
ABSTRACT
The Japanese otter (Lutra nippon), once inhabited in most islands of Japan, is now considered as an extinct species. Although the Japanese otter is regarded as a distinct species from the Eurasian otter (L. lutra), its phylogeny and taxonomic status are based on limited information on morphological and genetic data, and thus further clarification is required. Here, we assessed the phylogenetic relationship among the genus Lutra and taxonomic status of L. nippon by using the complete sequences of cytochrome b gene of its holotype. The present phylogenic trees supported that the genus Lutra specimens largely formed monophyletic group, with L. sumatrana as a basal to other Lutra species. Within Lutra species, L. nippon was distantly related with L. lutra. The European otter population of L. l. lutra were clustered together with its subspecies, L. l. chinensis rather than the same subspecies, Korean otter population. The discrepancy between the genetic data and traditional taxonomy justifies the necessity of reexamination of the current subspecific classification system of Eurasian otters. Level of genetic divergence between the holotype of L. nippon and L. lutra was two to three-fold lower than those among the other sister species of the Lutrinae. Based on the level of divergence between the L. nippon and L. lutra, and insufficient evidence of morphological difference between them, it is suggested that designation of Japanese otter as a separate species from L. lutra will be reconsidered.
Introduction
The Japanese otter (Lutra nippon) once inhabited most of the main Japanese islands except Hokkaido (Sasaki Citation1995). However, due to a rapid decline in its population, it has not been sighted since the 1980s and was officially declared extinct by the Japanese government on 29 August Citation2012 (Kyodo News). Since the 1980s, there has been increased attention to this species (Yamamoto and Ando Citation2011) and it has become the subject of several morphological and genetic studies. These studies largely concluded that L. nippon is a distinct species separate from the Eurasian otter Lutra lutra (Imaizumi and Yoshiyuki Citation1989; Suzuki et al. Citation1996; Endo et al. Citation2000). However, the taxonomic status of L. nippon still remains uncertain because the conclusion is derived from limited morphological and genetic data.
According to Wozencraft (Citation2005), the genus Lutra includes three species: the Eurasian otter, L. lutra; the hairy-nosed otter, L. sumatrana; and the Japanese otter, L. nippon. L. sumatrana and L. nippon have limited distributions in southeast Asia and the Japanese islands, respectively. On the contrary, L. lutra is widely distributed across the Eurasian continent including south of the tundra line and in north Africa, and is divided into seven subspecies (Pocock Citation1941; Roos et al. Citation2015): L. l. lutra is distributed in Eurasia from England to the Korean peninsula, excluding India, southeast Asia, and southern China; L. l. chinensis inhabits the southern part of China and Kimmen Island of Taiwan; and the remaining five subspecies (L. l. barang, L. l. nair, L. l. monticola, L. l. kutab, and L. l. aurobrunnea) are distributed in the southern part of Asia. L. nippon was traditionally identified as a subspecies of L. lutra, L. l. whiteleyi, which is synonymous with L. l. lutra, based on morphological features of its skin and skull (Gray Citation1867; Imaizumi Citation1949). Conversely, Imaizumi and Yoshiyuki (Citation1989) suggested that L. nippon was a distinct species, based on skulls from the Shikoku, Honshu, and Hokkaido areas, which were morphologically distinct from the skulls of L. lutra, including L. l. whiteleyi. Likewise, Suzuki et al. (Citation1996) found L. nippon to be a distinct species from L. lutra based on the genetic difference of 3.6% in the partial sequences of its cytochrome b gene (224 bp). The length of analyzed sequence in their study was, however, too short to define a species. Furthermore, the sample size in their study was not sufficient to compare inter- and intraspecific variation among the genus Lutra. Therefore, Roos et al. (Citation2015) concluded that the taxonomic position of L. nippon remains uncertain requiring further studies and cannot view it as a separate species from L. lutra. Regarding morphological differences, Lau et al. (Citation2016) identified that L. lutra from South Korea is sexually dimorphic. Therefore, sexual dimorphism in L. nippon is a possible confounding effect and thus interpretation should be cautioned when using morphology to evaluate their taxonomy. Waku et al. (Citation2016) analyzed the phylogenetic relationship among Lutra spp., including L. nippon, using mitochondrial genome sequences (14,740 bp) and consequently divided the Japanese populations into two lineages: L. lutra and another Lutra species or subspecies. Additionally, Waku et al. (Citation2016) identified two lineages of Eurasian otter (L. lutra) in East Asia; one lineage comprising of Chinese otter (L. l. chinensis) and another comprising of Eurasian otter (L. l. lutra) from South Korea and Sakhalin, Russia. However, limited sampling of the Eurasian otter range in East Asia led to phylogenetic relationships among East Asian otter populations an uncertain state. Koh et al. (Citation2004) concluded that partial mitochondrial DNA sequence of Korean otter was distinct from those of European otters, but authors provided limited information on relationships of Eurasian otter populations. Therefore, the phylogenetic relationship of Eurasian otters at species and subspecies level in East Asia still remains unclear and the taxonomic status of L. nippon remains controversial.
