
ABSTRACT
Drought is an important driver of change in aquatic ecosystems and generally acts as a filter to select biological traits capable of persisting under severe environmental circumstances. Drylands are highly vulnerable to climate change and increases in climate variability. Consequently, an increase in the processes of eutrophication, salinization, and habitat desiccation may be observed in response to the loss of the hydrological connectivity. The International Network on Limnology of Drylands (INLD) was created with the overall goal of understanding the functioning and conservation status of aquatic ecosystems in drylands globally. INLD was made official in 2016 through the International Society of Limnology. Its objectives are to (1) assess the current state of biological diversity in dryland aquatic ecosystems, (2) evaluate the multiple environmental stressors acting in drylands, and (3) develop models to predict effects of global change on drylands. This special issue, Limnology of Drylands, consists of 10 manuscripts focused on the effects of environmental pressures, including global warming, on the biodiversity and distribution of aquatic communities in drylands.
Introduction
More than one-third of the human population lives in areas where there are drylands, and these areas cover more than 41% of Earth's land surface, including hyperarid, arid, semiarid, and dry-subhumid areas (Millennium Ecosystem Assessment Citation2005). According to recent estimates, drylands may increase to 56% of the land surface by the end of the 21st century (Huang et al. Citation2016). This increase may cause several ecological and social problems, as well as climate change, affecting research, management, and policy decisions concerning drylands (Reynolds et al. Citation2007).
Several types of aquatic ecosystems are included in dryland regions, such as permanent and intermittent rivers, lakes, and rock pools, among others. Considering their elevated species richness and the high rate of endemism, aquatic ecosystems in semiarid and dry-subhumid areas are among the most biodiverse regions in the world (Gudka et al. Citation2014). They are strongly influenced by irregular rainfall patterns, where the spatial–temporal variability of precipitation increases with the level of aridity, influencing management and conservation practices (Williams Citation2000). Semiarid regions are considered more vulnerable to climate change, and some predictions show that an increased aridity trend is occurring more intensely in semiarid areas, as a function of a gradual decrease in precipitation and soil moisture, and increased drought frequency (Ji et al. Citation2014, Huang et al. Citation2016). This tendency, as well as rapid human population growth in dryland regions, will lead to an enhanced rate of desertification and increased vulnerability of aquatic ecosystems to deterioration (Rotenberg and Yakir Citation2010, Huang et al. Citation2016). Prolonged and extreme drought events have already threatened the biodiversity of dryland ecosystems and disrupted trophic interactions (Ledger et al. Citation2013).
Temporary lakes of dryland areas have different hydrological regimes, maintaining dry phases that can last a few days or months, with drying duration crucial for the establishment and maintenance of biodiversity (Naselli-Flores and Barone Citation2012). Frequent water level reductions, owing to prolonged droughts, pose a threat to systems through eutrophication and salinization in addition to leading to the proliferation of harmful algae (Brasil et al. Citation2016, Lins et al. Citation2016, Rocha Junior et al. Citation2018).
Intermittent rivers are the main type of lotic ecosystem in drylands, composing about half of the global river network (Datry et al. Citation2014). Although these rivers have been considered to have low species richness, the increased habitat heterogeneity and terrestrial connectivity associated with the drying of the main channel is a source of biodiversity and consequently compensates for the decreased lotic biodiversity (Larned et al. Citation2010). Rock pools on the riverbed can act as aquatic refuges for species not adapted to desiccation in drylands (Sheldon et al. Citation2010). Nevertheless, intermittent rivers in dryland areas are fragile and threatened by climate change, land-use changes, and water abstraction (Datry et al. Citation2014).
This special issue of Inland Waters (Limnology of Drylands) focuses on the effects of climate change and environmental pressures (naturally and anthropogenically driven) on the biodiversity and structure of aquatic communities in several dryland regions around the world.
International Network on Limnology of Drylands (INLD): highlights, hypotheses, and targets
Historically, human population growth in dryland areas has led to chronic ecosystem degradation, often expressed by severe reductions in the quality and quantity of water resources. Global warming will cause severe and irreversible impacts on Earth's ecosystems (IPCC Citation2014). Empirical observations for the period of 1948–2008 show that the world’s drylands have expanded and will continue to expand by the end of the 21st century (Feng and Fu Citation2013). For arid regions, the major expansion will take place over southwest North America and northern and southern Africa and Australia, and in semiarid regions it will occur in northern Mediterranean lands, southern Africa, and North and South America (Feng and Fu Citation2013). These shifts will increase water shortages and biodiversity loss at local, regional, and global scales and reduce ecosystem resilience to natural disturbances, leading to unprecedented changes in freshwater ecosystem services (e.g., McNeely Citation2003, Baho et al. Citation2017). Thus, the combined effects of global dryland expansion and increasing demand for water resources will magnify extreme droughts events in hyperarid, arid, semiarid, and dry-subhumid regions (Feng and Fu Citation2013).
