Abstract
In 2001, we conducted a study to assess the effects of differential arrival times and nest-site selection on reproductive performance in a mixed-species heronry consisting of six species in Taeseong-ri, Chungbuk, Korea. We recorded the arrival dates, nest heights, clutch sizes, and brood sizes after 15–20 days of the age of the birds' chicks. The grey herons and cattle egrets arrived first and last, respectively, on the colony site. In the homogenous vegetation structure of the breeding site, the pitch pine trees (Pinus rigida) were mainly used for building nests on 48 of the 50 pine trees (96%). The breeding species vertically stratifies the nest sites according to their body size, except for the cattle egrets and black-crowned night herons that nested at sites higher than those predicted from their body size. The mean nest success rates of the six species under study were positively correlated with the mean nest heights. Our findings suggest that aggressive interspecific interactions among neighbors influence nest-site selection to enhance breeding success.
Introduction
Colonial breeding may benefit various avian species because the colonies serve as sites for exchanging information about threat surveillance, feeding sites, and predation risk (Krebs Citation1974; Turner and Pitcher Citation1986; Roberts Citation1996; Hafner Citation2000). Differential resource partitioning permits the birds to use resources from overlapping habitats. However, if resources such as food, shelter, and nesting sites are limited, competition develops among species as well as within species, and the cohabitation of birds may increase fitness costs (Martin Citation1996).
Nest-site competition is particularly obvious in the colonial heron species. They make nests in small areas and these nests are densely populated with hundreds or thousands of breeding birds. Many studies on this species show that nest-site competition may cause temporal and spatial segregation of the breeding colony (Custer and Osborn Citation1978; Arendt and Arendt Citation1988; Fasola and Alieri Citation1992; Si Bachir et al. Citation2008). For example, the nesting peak of cattle egrets (Bubulcus ibis) was later than that of other co-existing heron species, which avoided competition for optimal nesting sites (Maxwell and Kale Citation1977). In a homogeneous vegetation structure (consisting of a pure stand of one plant species; Burger Citation1979), the nest sites were found to be vertically stratified with mixed-species colonies of herons, with the larger species nesting higher (Burger Citation1978; Fasola and Alieri Citation1992; Naugle et al. Citation1996). Although the factors affecting nest-site choice vary among regions, the followings are considered to be the major ones: nest initiation time (Jenni Citation1969; Frederick and Collopy Citation1989), a social dominance system among neighboring birds (Parejo et al. Citation1999), nest density (Naugle et al. Citation1996; Bennetts et al. Citation2000), and environmental parameters (e.g. strong winds; Bennetts et al. Citation2000). However, the vertical stratification patterns disappear in heterogeneous (consisting of a mixed stand of plant species; Burger Citation1979) and low-height vegetation (Burger and Gochfeld Citation1990).
Because nest predation is the chief reason for nest failure in many birds, building a nest at a higher level in the vegetation has some selective advantages: (1) increased visibility of predators, (2) easy access to the nest, and (3) avoidance of contact with ground or water predators (Burger Citation1979). Thus, nest-site selection is a critical factor affecting breeding success as well as the survival of the individuals of the colonial heron species. To date, only a few studies conducted on the colonial heron species show the direct effect of nesting at a higher level on breeding success.
In this study, we examined the effects of differential arrival at the breeding site and the vertical stratification pattern of nest sites in a six mixed-species heronry. The following questions were considered. (1) Do the species arriving first at the site occupy higher nest sites in a breeding colony? (2) Do the species with a large body size occupy higher nest sites, thereby generating a vertical stratification pattern? (3) Are the breeding success rates higher in the species and individuals nesting at a higher level?
Materials and methods
Study area
The study was conducted in Taeseong-ri, Chungbuk, Korea (127°21'N, 36°35'E; ) from early March to late July 2001. The site was located on a small hill (area: approximately 625 m2; height: 63 m above sea level; 3 km southwest of the Korea National University of Education and 1.2–2.0 km west of the Meeho river), close to a residential area with approximately 30 houses. The vegetation consisted of four species: pitch pine (Pinus rigida), Japanese chestnut (Castanea crenata), black locust (Robinia pseudoacacia) and American pokeweed (Phytolacca americana), in which the pitch pine was the dominant species. The breeding colony consisted of six heron species: black-crowned night heron (Nycticorax nycticorax), cattle egret (Bubulcus ibis), great egret (Egretta alba), intermediate egret (Egretta intermedia), little egret (Egretta garzetta) and grey heron (Ardea cinerea). The site had been used as a breeding ground for the last 10 years, and increasing human disturbance had recently forced the colony to migrate from the site.