Genetic markers based on mitochondrial DNA, such as the cytochrome b gene, hypervariable portion of the control region (D-loop), and cytochrome c oxidase I, have been used for phylogenetic and population genetic analysis for most mammalian taxa. Specifically, the cytochrome b gene sequences have been used to investigate relationships among mammalian taxa at a family-subspecific level (Ledje and Arnason Citation1996; Johns and Avise Citation1998; Koepfli and Wayne Citation1998; Bradley and Baker Citation2001; Kurose et al. Citation2008; Koepfli, Deere, et al. Citation2008; Koepfli, Kanchanasaka, et al. Citation2008). Hence, the objective of this study is to determine the molecular phylogeny of L. nippon using the cytochrome b gene and to clarify its taxonomic status. Only few Japanese otter specimens have a reliable information on locality, and therefore we focused on the relationship among the holotypes of L. nippon (Imaizumi and Yoshiyuki Citation1989), L. lutra, and L. sumatrana. We also investigated the phylogenetic relationship of Eurasian otters at subspecific level.
Materials and methods
Samples examined in this study and sequence data from GenBank are summarized in . The holotype of L. nippon (Imaizumi and Yoshiyuki Citation1989) was employed in this study. This holotype was collected in 1972 from the Nenokubi seaside of Kochi Prefecture in Japan where its skeleton and mounted skin are preserved in the National Museum of Nature and Science, Tokyo, Japan.
Table 1. Sample and DNA sequence information used in this study.
Six L. l. lutra specimens from the Korean peninsula were also examined to assess the possible sequence variation among L. l. lutra individuals. Korean otter specimens were collected from several areas in South Korea and they were obtained from a variety of sources, including individuals that were road killed, caught in a fishing net or illegal trap, or that had been rescued as cubs but subsequently died. These tissue samples had been preserved in the Conservation Genome Resource Bank for Korean Wildlife (CGRB) and Association of Korean Otter Conservation (AKOC) with proper permits from the Cultural Heritage Administration (CHA) of South Korean government because Korean otter is designated as a natural monument species by the CHA. The sequence data of complete cytochrome b genes of L. l. lutra from Europe and South Korea, L. l. chinensis, L. nippon, L. sumatrana, Aonyx capensis, A. cinereus, Lontra felina, Lontra longicaudis, Lontra canadensis (all subfamily Lutrinae), and Taxidea taxus (family Mustelidae) were cited from GenBank as reference data.
Total DNA from the holotype of L. nippon was extracted from dried costal cartilage that had been preserved in the National Museum of Nature and Science, Tokyo. The cartilage was washed in 99% ethanol after treatment with a TE buffer. The cartilage pieces were cut into 0.5–1.0 cm and then decalcified with EDTA (pH 8.0) at room temperature for 5 days. DNA was extracted using an Ultra Clean™ Tissue DNA Isolation Kit (MO BIO Laboratories Inc.) following the manufacturer’s protocol. All procedure of DNA extraction from the holotype specimen was performed in the clean bench for preventing contamination.