In the face of these threats, INLD aims to understand how environmental stressors (i.e., water level reduction, eutrophication, salinization, and extreme climate events) affect community composition, distribution, and dynamics of dryland aquatic ecosystems. One of the main predictions is that climate change (e.g., rainfall reduction, water level reduction, and extreme drought events) and local human impacts (e.g., habitat destruction, deforestation, pollution, water abstraction, and land-use change) will synergistically change hydrological regimes, leading to a permanent state of low biodiversity and resilience in dryland aquatic ecosystems. The first steps of INLD are to elucidate how climate and anthropically driven environmental change affect drylands based on 3 assessments: (1) the current state of aquatic biological diversity; (2) how multiple environmental stressors affect aquatic communities; and (3) the development of predictive models for dryland aquatic ecosystems ().
Figure 1. Schematic presentation of the International Network on Limnology of Drylands (INLD) indicating global patterns and the functioning of biodiversity in dryland freshwater ecosystems.
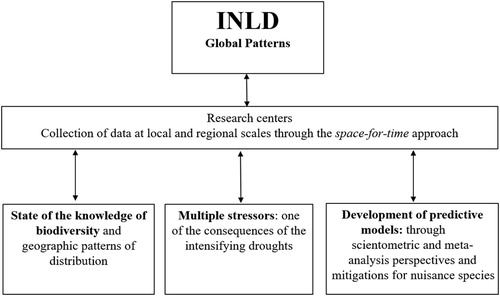
Droughts have previously been considered a pulse disturbance of high intensity and, because of climate change, they can now be considered a “ramp disturbance,” increasing in severity through time (Lake Citation2011). This ramp disturbance is likely to magnify the effects of eutrophication and salinization, among other impacts, selecting species best adapted to severe conditions. The assessment of these effects will require biological indicators (e.g., Chessman Citation2015). Thus, research on the life history of saline and freshwater species and their tolerance to prolonged drought, and other impacts, may help us to understand their biology and develop predictive models about their resilience and persistence of species in dry habitats (e.g., Chessman Citation2015, Kouba et al. Citation2016, Sarremejane et al. Citation2019).
Despite the large extension of dryland zones around the world, knowledge on the diversity of species and functioning of the temporary and permanent ecosystems is poorly distributed, creating gaps and preventing decisions regarding conservation plans in these areas. The persistence of the aquatic phase of waterbodies in dryland areas will decrease, causing direct mortality of the biota, leading to a reduction in the taxonomic and functional species richness. The gaps in knowledge and the increasing human and climate warming impacts on dryland aquatic ecosystems are the leading causes of failures in restoration practices, unraveling with ongoing decreases in ecosystem and organism resilience.
Contributions
Space-for-time substitution approach
Long-term data series are important tools to answer ecological and evolutionary questions that need broad spatial and temporal monitoring, especially when dealing with episodic and variable processes, such as extreme droughts and flood events (e.g., Dodds et al. Citation2012). The lack of temporal information (i.e., long-term data series) promotes serious misjudgments that can interfere not only with attempts to understand and predict changes but also with efforts to manage environments (Magnuson Citation1990).
On an evolutionary scale, the length of the dry phase is a critical filter, selecting biological traits (Matthews and Marsh-Matthews Citation2003). Species adaptations can be considered an ecological phenomenon, favoring physiological and morphological adjustments that amplify the competitive ability and compensate for severe changes occurring in dryland aquatic ecosystems (Williams Citation1985). Some morphological and physiological characteristics, in addition to some adaptations of organisms, can be associated with their ability to proliferate during the filling period and in other phases of the hydroperiod. Among these features, hatching, viability of resting eggs, high dispersal capacity of the adults, short life cycle, and generalist strategies enabling adaptation when habitat changes occur are more important for aquatic communities, such as zooplankton (Williams Citation1996).
Nevertheless, long-term studies assessing drought effects in temporary waterbodies are generally lacking, resulting in gaps in knowledge about the ecological memory of these ecosystems. Changes in the frequency and predictability of droughts caused by human activities and climate variations can be considered an “invisible place” (Magnuson Citation1990) or “invisible space” where the predictive potential is low (Swanson and Sparks Citation1990). In both cases, the absence of evolutionary and historical perspectives can produce results that range from highly uncertain to inconclusive.
Desiccation of rivers or wetlands can generate barriers and hinder biotic flows during drought (Bond and Cottingham Citation2008). During the wet season, however, the increased water level in these systems enhances their hydrological connections (Larned et al. Citation2010). With climate change, drylands are expected to experience extreme drought episodes, increasing habitat isolation, and hindering river flows and connections (Bonada et al. Citation2006).
Some metaphors may be used as a resource to show the relevance of the long-term studies in comprehending events with high interannual variability or high spatial variation, such as ice cover or drought events. Among these metaphors, the invisible present and the invisible place may indicate the relevance of historical databases in multiple and broad scales, preventing errors concerning data interpretation in the short term (Magnuson Citation1990, Swanson and Sparks Citation1990) and allowing distinction between natural and human-induced ecological events (Larson et al. Citation2007). The space-for-time substitution approach has been used to infer past or future trajectories of ecological systems (Blois et al. Citation2013). Patterns or trends in space are used to predict temporal changes, such as the role of macrophytes in shallow lakes in different climate zones (Meerhoff et al. Citation2007) or the effects of rainfall reduction on abiotic and biotic variables (Blois et al. Citation2013, Costa et al. Citation2019). From this insight, INLD aims to develop 3 main approaches: site monitoring (see Supplementary Material), implementation of experiments, and paleolimnological investigations.