Figure 1. Map of a breeding site of the six mixed–species of herons at Taeseong, Chungbuk, Korea 2001.
Data collection
A camouflaged viewing platform (160×160×120 cm) was built on 6-m-high iron bars to reduce the disturbance caused by frequent human visitors; from this platform, the behavior of the birds was observed with binoculars or a telescope focused on the breeding colony. Weekly sampling was performed for all the nests using a pole fitted with a mirror at the top. We recorded the species, arrival times, nest heights, clutch and brood sizes after 15–20 days. Nest success rate was calculated by dividing brood size by clutch size. However, we did not count the number of young fledged aged 15–20 days at this point of time because chances were that they would leave the nests soon.
Data analysis
We compared body sizes (body length) (data obtained from Lee et al. Citation2000) with the nest heights. The collected data did not satisfy the normality assumptions of parametric statistical tests (one-sample Kolmogorov-Smirnov test, p<0.05). Therefore, we used the Kruskal-Wallis test (the nonparametric equivalent of a one-way analysis of variance (ANOVA)) and the Mann-Whitney U test (the nonparametric equivalent of an independent t-test) to test and compare the nest heights among the six heron species. Further, we used Spearman's rho correlation test to evaluate the relationship between the two measurements. Numerical data in the text are presented as mean±SD.
Results
The grey herons arrived at the breeding site in mid-March, and their number peaked by mid-April. They were followed by the great egrets, black-crowned night herons, intermediate egrets, and little egrets, whose populations peaked at the end of April (). The first flock of cattle egrets arrived in end April and their number peaked in end May. The grey herons nested at the highest level (8.0±1.27 m), followed by the black-crowned night herons, great egrets, intermediate egrets, cattle egrets, and little egrets (). There was a significant difference in the nest heights of the six heron species (Kruskal-Wallis test; χ2=34.68, p<0.001). The Mann-Whitney U test showed a significant difference in the nest heights between the cattle egrets and other species and between the grey herons and other species, except for the great egrets (). The mean nest heights did not significantly correlate with the body sizes of the six species (r s=0.543, n=6, p=0.266). The largest species occupied nests at the highest level, and the little egrets and black-crowned night herons occupied nests at heights higher than those predicted from their body sizes. The early arriving bird species occupied significantly higher nesting sites (r s=0.886, n=6, p=0.019).
Figure 2. Frequency of arrival of herons in the breeding area per two weeks at Taeseong, Chungbuk, Korea.
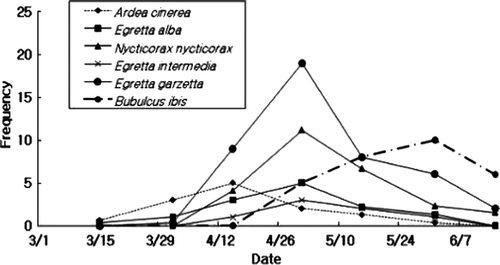
Figure 3. Box plot (median, interquartile ranges, full ranges outliers) comparison of nest heights of six herons species; LE, little egret; CE, cattle egret; IE, intermediate egret; GE, great egret; BH, black-crowned night heron; GH, grey heron.
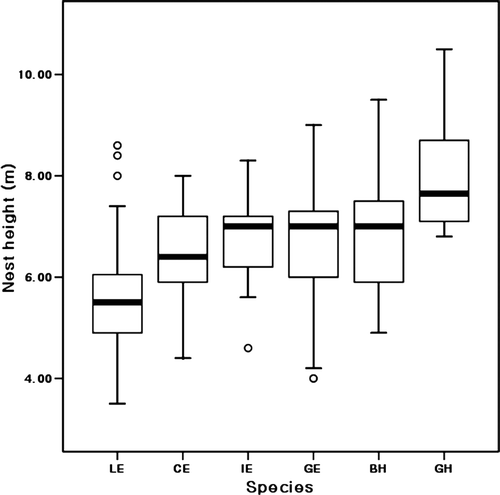
Table 1. Results of comparing nest heights in six heron species at Taeseong, Chungbuk, Korea, 2001.