Polymerase chain reaction (PCR) amplification of the cytochrome b gene of the holotype of L. nippon was performed in a 10-µL reaction volume containing the following reagents: 10× Ex Taq Buffer (Takara Bio Inc.), 0.5 mM of each dNTP mix (Takara Bio Inc.), 2 µM forward and reverse primers (Supplement A, B), 0.5 U Ex Taq (Takara Bio Inc.), and 1.0 µL template DNA. Amplification was conducted for a total of 46 cycles using the 14 primers designed in the present study (Supplement A, B). The conditions for the initial 10 cycles were as follows: 94°C for 30 s, 45°C for 20 s, and 72°C for 20 s; and the conditions for the remaining 36 cycles were: 94°C for 30 s, 55°C for 20 s, and 72°C for 20 s. Each of the partial cytochrome b sequences of amplicons that were amplified by a combination of 14 primers was analyzed using an IBM 3130 sequencer analyzer (Applied Biosystems™), and those sequences were aligned and assembled using Geneious Pro v5.3 to obtain the complete cytochrome b sequence (1140 bp) from the holotype of Lutra nippon.
Genomic DNA of L. l. lutra from Korea was extracted from the tissue using a DNA extraction kit (Blood & Tissue Kit, QiagenTM) according to the manufacturer’s manual. PCR amplification of the complete cytochrome b gene from the Korean population of L. l. lutra was carried out under the following conditions: one cycle of 94°C for 4 min, 35 cycles of 94°C for 30 s, 40°C for 60 s, 72°C for 90 s, and a final cycle of 72°C for 5 min. Each 30-µL reaction volume contained 10× PCR buffer (iNtRON Biotechnology, Inc.), 0.2 mM of each dNTP mix (iNtRON Biotechnology,Inc.), 0.5 µM forward (L14724: CGA AGC TTG ATA TGA AAA ACC ATC GTT G) and reverse (H15915: AAC TGC AGT CAT CTC CGG TTT ACA AGA C) primers (Collura et al. Citation1996), 1U i-StarTaq (iNtRON Biotechnology, Inc.), and 1.5 µL template DNA (30 ng). Six complete cytochrome b sequences (1140 bp) were analyzed using an ABI3730 XL sequencer analyzer (Applied Biosystems™). Sequences were aligned using Geneious Pro v5.3 (Kearse et al. Citation2012).
The pairwise genetic distance among those sequences was calculated by using PAUP4.0 based on Kimura 2 Parameter (Kimura Citation1980). Jmodeltest2.1.8 was used to find the best-fit substitution model of sequence evolution for constructing phylogenetic trees (Posada Citation2008). A maximum likelihood (ML) tree was reconstructed using PAUP4.0 (Swofford Citation2001), with an application of 1000 pseudoreplicates of this ML tree to obtain bootstrap support values. The Bayesian inference (BI) tree was obtained using MrBayes 3.2.3 (Ronquist et al. Citation2012). BI employed four simultaneous Monte Carlo Markov chains (one cold and three heated) with 1,000,000 generations and sampled every 500 generations. The first 25% of the data points were discarded as burn-in. The consensus trees from both ML and BI were illustrated using FigTree v 1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/).