Some contributions in this special issue address monitoring at a broad spatial scale to understand the distribution and abundance patterns of species and communities (e.g., Ilhéu et al. forthcoming Citation2020, Lanfranco et al. Citation2020, Silva et al. Citation2020a, Citation2020b). Other contributions use the space-for-time substitution approach for ecosystems located at different latitudes to explain the distribution of organisms along environmental gradients. In this context, Lanfranco (Citation2020) investigated the influence of morphometric and geographic factors on the structure and composition of plant communities in 56 temporary freshwater rock pools in the Maltese Islands, Europe. In summary, the most influential predictors of community composition varied across the landscape suggesting that pool “location” can be an important determinant of community structure and composition because pool morphometry would favor some functional forms and species over others.
Submerged macrophytes play a crucial role in increasing habitat and food webs complexity in lakes (Manatunge et al. Citation2000, Thomaz and Cunha Citation2010), controlling nuisance species (Hilt and Gross Citation2008) and working as shelter and feeding sites for fish and zooplankton species (Jeppesen et al. Citation1997, Meerhoff et al. Citation2006). However, the vulnerability of these communities to global warming has not yet been clarified (Jeppesen et al. Citation2014). One of the gaps in ecosystem understanding for tropical arid and semiarid areas is the influence of submerged macrophytes on the attributes of algal communities, mainly concerning the colonization of periphytic algae on substrates, control of phytoplankton biomass, and maintenance of clear-water state (Hilt and Gross Citation2008). In this special section, Silva et al. (Citation2020b) indicated that the morphological and architectural differences between the charophyte genera Chara and Nitella may induce differences in the colonization by periphyton in 19 freshwater ecosystems in the semiarid region of Brazil, indicating that the colonization process may be driven by the structure of the substrate, which selects different life forms and species.
State of knowledge of biodiversity and biogeographic patterns in dryland zones
Spatial patterns induced by drought
Aquatic ecosystems in dryland regions can be classified according to the length of their hydroperiod (Williams Citation2006). Droughts, as an unpredictable and unstable forcing, can favor resilient biota well adapted to deal with water loss following more predictable periods (Brock et al. Citation2003, Williams Citation2006). Therefore, aquatic fauna and flora with favorable physiological and morphological features will be better able to compensate for severe changes in these ecosystems (Williams Citation1985).
The length of the drought phase is a critical factor influencing the abundance and composition of species (Williams Citation2006). Intermittent flow may induce a fragmented distribution of resources and communities (Leibold et al. Citation2004). Farias et al. (Citation2020) demonstrated that nonconnected temporary pools in dryland river segments and streams can be important for regional diversity under drought conditions. For some larger fish species, larger pools during periods of desiccation provide better environmental conditions (Ilhéu et al. Citation2020). Fish broodstock is assumed to migrate up tributaries when desiccation of waterbodies or reservoirs flushing begins, but broodstock may also bury itself in river sand/mud (Kolding et al. Citation2016) allowing its survival during unfavorable seasons.
Some studies showed the successful recruitment of fish in intermittent rivers even in dry years (Silva et al. Citation2020a). Therefore, despite the unpredictable regime, species from dryland rivers tend to have a rapid population recovery following droughts because of higher fecundity (Mallen-Cooper and Stuart Citation2003, Silva et al. Citation2020a), earlier breeding, and drought-resistant eggs (Williams Citation2006).
Multiple stressor analysis
The wide typological diversity of organisms from the humid areas in drylands can be considered hotspots sustained by the provision of environmental services (Williams Citation2006). Temporary wetlands are the most supersaturated in carbon dioxide (CO2) among the aquatic ecosystems in the global carbon budget (Holgerson Citation2015). But, despite the relevance to the carbon fluxes at the local and global scale, temporary wetlands are neglected (Holgerson Citation2015). For a better understanding of carbon fluxes in drylands, more accurate estimates of the carbon and carbon greenhouse gases (CO2 and CH4) are needed. In this regard, Morant et al. (Citation2020) showed that carbon fluxes, which can assist with the application of restoration and management practices in dryland ecosystems.
Climate change is affecting global hydrological cycles and influencing the dynamics of intermittent and temporary aquatic in dryland ecosystems, with severe impacts related to the reduction of water levels and changes in hydroperiod (Landkildehus et al. Citation2014). Expected reductions in water levels are likely to result in dramatic changes in aquatic communities because of increasing salinity and evapotranspiration, leading to high nutrient levels (Rocha Junior et al. Citation2018, Costa et al. Citation2019); however, the critical limit for those biological changes is still unknown. Some hypotheses indicate that Mediterranean (Jeppesen et al. Citation2015) and Australian (Nielsen et al. Citation2003) dryland lakes may have higher salinity levels in the future, a pattern expected to occur in other dryland regions around the world ().
Figure 2. Potential changes in dryland aquatic ecosystems and associated landscape in response to severe droughts and related effects, especially drastic water level reduction of lakes.
