NER and ubiquitinates the repair factor CSB. Other CRL4 ligases have been implicated in directly and indirectly mediating histone modification events, or in degradation of transcription factors, both altering the transcriptional landscape of a cell.Citation9,18,36,53 Additionally, CRL4s have recently received special attention as they are prone to reprogramming by exogenous factors such as drugs or viruses, thereby creating de novo ubiquitination substrates, many of which are disease-relevant transcription factors.Citation25,26,40 In summary, cullin4-RING E3 ubiquitin ligases are guardians of virtually all chromatin-relevant processes and our recent findingsCitation7 have now expanded their target range to histone production. Besides the functional characterization of the remaining CRL4 adaptor proteins, future work will increasingly aim to zoom in on non-proteolytic ubiquitination events, and mechanistically dissect the functional consequences for the target protein as well as the activity of the respective pathway.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank A. Smith and M. Soste for critical manuscript reading and helpful discussions.
Funding
Work in the Peter laboratory is funded by the ETHZ, an ERC senior award and a SNF research grant. FL is supported by the Human Frontier Science Program (HFSP; LT000376/2014).
References
- Abbas T, Dutta A. CRL4Cdt2: master coordinator of cell cycle progression and genome stability. Cell Cycle 2011; 10:241-9; PMID:21212733; http://dx.doi.org/10.4161/cc.10.2.14530
- Abbas T, Mueller AC, Shibata E, Keaton M, Rossi M, Dutta A. CRL1-FBXO11 promotes Cdt2 ubiquitylation and degradation and regulates Pr-Set7/Set8-mediated cellular migration. Mol Cell 2013; 49:1147-58; PMID:23478445; http://dx.doi.org/10.1016/j.molcel.2013.02.003
- Abbas T, Shibata E, Park J, Jha S, Karnani N, Dutta A. CRL4(Cdt2) regulates cell proliferation and histone gene expression by targeting PR-Set7/Set8 for degradation. Mol Cell 2010; 40:9-21; PMID:20932471; http://dx.doi.org/10.1016/j.molcel.2010.09.014
- Abbas T, Sivaprasad U, Terai K, Amador V, Pagano M, Dutta A. PCNA-dependent regulation of p21 ubiquitylation and degradation via the CRL4Cdt2 ubiquitin ligase complex. Gen Dev 2008; 22:2496-506; PMID:18794347; http://dx.doi.org/10.1101/gad.1676108
- Bond U, Yario TA. The steady state levels and structure of the U7 snRNP are constant during the human cell cycle: lack of cell cycle regulation of histone mRNA 3′ end formation. Cell Mol Biol Res 1994; 40:27-34; PMID:7804324
- Bond UM, Yario TA, Steitz JA. Multiple processing-defective mutations in a mammalian histone pre-mRNA are suppressed by compensatory changes in U7 RNA both in vivo and in vitro. Gen Dev 1991; 5:1709-22; PMID:1885007; http://dx.doi.org/10.1101/gad.5.9.1709
- Brodersen MM, Lampert F, Barnes CA, Soste M, Piwko W, Peter M. CRL4(WDR23)-Mediated SLBP Ubiquitylation Ensures Histone Supply during DNA Replication. Mol Cell 2016; 62:627-35; PMID:27203182; http://dx.doi.org/10.1016/j.molcel.2016.04.017
- Chen W, Moore MJ. The spliceosome: disorder and dynamics defined. Curr Opin Struct Biol 2014; 24:141-9; PMID:24530854; http://dx.doi.org/10.1016/j.sbi.2014.01.009
- Choi SH, Wright JB, Gerber SA, Cole MD. Myc protein is stabilized by suppression of a novel E3 ligase complex in cancer cells. Gen Dev 2010; 24:1236-41; PMID:20551172; http://dx.doi.org/10.1101/gad.1920310
- Dankert JF, Rona G, Clijsters L, Geter P, Skaar JR, Bermudez-Hernandez K, Sassani E, Fenyo D, Ueberheide B, Schneider R, Pagano M. Cyclin F-Mediated Degradation of SLBP Limits H2A.X Accumulation and Apoptosis upon Genotoxic Stress in G2. Mol Cell 2016; 64:507-19; PMID:27773672; http://dx.doi.org/10.1016/j.molcel.2016.09.010
- Djakbarova U, Marzluff WF, Koseoglu MM. DDB1 and CUL4 associated factor 11 (DCAF11) mediates degradation of Stem-loop binding protein at the end of S phase. Cell cycle 2016; 15:1986-96; PMID:27254819; http://dx.doi.org/10.1080/15384101.2016.1191708