Results and discussion
The phylogenic relationship among Lutrinae species was constructed using the ML and Bayesian methods under GTR + G substitution model, which was selected from Jmodeltest 2.1.8, based on their Akaike information criterion (AIC = 8952.42) value (). The phylogenic tree supported that the genus Lutra specimens largely formed a monophyletic group, with L. sumatrana as a basal to other Lutra species (, node 1). In the Chinese otter group, CH2 and CH3 were clustered with L. l. chinensis (CH1), so they were presumed to be the same subspecies, L. l. chinensis. Within Lutra species, the European otter population of L. l. lutra were clustered together with its subspecies, L. l. chinensis (or L. l. spp) from China rather than the same subspecies, Korean otter population (, node 2). The results of the molecular phylogenetic analysis in this study are not in agreement with the traditional subspecific taxonomic system of L. lutra, in which Korean otters and European otters are classified as the same subspecies (L. l. lutra), and otters in Southern part of China are regarded as a distinguished subspecies, L. l. chinensis. However, the results of this study identified haplotypes of Korean otter population as a monophyletic group distinct from the European and Chinese otter populations (, node 2) implying that Korean otters had been branched out earlier than the divergence between L. l. chinensis and European otters of L. l. lutra. The discrepancy between the genetic data and traditional taxonomy justifies the necessity of reexamination of the current subspecific classification system of Eurasian otters.
Figure 1. Phylogenetic tree reconstructed by Bayesian method using MrBayes 3.1.3, and this Bayesian tree has same topology with maximum likelihood tree (-Ln likelihood = 4427.196) reconstructed by PAUP4.0. (a) Bootstrap support values (%) obtained from 1000 pseudoreplicates of maximum likelihood tree. (b) Posterior probabilities (%) from Bayesian method.
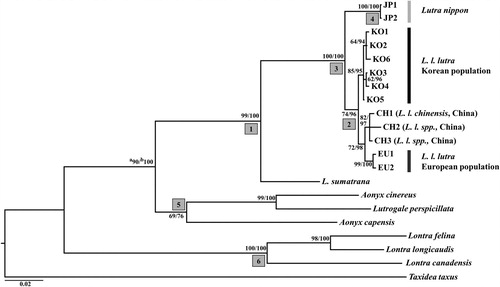
It is notable that a clade clustering the L. nippon and L. lutra specimens was strongly supported by both phylogenetic trees, with a bootstrap value of 100% in ML trees and a posterior probability of 100% in BI tree (, node 3). The phylogenetic relationship showed that the holotype of L. nippon (JP1) and another Japanese otter (JP2) identified by Waku et al. (Citation2016) were a member of the genus Lutra (, node 1, 4). Furthermore, the holotype of L. nippon formed a monophyletic group with L. lutra although it was reported that they were isolated from two other Lutra species, L. lutra and L. sumatrana (Imaizumi and Yoshiyuki Citation1989; Suzuki et al. Citation1996). The phylogenetic relationship shown in this study was consistent with that of Waku et al. (Citation2016), who used part of the 14,740 bp mitochondrial genome sequences for comparison. Waku et al. (Citation2016) identified two lineages that one belongs to L. lutra and the other is an old Japanese lineage, and thus they were regarded as either a new Lutra species or a subspecies of L. lutra. Since complete cytochrome b sequences of the old Japanese lineage (JP2) in Waku et al. (Citation2016) is identical with that of the holotype of L. nippon, the lineage is considered to represent L. nippon (. node 4). Pairwise genetic distances of the cytochrome b gene among the holotype of L. nippon, L. lutra, including L. l. lutra and L. l. chinensis, and the other seven species of Lutrinae are shown in . In spite of the small sample size, our findings based on genetic distance support the distinction of L. nippon from L. lutra. The holotype of L. nippon had genetic distances of 2.4–2.8% and 2.8–3.3% from L. l. lutra and L. l. chinensis (or L. l. spp) from China, respectively, whereas the distance between the two Lutra subspecies, L. l. lutra and L. l. chinensis, was only 0.8–1.4%. Korean and European populations of L. l. lutra, respectively located at the east and west extreme of Eurasian continent, had a genetic distance of 0.9–1.2% between them despite long geographic distance. It is thus unlikely that L. nippon is part of the variation within L. lutra. Johns and Avise (Citation1998) concluded that the genetic distance of the cytochrome b gene among mammalian sister species generally ranged from 2% to 24%. According to their index, the genetic distance between the holotype of L. nippon and L. lutra (L. l. lutra + L. l. chinensis, 2.4–3.3%) found in this study suggests that they differ at a marginal level. The genetic divergence between the holotype of L. nippon and L. lutra was two to three-fold smaller than those between the sister species in the subfamily Lutrinae (6.7–7.2% between L. lutra and L. sumatrana, 8.1–11.6% among A. capensis, A. cinereus, and Lutrogale perspicillata (, node 5), and 5.8–11% among L. felina, L. longicaudis, and L. canadensis (, node 6)). Therefore, it is suggested that the holotype of L. nippon diverged from L. lutra at a boundary point between a subspecies and a species of the genus Lutra.