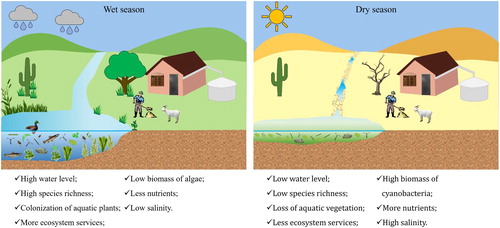
Zadereev et al. (Citation2020) assessed the conditions of saline lakes in Europe and Central Asia based on the anthropogenic and climate change drivers leading to reductions in water level. This synergy of drivers may be a great challenge to the management and sustainable use of saline lakes in that region.
Hulot et al. (Citation2020) analyzed 30 salt pans in Zimbabwe to show how water level fluctuations can influence the aquatic communities and food web structure by the replacement or loss of species. They found seasonal changes in the food webs associated with the simplification of networks during the dry period. Among the main drivers of this change, a drastic water level reduction induced an increase of potential pollutants, mainly ammonium. In this regard, sublethal concentrations of toxicants (e.g., copper, ammonium, or cyanotoxins) affect traits related to sexual reproduction and diapause egg production, as well as genetic diversity, in several zooplankton species (Hairston et al. Citation1999, Liang et al. Citation2018). This point highlights the ecological and evolutionary consequences of the interactive effects of multiple stressors (e.g., pollutants, toxins, higher temperatures) on aquatic biodiversity (Aránguiz-Acuña and Pérez-Portilla Citation2017).
The transition from a clear to a turbid water state is associated with a decrease in submerged vegetation and changes in the structure of fish communities in shallow lakes (Scheffer et al. Citation1993, Scheffer and Carpenter Citation2003). However, droughts can induce a transient state that represents the “ghost of a stable state,” keeping the lake in an unstable, clear state most of the time, despite high nutrient concentrations (Scheffer and van Nes Citation2007). Usually, water level reductions also have negative impacts, causing, for example, an increase of cyanobacterial biomass (Bakker and Hilt Citation2016), threatening the aquatic biota, enhancing eutrophication, and reducing ecosystem services. Among the main consequences of the severe water level reductions are increased total phosphorus concentrations, salinization, low water transparency, and loss of services (Jeppesen et al. Citation2015; ).
Figure 3. Schematic presentation of changes caused by the extreme water level changes in dry zones.
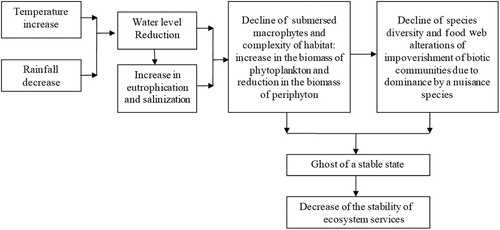
Development of predictive models
Human-induced climate change is predicted to cause both an increase in temperature (resulting in increased evaporation) and a decrease in the amount of precipitation in most arid and semiarid regions worldwide (Ragab and Prudhomme Citation2002), which will increase aridity, frequency, and intensity of droughts. These shifts in local and regional climate are expected to reduce the flushing waterbodies and alter the local nutrient cycles, potentially resulting in periods of substantial eutrophication (Nielsen and Brock Citation2009).
In Mediterranean temporary and permanent dryland ponds, studies suggest that the temperature and precipitation patterns determine the structure and dynamics of phytoplankton on a regional scale (e.g., Naselli-Flores and Barone Citation2012). In these habitats, longer periods of relatively high temperatures and consequent water level reduction favor algal growth (Naselli-Flores and Barone Citation2005). Indeed, this pattern has also been demonstrated in several constructed lakes in northeast Brazil (e.g., Brasil et al. Citation2016). Nonetheless, with predictions of a warmer and drier world by the end of the 21st century (IPCC Citation2014), water resources will probably reduce in quantity and quality, increasing susceptibility to salinization and eutrophication, in addition to favoring cyanobacterial blooms in freshwaters (Jeppesen et al. Citation2015, Brasil et al. Citation2016).
In the last decade, global warming, increasingly contaminated waters, and illegal transportation have been the main elements favoring the expansion of invasive species (e.g., cyanobacteria) in arid and semiarid regions (e.g. Moura et al. Citation2018). This trend is expected for the coming years, considering that climate change will intensify the threats (Early et al. Citation2016), particularly in constructed reservoirs regarded as hotspots for invasive species (Rahel and Olden Citation2008). Naselli-Flores and Marrone (Citation2019) showed how artificial lakes and ponds in a semiarid Mediterranean island are more susceptible to the establishment of non-native species because these environments are more recent than natural environments and have fewer of biological filters to prevent colonization by invaders. Also, the high number of constructed aquatic ecosystems in dryland regions facilitates invasive species by increasing the surface area for colonization (Naselli-Flores and Marrone Citation2019).
Predictive models would be useful to understand and implement restoration programs that include the interactive effect of environmental variables and aquatic communities. Recent studies have shown the influence of nutrients, temperature, and food quality on the abundance, composition, and diversity of the zooplankton during a long-term restoration program (Li et al. Citation2019).