- Dominski Z, Yang XC, Raska CS, Santiago C, Borchers CH, Duronio RJ, Marzluff WF. 3′ end processing of Drosophila melanogaster histone pre-mRNAs: requirement for phosphorylated Drosophila stem-loop binding protein and coevolution of the histone pre-mRNA processing system. Mol Cell Biol 2002; 22:6648-60; PMID:12192062; http://dx.doi.org/10.1128/MCB.22.18.6648-6660.2002
- Dominski Z, Zheng LX, Sanchez R, Marzluff WF. Stem-loop binding protein facilitates 3′-end formation by stabilizing U7 snRNP binding to histone pre-mRNA. Mol Cell Biol 1999; 19:3561-70; PMID:10207079; http://dx.doi.org/10.1128/MCB.19.5.3561
- Emanuele MJ, Elia AE, Xu Q, Thoma CR, Izhar L, Leng Y, Guo A, Chen YN, Rush J, Hsu PW, Yen HC, Elledge SJ. Global identification of modular cullin-RING ligase substrates. Cell 2011; 147:459-74; PMID:21963094; http://dx.doi.org/10.1016/j.cell.2011.09.019
- Erkmann JA, Sanchez R, Treichel N, Marzluff WF, Kutay U. Nuclear export of metazoan replication-dependent histone mRNAs is dependent on RNA length and is mediated by TAP. Rna 2005; 11:45-58; PMID:15611298; http://dx.doi.org/10.1261/rna.7189205
- Fischer ES, Scrima A, Bohm K, Matsumoto S, Lingaraju GM, Faty M, Yasuda T, Cavadini S, Wakasugi M, Hanaoka F, et al. The molecular basis of CRL4DDB2/CSA ubiquitin ligase architecture, targeting, and activation. Cell 2011; 147:1024-39; PMID:22118460; http://dx.doi.org/10.1016/j.cell.2011.10.035
- Friend K, Lovejoy AF, Steitz JA. U2 snRNP binds intronless histone pre-mRNAs to facilitate U7-snRNP-dependent 3′ end formation. Mol Cell 2007; 28:240-52; PMID:17964263; http://dx.doi.org/10.1016/j.molcel.2007.09.026
- Groh BS, Yan F, Smith MD, Yu Y, Chen X, Xiong Y. The antiobesity factor WDTC1 suppresses adipogenesis via the CRL4WDTC1 E3 ligase. EMBO Rep 2016; 17:638-47; PMID:27113764; http://dx.doi.org/10.15252/embr.201540500
- Han J, Zhang H, Zhang H, Wang Z, Zhou H, Zhang Z. A Cul4 E3 ubiquitin ligase regulates histone hand-off during nucleosome assembly. Cell 2013; 155:817-29; PMID:24209620; http://dx.doi.org/10.1016/j.cell.2013.10.014
- Harper JW, Bennett EJ. Proteome complexity and the forces that drive proteome imbalance. Nature 2016; 537:328-38; PMID:27629639; http://dx.doi.org/10.1038/nature19947
- Hart T, Chandrashekhar M, Aregger M, Steinhart Z, Brown KR, MacLeod G, Mis M, Zimmermann M, Fradet-Turcotte A, Sun S, et al. High-Resolution CRISPR Screens Reveal Fitness Genes and Genotype-Specific Cancer Liabilities. Cell 2015; 163:1515-26; PMID:26627737; http://dx.doi.org/10.1016/j.cell.2015.11.015
- Jaeger S, Barends S, Giege R, Eriani G, Martin F. Expression of metazoan replication-dependent histone genes. Biochimie 2005; 87:827-34; PMID:16164992; http://dx.doi.org/10.1016/j.biochi.2005.03.012
- Kaygun H, Marzluff WF. Translation termination is involved in histone mRNA degradation when DNA replication is inhibited. Mol Cell Biol 2005; 25:6879-88; PMID:16055702; http://dx.doi.org/10.1128/MCB.25.16.6879-6888.2005
- Koseoglu MM, Graves LM, Marzluff WF. Phosphorylation of threonine 61 by cyclin a/Cdk1 triggers degradation of stem-loop binding protein at the end of S phase. Mol Cell Biol 2008; 28:4469-79; PMID:18490441; http://dx.doi.org/10.1128/MCB.01416-07
- Kronke J, Hurst SN, Ebert BL. Lenalidomide induces degradation of IKZF1 and IKZF3. Oncoimmunology 2014a; 3:e941742; PMID:25610725; http://dx.doi.org/10.4161/21624011.2014.941742
- Kronke J, Udeshi ND, Narla A, Grauman P, Hurst SN, McConkey M, Svinkina T, Heckl D, Comer E, Li X, et al. Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells. Science 2014b; 343:301-5; PMID:24292625; http://dx.doi.org/10.1126/science.1244851
- Lee J, Zhou P. Pathogenic role of the CRL4 ubiquitin ligase in human disease. Front Oncol 2012; 2:21; PMID:22649780
- Lee JE, Sweredoski MJ, Graham RL, Kolawa NJ, Smith GT, Hess S, Deshaies RJ. The steady-state repertoire of human SCF ubiquitin ligase complexes does not require ongoing Nedd8 conjugation. Mol Cell Proteomics 2011; 10:M110 006460.