Table 2. Pairwise genetic distance based on cytochrome b gene (1140 bp) sequence variance.
Despite the geographic proximity of Korea to Japan than Europe, Korean otter populations are more closely related to those of Europe than to Japanese populations. A similar pattern has been reported in other mammalian species such as the Siberian flying squirrel (Pteromys volans), the Asiatic black bear (Ursus thibetanus), and the raccoon dog (Nyctereutes procyonoides) (Lee et al. Citation2008; Kim et al. Citation2011, Citation2013). The isolation and differentiation of L. nippon population are assumed owing to the geographic isolation of the Japanese islands from the Eurasian continent by the sea. Over time, vicariance such as the geographic isolation of L. nippon from L. lutra in the Eurasian continent may have led to its speciation. Indeed, L. nippon also exhibits a certain level of morphological divergence from L. lutra. Imaizumi and Yoshiyuki (Citation1989) described L. nippon as being generally similar to L. lutra but with certain morphological differences, with, in the former species, a larger skull with a longer facial portion, a relatively small inner lobe of P4 (protocone), a longer tail, and a naked and larger rhinalium. Endo et al. (Citation2000) also outlined some obvious differences in skull shape between L. nippon and the continental L. lutra populations using multivariate analyses. Whereas, Waku et al. (Citation2016) identified two lineages of otters from Japan – L. lutra and another Lutra species or subspecies – the latter being considered as L. nippon (Imaizumi and Yoshiyuki Citation1989). If L. nippon and L. lutra had occurred sympatrically in Japan, the evidence of genetic difference between the holotype of L. nippon and L. lutra found in our study could be explained by the existence of reproductive barriers between them. However, there is no evidence of sympatry between L. nippon and L. lutra. Both the holotype of L. nippon in this study and the specimen examined by Waku et al. (Citation2016), which was considered to be the L. nippon lineage, were obtained from the Kochi Prefecture, whereas the specimen regarded as L. lutra in Waku et al. (Citation2016) was from Jogashima, Kanagawa Prefecture. Furthermore, Waku et al. (Citation2016) stated that L. lutra from Jogashima may have been brought there artificially. Further clarification of the taxonomic status of L. nippon requires the inclusion of more specimens from the Jogashima region. Although L. nippon has diverged from L. lutra at certain genetic and morphological levels, there seems insufficient evidence yet to categorize it as a distinct species.
Based on the level of divergence between the L. nippon and L. lutra, and limited evidence of morphological difference between them, it is, therefore, suggested that designation of Japanese otter as a separate species from L. lutra will be reconsidered until comprehensive and robust evidence supporting independent specific status of L. nippon are discovered. In addition, taxonomic classification of a regionally extirpated population as a separate species without concrete scientific evidence would not be desirable because it may preclude the potential restoration or reintroduction planning or discussion of the species into the historical range in the future.
Supplemental Material
Download MS Word (24.4 KB)Supplemental Material
Download MS Word (120.1 KB)Acknowledgments
We gratefully express our gratitude to all who donated Otter samples for this study: The National Museum of Nature and Science, Tokyo, Japan and Korean Otter Research Center (KORC), Hwacheon-Gun, South Korea. This study was partially supported by The Research Institute for Veterinary Science, Seoul National University.
Disclosure statement
No potential conflict of interest was reported by the authors.