The total number of active nests was 131, including 44 nests of little egrets, 29 of cattle egrets, 26 of black-crowned night herons, 13 of great egrets, 12 of grey herons, and seven of intermediate egrets. Almost all the birds built their nests on pitch pine trees (96%, 48/50 trees), and only two nests were located on the black locust trees. Thus, the vegetation structure of the colony site was homogenous.
The mean clutch sizes of little egrets and cattle egrets nesting at a lower height were higher than those of the other species, and their mean brood sizes were higher than those of great egrets and black-crowned night herons but lower than those of grey herons and intermediate egrets (). Further, the mean nest success rates of little egrets and cattle egrets were lower than those of the other species and the highest value was observed for the grey herons. Thus, the mean nest success rates of the six species were positively correlated with the mean nest heights (r s=0.829, n=6, p<0.05; ). However, in the little egret, the nest heights were negatively related to both clutch sizes (r s= − 0.308, n=44, p<0.05) and brood sizes (r s= − 0.310, n=44, p<0.05).
Figure 4. The positive relationship of mean nest success rates with mean nest heights of six species at Taeseong, Chungbak, Korea, 2001.
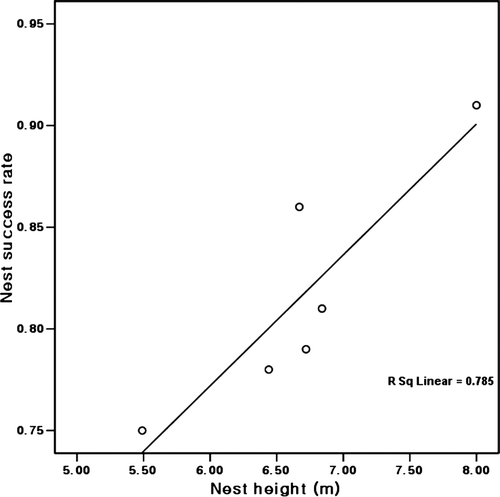
Table 2. Mean Clutch size, brood size, and nest success rate of six heron species in Taeseong, Chungbuk, Korea 2001.
Discussion
Many factors affect how the colonial heron species choose a specific nest site. Vertical stratification of the nest sites is common in homogeneous heronries with the larger birds nesting higher (Burger Citation1978). Our results are not in complete agreement with the vertical nest stratification pattern. We found that the large-size individuals of four species nested higher, except cattle egrets and black-crowned night herons, which, too, nested higher than predicted from their body sizes. Although we did not measure the degree of social interactions, judging from the aspect of nest heights, there seemed to be a direct correlation between little egrets and cattle egrets and black-crowned night herons and great egrets in competition for overlapping nest sites.
Studies on nest-site selection by little egrets and cattle egrets have shown that both species favor nesting higher in the vegetation (Burger Citation1982; Bennetts et al. Citation2000). According to the vertical nest stratification pattern reported by Burger (Citation1978), little egrets nest higher than cattle egrets in a colony of only these two species, which have similar breeding schedules (Parejo et al. Citation1999) and have existed long enough to co-evolve and nest together (Burger Citation1982).
In contrast, in our study, cattle egrets nested higher than little egrets; this was not predicted from the body lengths and the smaller number of breeding pairs in the former studies. A possible reason could be that the early-breeding cattle egrets evicted the late-breeding little egrets to lower sites in the vegetation (Hilaluddin et al. Citation2003). As the cattle egrets were the last species to arrive at the breeding sites, almost two weeks after the little egrets, they could not select higher sites from the available space in the colony without competing with the other species, especially the little egrets. However, similar findings have recently been reported by a breeder of cattle egrets in North America, where these species started breeding only from the early 1950s (Crosby Citation1972). These species have maintained unstable social relationships with their neighbors accompanied by frequent aggressive encounters. The most aggressive species, the cattle egrets, usually won the competition for securing higher nesting sites (Blaker Citation1969; Siegfried Citation1972) and their presence significantly lowered the number of nests of the other species (Burger Citation1979).