- Li JM, Jin J. CRL Ubiquitin Ligases and DNA Damage Response. Front Oncol 2012; 2:29; PMID:22655267
- Liao H, Liu XJ, Blank JL, Bouck DC, Bernard H, Garcia K, Lightcap ES. Quantitative proteomic analysis of cellular protein modulation upon inhibition of the NEDD8-activating enzyme by MLN4924. Mol Cell Proteomics 2011; 10:M111 009183; http://dx.doi.org/10.1074/mcp.M111.009183
- Lydeard JR, Schulman BA, Harper JW. Building and remodelling Cullin-RING E3 ubiquitin ligases. EMBO Rep 2013; 14:1050-61; PMID:24232186; http://dx.doi.org/10.1038/embor.2013.173
- Marzluff WF, Wagner EJ, Duronio RJ. Metabolism and regulation of canonical histone mRNAs: life without a poly(A) tail. Nat Rev Genet 2008; 9:843-54; PMID:18927579; http://dx.doi.org/10.1038/nrg2438
- Mejlvang J, Feng Y, Alabert C, Neelsen KJ, Jasencakova Z, Zhao X, Lees M, Sandelin A, Pasero P, Lopes M, Groth A. New histone supply regulates replication fork speed and PCNA unloading. J Cell Biol 2014; 204:29-43; PMID:24379417; http://dx.doi.org/10.1083/jcb.201305017
- Michalski D, Steiniger M. In vivo characterization of the Drosophila mRNA 3′ end processing core cleavage complex. Rna 2015; 21:1404-18; PMID:26081560; http://dx.doi.org/10.1261/rna.049551.115
- Mouysset J, Gilberto S, Meier MG, Lampert F, Belwal M, Meraldi P, Peter M. CRL4(RBBP7) is required for efficient CENP-A deposition at centromeres. J Cell Sci 2015; 128:1732-45; PMID:25795299; http://dx.doi.org/10.1242/jcs.162305
- Nakagawa T, Xiong Y. X-linked mental retardation gene CUL4B targets ubiquitylation of H3K4 methyltransferase component WDR5 and regulates neuronal gene expression. Mol Cell 2011; 43:381-91; PMID:21816345; http://dx.doi.org/10.1016/j.molcel.2011.05.033
- Olma MH, Roy M, Le Bihan T, Sumara I, Maerki S, Larsen B, Quadroni M, Peter M, Tyers M, Pintard L. An interaction network of the mammalian COP9 signalosome identifies Dda1 as a core subunit of multiple Cul4-based E3 ligases. J Cell Sci 2009; 122:1035-44; PMID:19295130; http://dx.doi.org/10.1242/jcs.043539
- Petroski MD, Deshaies RJ. Function and regulation of cullin-RING ubiquitin ligases. Nat Rev Mol Cell Biol 2005; 6:9-20; PMID:15688063; http://dx.doi.org/10.1038/nrm1547
- Piwko W, Olma MH, Held M, Bianco JN, Pedrioli PG, Hofmann K, Pasero P, Gerlich DW, Peter M. RNAi-based screening identifies the Mms22L-Nfkbil2 complex as a novel regulator of DNA replication in human cells. EMBO J 2010; 29:4210-22; PMID:21113133; http://dx.doi.org/10.1038/emboj.2010.304
- Precious B, Childs K, Fitzpatrick-Swallow V, Goodbourn S, Randall RE. Simian virus 5 V protein acts as an adaptor, linking DDB1 to STAT2, to facilitate the ubiquitination of STAT1. J Virol 2005; 79:13434-41; PMID:16227264; http://dx.doi.org/10.1128/JVI.79.21.13434-13441.2005
- Sang Y, Yan F, Ren X. The role and mechanism of CRL4 E3 ubiquitin ligase in cancer and its potential therapy implications. Oncotarget 2015; 6:42590-602; PMID:26460955
- Schaufele F, Gilmartin GM, Bannwarth W, Birnstiel ML. Compensatory mutations suggest that base-pairing with a small nuclear RNA is required to form the 3′ end of H3 messenger RNA. Nature 1986; 323:777-81; PMID:3022153; http://dx.doi.org/10.1038/323777a0