In arid and semiarid zones, some studies have suggested a strong correlation between submerged macrophytes and flood intensity, demonstrating high resistance and resilience in the face of low-impact hydrological disturbances (Maltchik and Florin Citation2002, Pedro et al. Citation2006). By contrast, severe flooding may induce low resilience in these communities (Maltchik and Florin Citation2002). Aquatic communities of the temporary ecosystems are resilient because they can survive disturbances, recovering biomass through attributes that allow proliferation when the area is reflooded (Brock et al. Citation2003) and persisting through the production of dormant propagules or resistance structures (Brock et al. Citation2003, Williams Citation2006).
Over the last few decades, several models have been developed to prevent or reduce the effects of eutrophication (e.g., Mooij et al. Citation2010). The most widely accepted and applied conceptual model is associated with alternative states in shallow lakes (Scheffer and Carpenter Citation2003, Scheffer and Jeppesen Citation2007). These models predict that changes in the abiotic components would have little effect until a nutrient threshold is reached, represented by a rapid and potentially irreversible transition in the trophic structure and resilience of the system (Scheffer and Carpenter Citation2003). Barbosa et al. (Citation2020) assessed the effects of allochthonous disturbances on phosphorus release in 2 shallow lakes by constructing models using partial least squares path modeling. Those models indicated that the submerged macrophyte cover acted as a stabilizing force, reducing the phosphorus released from the sediments and preventing shifts between the stable states.
Conclusions and perspectives: challenges for INLD
Ongoing climate change and global warming may be magnifying eutrophication and salinization in dryland aquatic ecosystems, owing to more extreme and more frequent drought events. Among the consequences linked to unpredictable and extreme drought events are (1) the advent of nuisance species (i.e., toxic cyanobacteria), (2) reduction of the complexity of the food webs, and (3) loss of ecosystem resilience.
INLD intends to expand the construction of predictive models through scientometric and metanalysis approaches, identifying patterns using already published databases. INLD also intends to address the consequences of climate change to help derive critical limits of water level reductions that do not result in the degradation of biodiversity and ecosystem services (e.g., Meerhoff et al. Citation2012). The development of initiatives to reduce eutrophication, for example, might help mitigate climate change effects (Moss et al. Citation2011). Therefore, INLD considers performing simultaneous standardized mesocosm experiments manipulating water level, biological communities, nutrients, and salinity in different dryland zones of the world to support the development of ecological models that infer future trajectories of freshwater systems as a function of rainfall reductions and worsening droughts. This approach may be useful to assess how critical changes in rainfall will affect water quality and freshwater community structure and composition in drylands, as well as to generate knowledge of the main mechanisms regulating those processes and develop strategies to mitigate the expected negative effects.
Under climate change scenarios, patch isolation through habitat drying is likely to occur, increasing vulnerability of rare and endemic species and leading to the loss of biodiversity. Endemic species vulnerability can also be magnified by the introduction of exotic species and eutrophication, which can lead to habitat homogenization. Strategies need to be developed that enhance biodiversity and ecosystem services as well as reduce fragmentation from drought in rivers and wetlands.
Supplemental Material
Download MS Word (23.4 KB)Acknowledgements
We dedicate this Special Issue to researchers who have developed their investigations in dry regions around the world, especially Dr. José Luiz Atayde (Coca) who contributed and supported in the creation and development of the INLD. The manuscripts in this special issue of Inland Waters, Limnology of Drylands, are the result of international efforts of the INLD working group. Our special thanks to the members and collaborators who contributed valuable papers. Also, our sincere thanks to the International Society of Limnology for support, especially to Dr. Tamar Zohary and Dr. David Hamilton. Finally, we express our great thanks to all anonymous reviewers for their valuable assistance.
Disclosure statement
No conflict of interest was reported by the authors.
References
- Aránguiz-Acuña A, Pérez-Portilla P. 2017. Metal stress in zooplankton diapause production: post-hatching response. Ecotoxicology. 26(3):329–339.
- Baho DL, Allen CR, Garmestani A, Fried-Petersen H, Renes SE, Gunderson L, Angeler DG. 2017. A quantitative framework for assessing ecological resilience. Ecol Soc. 22(3): art17.
- Bakker ES, Hilt S. 2016. Impact of water-level fluctuations on cyanobacterial blooms: options for management. Aquat Ecol. 50(3):485–498.
- Barbosa LG, Alves RMA, Santos JPO, Dantas EW. 2020. Role of submerged macrophytes in phosphorus stabilization of sediments in shallow lakes from the Brazilian semiarid region. Inland Waters. doi.org/10.1080/20442041.2020.1814090
- Blois JL, Williams JW, Fitzpatrick MC, Jackson ST, Ferrier S. 2013. Space can substitute for time in predicting climate-change effects on biodiversity. P Natl Acad Sci USA. 110(23):9374–9379.
- Bonada N, Rieradevall M, Prat N, Resh VH. 2006. Benthic macroinvertebrate assemblages and macrohabitat connectivity in Mediterranean-climate streams of northern California. J N Am Benthol Soc. 25(1):32–43.
- Bond NR, Cottingham P. 2008. Ecology and hydrology of temporary streams: implications for sustainable water management. Canberra (Australia): eWater Technical Report, Vol. 29.
- Brasil J, Attayde JL, Vasconcelos FR, Dantas DDF, Huszar VLM. 2016. Drought-induced water-level reduction favors cyanobacteria blooms in tropical shallow lakes. Hydrobiologia. 770(1):145–164.