Cattle egrets are new breeders in the Korean peninsula as well; the first breeding record of this species was documented in the late 1960s, and their numbers have rapidly increased since the 1970s (Austin Citation1948; Chung Citation2004). As observed in the heronries in North America, in our study, the social interactions (e.g. aggressive interspecific interactions) between cattle egrets and little egrets may have strongly influenced nest-site selection. This factor did affect the breeding performance of the low-nesting little egrets, possibly because they were excluded from their nesting sites (Dami et al. Citation2006). Although a significant decrease was noted in the clutch and brood sizes of the little egrets, the overall nest success rates of the six species under study significantly increased with increasing nest heights. Thus, the low-nesting pairs among these six species, particularly, the little egrets may have low nest success rates. Similarly, the nest heights of the cattle egrets were positively correlated with hatching success and chick survival rates (Ranglack et al. Citation1991; Si Bachir et al. Citation2008). Hilaluddin et al. (2003) reported a higher hatching success rate in cattle egrets that nested higher than little egrets. In contrast to the cattle egrets and little egrets, the black-crowned night herons nested at sites much higher than those predicted from their body sizes. The average nest height in this species was 6.84 m, second to the grey herons (8.00 m), which nested at the highest site, and above the great egrets (6.72 m). Black-crowned night herons are also new breeders in Korea; the first breeding record was documented in 1987, and since then, the breeding distributions have expanded regionally (Park Citation2002). Until now, no information has been available to compare the factors affecting the nest heights and nest success rates between black-crowned night herons and neighboring species. In some areas where the black-crowned night herons and little egrets competed for nest sites, the early-breeding black-crowned night herons nested higher than the little egrets. The little egrets showed reduced hatching success, while the black-crowned night herons showed no variation in this rate (Kazantzidis et al. Citation1997).
In our study area, the black-crowned night herons favored the peripheral sites in the vegetation for building nests. This finding was in concordance with the peripheral nesting pattern reported by Kwon (Citation2007). Usually, the center of colonies is characterized by low predation risk and high reproductive success (Hafner Citation1980). However, if no differences are observed in the reproductive performances between the central and peripheral nests (Ranglack et al. Citation1991; Baxter Citation1994), the peripheral sites may be beneficial in that the severe competition from neighbors, especially a large colony of heterospecific species, such as the great egrets, can be avoided.
Acknowledgements
Funding was provided by the Korea National University of Education (2009 School Scholarship to Park SR).
Related Research Data
References
- Arendt , WJ and Arendt , AI. 1988 . Aspects of the breeding biology of the cattle egret (Bubulcus ibis) in Montserrat, West Indies, and its impact on nest vegetation . Colon Waterbird. , 11 : 72 – 84 .
- Austin , OL. Jr . 1948 . The birds of Korea , Cambridge, MA : Bulletin of the Museum of Comparative Zoölogy at Harvard College .
- Baxter , GS. 1994 . The influence of synchronous breeding, natal tree position, and rainfall on egret nesting success . Colon Waterbird. , 17 : 120 – 129 .
- Bennetts , RE , Fasola , M , Hafner , H and Kayser , Y. 2000 . Influence of environmental and density-dependent factors on reproduction of Little Egrets . Auk. , 117 : 634 – 639 .
- Blaker , D. 1969 . Behaviour of the cattle egret Ardeola ibis . Ostrich. , 40 : 75 – 129 .
- Burger J. 1978 . The pattern and mechanism of nesting in mixed species heronries . In : Sprunt A , Odgen JC , Wincker S , . Wading birds. Research Report No.7 . New York, NY : National Audubon Society . p. 45 58 .
- Burger , J. 1979 . Resource partitioning: Nest site selection in mixed-species colonies of herons, egrets, and ibises . Am Midl Nat. , 101 : 191 – 210 .
- Burger , J. 1982 . On the nesting location of cattle egrets Bubulcus ibis in South African heronries . Ibis. , 124 : 523 – 529 .
- Burger , J and Gochfeld , M. 1990 . Vertical nest stratification in heronry in Madagascar . Colon Waterbird. , 15 : 185 – 192 .
- Chung H. 2004 . Breeding strategies and social behaviour of Ardeidae [Ph.D. thesis] . Seoul : Kyung Hee University .
- Crosby , GT. 1972 . Spread of the cattle egret in the Western Hemisphere . Bird-Banding. , 43 : 205 – 212 .