Related Research Data
References
- Bradley RD, Baker RJ. 2001. A test of the genetic species concept: cytochrome-b sequences and mammals. J Mammal. 82:960–973. doi: 10.1644/1545-1542(2001)082<0960:ATOTGS>2.0.CO;2
- Collura RV, Auerbach MR, Stewart C-B. 1996. A quick, direct method that can differentiate expressed mitochondrial genes from their nuclear pseudogenes. Curr Biol. 6:1337–1339. doi: 10.1016/S0960-9822(02)70720-3
- Endo H, Ye X, Kogiku H. 2000. Osteometrical study of the Japanese otter (Lutra nippon) from Ehime and Kochi prefectures. Mem Natl Sci Mus (Tokyo). 33:195–201.
- Fernandes CA., Ginja C., Pereira I., Tenreiro R., Bruford MW., Santos-Reis M. 2008. Species-specific mitochondrial DNA markers for identification of non-invasive samples from sympatric carnivores in the Iberian Peninsula. Conserv Genet. 9:681. doi: 10.1007/s10592-007-9364-5.
- Gray JE. 1867. Notice of Lutronectes whiteleyi, an otter from Japan. P Zool Soc Lond. 35:180–182.
- Imaizumi Y. 1949. The natural history of Japanese mammals. Tokyo: Yoyo shobo. p. 348.
- Imaizumi Y, Yoshiyuki M. 1989. Taxonomic status of the Japanese otter (Carnivora, Mustelidae), with a description of a new species. Bull Natl Sci Mus Ser A Zool. 15:177–188.
- Jang KH, Ryu SH, Hwang UW. 2009. Mitochondrial genome of the Eurasian otter Lutra lutra (Mammalia, Carnivora, Mustelidae). Genes Genom. 31:19–27. doi: 10.1007/BF03191134
- Johns GC, Avise JC. 1998. A comparative summary of genetic distances in the vertebrates from the mitochondrial cytochrome b gene. Mol Biol Evol. 15:1481–1490. doi: 10.1093/oxfordjournals.molbev.a025875
- Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, et al. 2012. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 28:1647–1649. doi: 10.1093/bioinformatics/bts199
- Ki JS, Hwang DS, Park TJ, Han SH, Lee JS. 2010. A comparative analysis of the complete mitochondrial genome of the Eurasian otter Lutra lutra (Carnivora; Mustelidae). Mol Biol Rep. 37:1943–1955. doi: 10.1007/s11033-009-9641-0
- Kim YK, Hong YJ, Min MS, Kim KS, Kim YJ, Voloshina I, Myslenkov A, Smith GJ, Cuong ND, Tho HH, et al. 2011. Genetic status of Asiatic black bear (Ursus thibetanus) reintroduced into South Korea based on mitochondrial DNA and microsatellite loci analysis. J Hered. 102:165–174. doi: 10.1093/jhered/esq121
- Kim SI, Park S-K, Lee H, Oshida T, Kimura J, Kim YJ, Nguyen ST, Sashika M, Min MS. 2013. Phylogeography of Korean raccoon dogs: implications of peripheral isolation of a forest mammal in East Asia. J Zool. 290:225–235. doi: 10.1111/jzo.12031
- Kimura M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 16:111–120. doi: 10.1007/BF01731581
- Koepfli KP, Deere KA, Slater GJ, Begg C, Begg K, Grassman L, Lucherini M, Veron G, Wayne RK. 2008. Multigene phylogeny of the Mustelidae: Resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biol. 6:10. doi: 10.1186/1741-7007-6-10
- Koepfli K-P, Kanchanasaka B, Sasaki H, Jacques HL, Louie KDY, Hoai T, Dang NX, Geffen E, Gutleb A, Han S-Y, et al. 2008. Establishing the foundation for an applied molecular taxonomy of otters in Southeast Asia. Conserv Genet. 9:1589–1604. doi: 10.1007/s10592-007-9498-5
- Koepfli KP, Wayne RK. 1998. Phylogenetic relationships of otters (Carnivora: Mustelidae) based on mitochondrial cytochrome b sequences. J Zool Lond. 246:401–416. doi: 10.1111/j.1469-7998.1998.tb00172.x
- Koh HS, Yoo MH, Lee BG, Park JG. 2004. Molecular DNA systematic analyses of East Asian mammals: sequence variation of cytochrome b gene and control region of mitochondrial DNA of common otter, Lutra lutra lutra L. (Mammalia, Carnivora) from Korea. Korean J Biol Sci. 8:231–233. doi: 10.1080/12265071.2004.9647755
- Kurose N, Abramov AV, Masuda R. 2008. Molecular phylogeny and taxonomy of the genus Mustela (Mustelidae, Carnivora), inferred from mitochondrial DNA sequences: new perspectives on phylogenetic status of the back-striped weasel and American mink. Mamm Study. 33:25–33. doi: 10.3106/1348-6160(2008)33[25:MPATOT]2.0.CO;2
- Kyodo News. 2012 Aug 29. Japanese river otter declared extinct. The Japan Times. p. 1.