- Scrima A, Konickova R, Czyzewski BK, Kawasaki Y, Jeffrey PD, Groisman R, Nakatani Y, Iwai S, Pavletich NP, Thoma NH. Structural basis of UV DNA-damage recognition by the DDB1-DDB2 complex. Cell 2008; 135:1213-23; PMID:19109893; http://dx.doi.org/10.1016/j.cell.2008.10.045
- Su C, Gao G, Schneider S, Helt C, Weiss C, O'Reilly MA, Bohmann D, Zhao J. DNA damage induces downregulation of histone gene expression through the G1 checkpoint pathway. EMBO J 2004; 23:1133-43; PMID:14976556; http://dx.doi.org/10.1038/sj.emboj.7600120
- Sullivan KD, Mullen TE, Marzluff WF, Wagner EJ. Knockdown of SLBP results in nuclear retention of histone mRNA. Rna 2009; 15:459-72; PMID:19155325; http://dx.doi.org/10.1261/rna.1205409
- Tan D, Marzluff WF, Dominski Z, Tong L. Structure of histone mRNA stem-loop, human stem-loop binding protein, and 3′hExo ternary complex. Science 2013; 339:318-21; PMID:23329046; http://dx.doi.org/10.1126/science.1228705
- Teixeira LK, Reed SI. Ubiquitin ligases and cell cycle control. Ann Rev Biochem 2013; 82:387-414; PMID:23495935; http://dx.doi.org/10.1146/annurev-biochem-060410-105307
- Terai K, Abbas T, Jazaeri AA, Dutta A. CRL4(Cdt2) E3 ubiquitin ligase monoubiquitinates PCNA to promote translesion DNA synthesis. Mol Cell 2010; 37:143-9; PMID:20129063; http://dx.doi.org/10.1016/j.molcel.2009.12.018
- Vasserot AP, Schaufele FJ, Birnstiel ML. Conserved terminal hairpin sequences of histone mRNA precursors are not involved in duplex formation with the U7 RNA but act as a target site for a distinct processing factor. Proc Natl Acad Sci U S A 1989; 86:4345-9; PMID:2734288; http://dx.doi.org/10.1073/pnas.86.12.4345
- Wang Z, Dai X, Zhong J, Inuzuka H, Wan L, Li X, Wang L, Ye X, Sun L, Gao D, et al. SCF(beta-TRCP) promotes cell growth by targeting PR-Set7/Set8 for degradation. Nat Commun 2015; 6:10185; PMID:26666832; http://dx.doi.org/10.1038/ncomms10185
- Yau R, Rape M. The increasing complexity of the ubiquitin code. Nat Cell Biol 2016; 18:579-86; PMID:27230526; http://dx.doi.org/10.1038/ncb3358
- Zeman MK, Cimprich KA. Causes and consequences of replication stress. Nat Cell Biol 2014; 16:2-9; PMID:24366029; http://dx.doi.org/10.1038/ncb2897
- Zeng M, Ren L, Mizuno K, Nestoras K, Wang H, Tang Z, Guo L, Kong D, Hu Q, He Q, et al. CRL4(Wdr70) regulates H2B monoubiquitination and facilitates Exo1-dependent resection. Nat Commun 2016; 7:11364; PMID:27098497; http://dx.doi.org/10.1038/ncomms11364
- Zhang J, Tan D, DeRose EF, Perera L, Dominski Z, Marzluff WF, Tong L, Hall TM. Molecular mechanisms for the regulation of histone mRNA stem-loop-binding protein by phosphorylation. Proc Natl Acad Sci U S A 2014; 111:E2937-2946; PMID:25002523; http://dx.doi.org/10.1073/pnas.1406381111
- Zhang M, Lam TT, Tonelli M, Marzluff WF, Thapar R. Interaction of the histone mRNA hairpin with stem-loop binding protein (SLBP) and regulation of the SLBP-RNA complex by phosphorylation and proline isomerization. Biochemistry 2012; 51:3215-31; PMID:22439849; http://dx.doi.org/10.1021/bi2018255
- Zheng L, Dominski Z, Yang XC, Elms P, Raska CS, Borchers CH, Marzluff WF. Phosphorylation of stem-loop binding protein (SLBP) on 2 threonines triggers degradation of SLBP, the sole cell cycle-regulated factor required for regulation of histone mRNA processing, at the end of S phase. Mol Cell Biol 2003; 23:1590-601; PMID:12588979; http://dx.doi.org/10.1128/MCB.23.5.1590-1601.2003