- Custer , TW and Osborn , RG. 1978 . Feeding habitat use by colonially breeding herons, egrets, and ibises in North Carolina . Auk. , 95 : 733 – 743 .
- Dami , L , Bennetts , RE and Hafner , H. 2006 . Do Cattle Egrets exclude Little Egrets from settling at higher quality sites within mixed-species colonies? . Waterbirds. , 29 : 154 – 162 .
- Fasola , M and Alieri , R. 1992 . Nest site characteristics in relation to body size in herons in Italy . Colon Waterbird. , 15 : 185 – 191 .
- Frederick , PC and Collopy , MW. 1989 . Nesting success of five ciconiiform species in relation to water conditions in the Florida Everglades . Auk. , 106 : 625 – 634 .
- Hafner , H. 1980 . Etude ecologique des colonies des herons arboricoles (Egretta g. garzetta L., Ardeola r. ralloides Scop., Ardeola i. ibis L., Nycticorax n. nycticorax L.) en Camargue . Bonn Zool Beit. , 31 : 249 – 287 .
- Hafner , H. 2000 . “ Heron nest site conservation ” . In Heron conservation , Edited by: Kushlan , AJ and Hafner , H . 201 – 215 . New York, NY : Academic Press .
- Hilaluddin , J , Shah , N and Shwal , TA. 2003 . Nest site selection and breeding success by cattle egret and little egret in Amroha, Uttar Pradesh, India . Waterbirds. , 26 : 444 – 448 .
- Jenni , DA. 1969 . A study of the ecology of four species of herons during the breeding season at Lake Alice, Alachua County, Florida . Ecol Monogr. , 39 : 245 – 270 .
- Kazantzidis , S , Goutner , V , Pyrovetsi , M and Sinis , A. 1997 . Comparative nest site selection and breeding success in 2 sympatric ardieds, black-crowned night heron Nycticorax nycticorax, and little egret Egretta garzetta in the Axios Delta, Macedonia, Greece . Colon Waterbird. , 20 : 505 – 517 .
- Krebs , JR. 1974 . Colonial nesting and social feeding as strategies for exploiting food resources in the great blue heron (Ardea herodias) . Behaviour. , 51 : 99 – 134 .
- Kwon IK. 2007 . Nest site selection in heron species [M.Sc. thesis] . Seoul : Kyung Hee University .
- Lee , WS , Koo , TH and Park , JY. 2000 . A field guide to the birds of Korea , Seoul : LG Evergreen Foundation .
- Martin , TE. 1996 . Fitness costs of resource overlap among coexisting bird species . Nature. , 380 : 338 – 340 .
- Maxwell , GR and Kale , W. 1977 . Breeding biology of five species of herons in coastal Florida . Auk. , 94 : 689 – 700 .
- Naugle , DE , Johnson , RR , Meeks , WA and Higgins , KF. 1996 . Colonization and growth of a mixed-species heronry in South Dakota . Colon Waterbird. , 19 : 199 – 206 .
- Parejo , D , Sacchez , JM and Aviles , JM. 1999 . Factors affecting the nest height of three heron species in heronries in the southwest Spain . Ardeola. , 42 : 227 – 230 .
- Park JY. 2002 . Current status and distribution of birds in Korea [Ph.D. thesis] . Seoul : Kyung Hee University .
- Ranglack , GS , Angus , RA and Marion , KR. 1991 . Physical and temporal factors influencing breeding success of cattle egret (Bubulcus ibis) in a west Alabama colony . Colon Waterbird. , 14 : 140 – 149 .
- Roberts , G. 1996 . Why individual vigilance declines as group size increases? . Anim Behav. , 51 : 1077 – 1086 .
- Si Bachir , A , Barbraud , C , Doumandji , S and Hafner , H. 2008 . Nest site selection and breeding success in an expanding species, the cattle egret Bubulcus ibis . Ardea. , 96 : 99 – 107 .
- Siegfried , WR. 1972 . Breeding success and reproductive output of the cattle egret . Ostrich. , 43 : 43 – 55 .
- Turner , GF and Pitcher , TJ. 1986 . Attack abatement: a model for group protection by combined avoidance and dilution . Am Nat. , 128 : 228 – 240 .