- Brock MA, Nielsen DL, Shiel RJ, Green JD, Langley JD. 2003. Drought and aquatic community resilience: the role of eggs and seeds in sediments of temporary wetlands. Freshwater Biol. 48(7):1207–1218.
- Chessman BC. 2015. Relationships between lotic macroinvertebrate traits and responses to extreme drought. Freshwater Biol. 60(1):50–63.
- Costa MRA, Menezes RF, Sarmento H, Attayde JL, Sternberg LdS, Becker V. 2019. Extreme drought favors potential mixotrophic organisms in tropical semi-arid reservoirs. Hydrobiologia. 831(1):43–54.
- Datry T, Larned ST, Tockner K. 2014. Intermittent rivers: a challenge for freshwater ecology. Bioscience. 64(3):229–235.
- Dodds WK, Robinson CT, Gaiser EE, Hansen GJA, Powell H, Smith JM, Morse NB, Johnson SL, Gregory SV, Bell T, et al. 2012. Surprises and insights from long-term aquatic data sets and experiments. Bioscience. 62(8):709–721.
- Early R, Bradley BA, Dukes JS, Lawler JJ, Olden JD, Blumenthal DM, Gonzalez P, Grosholz ED, Ibañez I, Miller LP, et al. 2016. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat Commun. 7(1):12485.
- Farias RL, Stenertc C, Maltchik L, Medeiros ESF. 2020. Partitioning of macroinvertebrate assemblages across temporary pools in an intermittent dryland river. Inland Waters. doi.org/10.1080/20442041.2020.1738841
- Feng S, Fu Q. 2013. Expansion of global drylands under a warming climate. Atmos Chem Phys. 13(19):10081–10094.
- Gudka M, Davies J, Poulsen L, Schulte-Herbrüggen B, MacKinnon K, Crawhall N, Henwood WD, Dudley N, Smith J. 2014. Conserving dryland biodiversity: a future vision of sustainable dryland development. Biodiversity. 15(2–3):143–147.
- Hairston NG, Lampert W, Cáceres CE, Holtmeier CL, Weider LJ, Gaedke U, Fischer JM, Fox JA, Post DM. 1999. Rapid evolution revealed by dormant eggs. Nature. 401(6752):446–446.
- Hilt S, Gross EM. 2008. Can allelopathically active submerged macrophytes stabilise clear-water states in shallow lakes? Basic Appl Ecol. 9(4):422–432.
- Holgerson MA. 2015. Drivers of carbon dioxide and methane supersaturation in small, temporary ponds. Biogeochemistry. 124(1–3):305–318.
- Huang J, Yu H, Guan X, Wang G, Guo R. 2016. Accelerated dryland expansion under climate change. Nat Clim Change. 6:166–171.
- Hulot FD, Msiteli-Shumbac S, Bonhomme M, Legendre S. 2020. Temporal changes in food web structure in semiarid waterholes. Inland Waters. doi.org/10.1080/20442041.2020.1754112
- Ilhéu M, Silva J, Morais M, Matono P, Bernardo JM. 2020. Summer stream pools types in an intermittent river network: environmental drivers and fish assemblages. Inland Waters. Forthcoming.
- [IPCC] Intergovernmental Panel on Climate Change. 2014. Summary for policymakers. In: Field CB, Barros VR, Dokken DJ, Mach KJ, Mastrandrea MD, Bilir TE, Chatterjee M, Ebi KL, Estrada YO, Genova RC, et al., editors. Climate change 2014: impacts, adaptation, and vulnerability. Contribution of Working Group II to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge (UK): Cambridge University Press; pp. 1–32.
- Jeppesen E, Brucet S, Naselli-Flores L, Papastergiadou E, Stefanidis K, Nõges T, Nõges P, Attayde JL, Zohary T, Coppens J, et al. 2015. Ecological impacts of global warming and water abstraction on lakes and reservoirs due to changes in water level and related changes in salinity. Hydrobiologia. 750(1):201–227.
- Jeppesen E, Meerhoff M, Davidson TA, Trolle D, Søndergaard M, Lauridsen TL, Beklioglu M, Brucet S, Volta P, González-Bergonzoni I, et al. 2014. Climate change impacts on lakes: an integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J Limnol. 73(s1):88–111.
- Jeppesen E, Peder Jensen J, Søndergaard M, Lauridsen T, Junge Pedersen L, Jensen L. 1997. Top-down control in freshwater lakes: the role of nutrient state, submerged macrophytes and water depth. In: Kufel L, Prejs A, Rybak JI, editors. Shallow lakes ‘95. Dordrecht: Springer Netherlands; p. 151–164.
- Ji F, Wu Z, Huang J, Chassignet EP. 2014. Evolution of land surface air temperature trend. Nat Clim Change. 4:462–466.
- Kolding J, van Zwieten PAM, Marttin F, Poulain F. 2016. Fisheries in the drylands of Sub-Saharan Africa: Fish come with the rains. Rome: Food and Agriculture Organization of the United Nations. Circular No. 1118.