- Lau ACC, Asahara M, Han SY, Kimura J. 2016. Sexual dimorphism of the Eurasian otter (Lutra lutra) in South Korea: Craniodental geometric morphology. J Vet Med Sci. 78:1007–1011. doi: 10.1292/jvms.16-0018
- Ledje C, Arnason U. 1996. Phlyogenetic analyses of complete cytochrome b genes of the order Carnivora with particular emphasis on the caniformia. J Mol Evol. 42:135–144. doi: 10.1007/BF02198839
- Lee MY, Park SK, Hong YJ, Kim YJ, Voloshina I, Myslenkov A, Saveljev AP, Choi TY, Piao RJ, An JH, et al. 2008. Mitochondrial genetic diversity and phylogenetic relationships of Siberian flying squirrel (Pteromys volans) populations. Animal Cells Syst. 12:269–277. doi: 10.1080/19768354.2008.9647182
- Pocock RI. 1941. The fauna of British India including Ceylon and Burma. Vol. II. London: Taylor and Francis; p. 503.
- Posada D. 2008. jModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution. 25(7):1253–1256. doi: 10.1093/molbev/msn083
- Ronquist F, Teslenko M, Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP. 2012. Mrbayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 61:539–542. doi: 10.1093/sysbio/sys029
- Roos A, Loy A, de Silva P, Hajkova P, Zemanová B. 2015. Lutra lutra. The IUCN Red List of Threatened Species 2015.
- Sasaki H. 1995. History of river otters in Japan. In: Proceedings of Korea-Japan otter symposium. p. 16–17.
- Suzuki T, Yuasa H, Machida Y. 1996. Phylogenetic position of the Japanese river otter Lutra nippon inferred from the nucleotide sequence of 224 bp of the mitochondrial cytochrome b gene. Zool Sci. 13:621–626. doi: 10.2108/zsj.13.621
- Swofford DL. 2001. PAUP*. Phylogenetic analysis using parsimony (*and other methods). Version 4. Sunderland (MA): Sinauer Associates.
- Waku D, Segawa T, Yonezawa T, Akiyoshi A, Ishige T, Ueda M, Ogawa H, Sasaki H, Ando M, Kohno N, et al. 2016. Evaluating the phylogenetic status of the extinct Japanese otter in the basis of mitochondrial genome analysis. Plos One. DOI:10.1371/journal.pone.0149341.
- Wozencraft WC. 2005. Mammal species of the world: a taxonomic and geographic reference. 3rd ed. Baltimore (MD): Johns Hopkins University Press; p. 532–628.
- Yamamoto K, Ando M. 2011. Trends in otter-related newspaper articles in Japan over 135 Years. Proceedings of XIth International Otter Colloquium, IUCN Otter Specialist Group. Bull. 28:31–35.