- Kouba A, Tíkal J, Císař P, Veselý L, Fořt M, Příborský J, Patoka J, Buřič M. 2016. The significance of droughts for hyporheic dwellers: evidence from freshwater crayfish. Sci Rep. 6(1):26569.
- Lake PS. 2011. Drought and aquatic ecosystems: effects and responses. Chichester (UK): Wiley-Blackwell.
- Landkildehus F, Søndergaard M, Beklioglu M, Adrian R, Angeler DG, Hejzlar J, Papastergiadou E, Zingel P, Çakiroğlu AI, Scharfenberger U, et al. 2014. Climate change effects on shallow lakes: design and preliminary results of a cross-European climate gradient mesocosm experiment. Est J Ecol. 63(2):71.
- Lanfranco S, Bellia AF, Cuschieri P. 2020. Influence of morphometric and geographic factors on community composition of temporary freshwater rockpools in Maltese Islands. Inland Waters. doi.org/10.1080/20442041.2019.1704152
- Larned ST, Datry T, Arscott DB, Tockner K. 2010. Emerging concepts in temporary-river ecology. Freshwater Biol. 55:717–738.
- Larson GL, Hoffman RL, McIntire DC, Buktenica MS, Girdner SF. 2007. Thermal, chemical, and optical properties of Crater Lake, Oregon. Hydrobiologia. 574:69–84.
- Ledger ME, Brown LE, Edwards FK, Milner AM, Woodward G. 2013. Drought alters the structure and functioning of complex food webs. Nat Clim Change. 3(3):223–227.
- Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, et al. 2004. The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett. 7(7):601–613.
- Li Y, Liu L, Cui S, Chen F. 2019. Long-term effects of nutrient changes on rotifer communities in a subtropical lake. Limnology. 20(2):191–201.
- Liang Y, Lu X, Min Y, Liu L, Yang J. 2018. Interactive effects of microcystin and ammonia on the reproductive performance and phenotypic traits of the rotifer Brachionus calyciflorus. Ecotoxicol Environ Saf. 147:413–422.
- Lins RPM, Barbosa LG, Minillo A, De Ceballos BSO. 2016. Cyanobacteria in a eutrophicated reservoir in a semi-arid region in Brazil: dominance and microcystin events of blooms. Braz J Bot. 39:583–591.
- Magnuson JJ. 1990. Long-term ecological research and the invisible present. Bioscience. 40(7):495–501.
- Mallen-Cooper M, Stuart IG. 2003. Age, growth and non-flood recruitment of two potamodromous fishes in a large semiarid/temperate river system. River Res Appl. 19(7):697–719.
- Maltchik L, Florin M. 2002. Perspectives of hydrological disturbance as the driving force of Brazilian semiarid stream ecosystems. Acta Limnol Bras. 14(3):35–41.
- Manatunge J, Asaeda T, Priyadarshana T. 2000. The influence of structural complexity on fish-zooplankton interactions: a study using artificial submerged macrophytes. Environ Biol Fishes. 58(4):425–438.
- Matthews WJ, Marsh-Matthews E. 2003. Effects of drought on fish across axes of space, time and ecological complexity. Freshwater Biol. 48(7):1232–1253.
- McNeely JA. 2003. Biodiversity, war, and tropical forests. J Sustain For. 16(3-4):1–20.
- Meerhoff M, Clemente JM, de Mello FT, Iglesias C, Pedersen AR, Jeppesen E. 2007. Can warm climate-related structure of littoral predator assemblies weaken the clear water state in shallow lakes? Global Change Biol. 13:1888–1897.
- Meerhoff M, Fosalba C, Bruzzone C, Mazzeo N, Noordoven W, Jeppesen E. 2006. An experimental study of habitat choice by Daphnia: plants signal danger more than refuge in subtropical lakes. Freshwater Biol. 51(7):1320–1330.
- Meerhoff M, Mello FT, Kruk C, Alonso C, Gonzalez-Bergonzoni I, Pablo Pacheco J, Lacerot G, Arim M, Beklioglu M, Brucet S, et al. 2012. Environmental warming in shallow lakes: a review of potential changes in community structure as evidenced from space-for-time substitution approaches. Adv Ecol Res. 46:259–349.
- Millennium Ecosystem Assessment. 2005. Ecosystems and human well-being: synthesis. Washington (DC): Island Press.
- Mooij W, Trolle D, Jeppesen E, Arhonditsis G, Belolipetsky P, Chitamwebwa DBR, Degermendzhy AG, DeAngelis DL, De Senerpont Domis LN, Downing AS, et al. 2010. Challenges and opportunities for integrating lake ecosystem modelling approaches. Aquat Ecol. 44:633–667.
- Morant D, Picazo A, Rochera C, Santamans AC, Miralles-Lorenzo J, Camacho A. 2020. Linkage of the conservation status to the carbon balance of semiarid Mediterranean coastal wetlands. Inland Waters. Forthcoming.
- Moss B, Kosten S, Meerhoff M, Battarbee RW, Jeppesen E, Mazzeo N, Havens K, Lacerot G, Liu Z, De Meester L, et al. 2011. Allied attack: climate change and eutrophication. Inland Waters. 1(2):101–105.
- Moura AN, Aragão-Tavares NKC, Amorim CA. 2018. Cyanobacterial blooms in freshwater bodies from a semiarid region, northeast Brazil: a review. J Limnol. 77(2):179–188.
- Naselli-Flores L, Barone R. 2005. Water-level fluctuations in Mediterranean reservoirs: setting a dewatering threshold as a management tool to improve water quality. Hydrobiologia. 548:85–99.
- Naselli-Flores L, Barone R. 2012. Phytoplankton dynamics in permanent and temporary Mediterranean waters: Is the game hard to play because of hydrological disturbance? Hydrobiologia. 698(1):147–159.
- Naselli-Flores L, Marrone F. 2019. Different invasibility of permanent and temporary waterbodies in a semiarid Mediterranean Island. Inland Waters. 9(4):411–421.
- Nielsen DL, Brock MA, Rees GN, Baldwin DS. 2003. Effects of increasing salinity on freshwater ecosystems in Australia. Aust J Bot. 51(6):655.
- Nielsen DL, Brock MA. 2009. Modified water regime and salinity as a consequence of climate change: prospects for wetlands of Southern Australia. Clim Change. 95(3–4):523–533.
- Pedro F, Maltchi L, Bianchini Junior I. 2006. Hydrologic cycle and dynamics of aquatic macrophytes in two intermittent rivers of the semi-arid region of Brazil. Braz J Biol. 66(2B):575–585.
- Ragab R, Prudhomme C. 2002. SW—soil and water: climate change and water resources management in arid and semi-arid regions: prospective and challenges for the 21st century. Biosyst Eng. 81(1):3–34.
- Rahel FJ, Olden JD. 2008. Assessing the effects of climate change on aquatic invasive species. Conserv Biol. 22(3):521–533.
- Reynolds JF, Stafford Smith DM, Lambin EF, Turner BL, Mortimore M, Batterbury SPJ, Downing TE, Dowlatabadi H, Fernández RJ, Herrick JE, et al. 2007. Global desertification: building a science for dryland development. Science. 316:847–851.
- Rocha Junior CAN, Costa MRA, Menezes RF, Attayde JL, Becker V. 2018. Water volume reduction increases eutrophication risk in tropical semi-arid reservoirs. Acta Limnol Bras. 30:e106.
- Rotenberg E, Yakir D. 2010. Contribution of semi-arid forests to the climate system. Science. 327(5964):451–454.
- Sarremejane R, Stubbington R, Dunbar MJ, Westwood CG, England J. 2019. Biological indices to characterize community responses to drying in streams with contrasting flow permanence regimes. Ecol Indic. 107:105620.
- Scheffer M, Carpenter SR. 2003. Catastrophic regime shifts in ecosystems: linking theory to observation. Trends Ecol Evol. 18(12):648–656.
- Scheffer M, Hosper SH, Meijer M-L, Moss B, Jeppesen E. 1993. Alternative equilibria in shallow lakes. Trends Ecol Evol. 8(8):275–279.
- Scheffer M, Jeppesen E. 2007. Regime shifts in shallow lakes. Ecosystems. 10:1–3.
- Scheffer M, van Nes EH. 2007. Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia. 584(1):455–466.
- Sheldon F, Bunn SE, Hughes JM, Arthington AH, Balcombe SR, Fellows CS. 2010. Ecological roles and threats to aquatic refugia in arid landscapes: dryland river waterholes. Mar Freshw Res. 61:885.
- Silva FO, Neto FRA, Silva SH, Silva JO, Filho EZ, Prado IG, Peressin A, Pelicice F. 2020a. Recruitment dynamics of a migratory fish in a semiarid river system. Inland Waters. doi.org/10.1080/20442041.2020.1805977
- Silva IC, Carvalho NM, Moura AN, Dantas ÊW. 2020b. Relationships between epiphyton and macroalgae (Characeae) in tropical reservoirs. Inland Waters. doi.org/10.1080/20442041.2020.1789417
- Swanson FJ, Sparks RE. 1990. Long-term ecological research and the invisible place. Bioscience. 40:502–508.
- Thomaz SM, Cunha Ed. 2010. The role of macrophytes in habitat structuring in aquatic ecosystems: methods of measurement, causes and consequences on animal assemblages’ composition and biodiversity. Acta Limnol Bras. 22(02):218–236.
- Williams DD. 1996. Environmental constraints in temporary fresh waters and their consequences for the insect fauna. J N Am Benthol Soc. 15(4):634–650.
- Williams DD. 2006. The biology of temporary waters. Oxford (UK): Oxford University Press.
- Williams WD. 1985. Biotic adaptations in temporary lentic waters with special reference to those in semi-arid regions. In: Davies BR, Walmsley RD, editors. Perspectives in Southern Hemisphere limnology. Dordrecht (Netherlands): Dr. W. Junk; p. 85–110.
- Williams WD. 2000. Dryland lakes. Lakes Reserv Res Manag. 5:207–212.
- Zadereev E, Lipka O, Karimov B, Krylenko M, Elias V, Pinto IS, Alizade V, Anker Y, Feest A, Kuznetsova D, Mader A, et al. 2020. Overview of past, current and future ecosystem and biodiversity trends of inland saline lakes of Europe and Central Asia. Inland Waters. doi.org/10.1080/20442041.2020.1772034