Abstract
The Kudremukh National Park in the central Western Ghats (India) is a mega-biodiversity hotspot. However, the dependence on forests of tribal and non-tribal settlements in the core area of the park has resulted in forest fragmentation, posing a threat to the endemic flora. The study focuses on the disturbed forest ecosystem in the park. Using the belt transect method, we studied the vegetation structure and floristic composition of forests in tribal and non-tribal settlement areas. We compared species diversity, richness, dominance and stand quality of the park with an undisturbed nearby forest. Due to harvesting practices of the rural communities, the percentage of light-demanding species was higher in tribal and non-tribal forest compared those in with the undisturbed forest. Differences in species composition were largely due to extensive establishment of light-demanding or pioneer forest species. The pioneer forest species pose a threat to endemic species of the region by suppressing seedling growth and establishment. Our observations will help forest functionaries to prepare site-specific restoration plans. Primary forest species such as Myristica dactyloides, Palaquium ellipticum, Garcinia gummi-gutta and Poeciloneuron indicum can be used in the restoration process. These species help to maintain forest ecosystem stability and improve provision of forest ecosystem services.
Introduction
The Convention on Biological Diversity (WRI et al. Citation1992) identified mechanisms that affect current levels of biodiversity, nearly all of which include significant human components. The extraction of timber and fuelwood by rural communities has hitherto been within the carrying capacity of surrounding forests. With the increase in human population, these activities have exceeded the carrying capacity of the forests. It is, however, inferred that harvesting of timber from a natural forest affects its structure and dynamics (Whitmore and Sayer Citation1992). Young forests or plantations that develop soon after harvesting generally contribute to higher biodiversity (Kimmins Citation1992).
The rain forests of the Western Ghats mountains are unique because of their geographical location, stable geological history, stable climate, heavy rainfall and good soil conditions that support a variety of tropical forest ecosystems (Pascal Citation1988). Rich in biodiversity, this area harbours numerous medicinal plants and important genetic resources for the wild relatives of grains, fruits and spices (Bawa et al. Citation2007). The heterogeneity of habitat and vegetation patterns that are found in this area provides ideal conditions for the occurrence of many amphibian species (Krishnamurthy et al. Citation2001). In recent years, mining activity, coffee and other plantations have caused large-scale encroachment of Kudremukh National Park (KNP) (Hussain Citation2003), exposing protected areas to human interventions, proving to be a major threat to the vast plant genetic resources of the region. The regeneration of shade-loving primary forest species in the Western Ghats region is particularly affected through ongoing human activities (Parthasarathy Citation1999).
The study area, Karchar, is located in the KNP of Chikkamaglur district, in the central Western Ghats of South India and is the home of the Gowdlu tribe. There is sparse distribution of families – nearly three families per 1 km2. Communities residing here date back to as early as AD 4, practicing shifting cultivation, hunting and utilising natural forest resources for their livelihoods (Gadgil and Subhas Chandran Citation1988). In recent decades, non-tribal communities have also become dependent on these forests for sustenance and have converted part of forest area into Areca plantations. This intensification of agricultural production to meet the needs of the rapidly increasing population is the result of the dependence of non-tribal communities on forests for sustenance (Venkatesh Nayak et al. Citation2000).
In this study we evaluated the impact of different intensities of anthropogenic activity on the composition, diversity and stand quality of forest tree species in the KNP. We compared results from two differently managed sites (tribal and non-tribal plantation) with that of undisturbed forest.
Study area and climate
The study area lies within the KNP Western Ghats of Karnataka state (South India). It covers an area of 600.34 km2 and lies between 13°1′–13° 29′ N and 75° 0′–75° 30′ E. The soils are mainly Inceptisols and Ultisols (Bourgeon Citation1989). The climate is typical tropical, with an annual rainfall ranging from 600 to 800 cm. The maximum temperature varies from 21°C to 34°C during April–July, while the minimum temperature ranges from 12°C to 18°C between January and May.
Material and methods
Vegetation studies
For vegetation sampling, a belt transect was used to investigate the distribution of individuals in relation to a particular area (Hill et al. Citation2005). Belt transects of 500 m × 10 m were laid in three sites, which differed in the way in which they were utilised and managed: tribal settlements (communities dating back to AD 4, practicing shifting agriculture, hunting and utilising forest resources for livelihoods), non-tribal settlements (communities dependent on forests for sustenance and who have converted part of the forest area into Areca plantations) and intact areas (undisturbed forest). The total sample area at each plot was 0.15 ha. Each plot was divided into 15 sub-plots measuring 10 m × 10 m, laid at 10 m distances inside the belt transect. The girth at breast height of mature trees (diameter at breast height (DBH) > 30 cm) and saplings (DBH between 10 cm and 30 cm) and collar diameter of seedlings (girth below 10 cm) were recorded.
The data on vegetation were analysed for relative frequency, density and dominance, and their sum represents the importance value index (IVI) of the species (Kershaw Citation1973). Species diversity was calculated using the formula of Shannon and Wiener (Citation1963) as
Determination of stand quality
Using a literature survey (Gamble Citation1928; Chandrashekaran Citation1960; Rai Citation1979; Rai and Proctor Citation1986; Chandrashekara and Ramakrishnan Citation1994), we categorised species into three groups. Each group was assigned a numerical value, viz., Pioneer index 1 (primary species) for the group requiring a small gap for regeneration; Pioneer index 2 (early secondary) for the species whose seedlings establish in small gaps but need small- to medium-sized gaps to grow; and Pioneer index 3 (late secondary) for the group of strong light demanders. We adopted the procedures from Whitmore (Citation1989) and Chandrashekara (Citation1998) to indicate intensity of disturbance.
Results
Characteristics of the vegetation
Our study recorded 63 tree species belonging to 53 genera and 28 families from 45 sub-plots in 0.45 ha. In the tribal plot, we found 33 species, with Lophopetalum wightianum (85.7 IVI) as the dominant mature tree, followed by Dimocarpus longon (24.6) and Persea macarantha (16.8). In the non-tribal plot, we recorded 29 species, with dominance of Aglaia anamallayana (47.6) followed by D. longon (42.3) and Mimusops elengi (27.2). Poeciloneuron indicum was abundant in the undisturbed forest (106.6), followed by Cleistanthus travancorensis (20.1) and Syzygium rubicundum (18.6), with a total of 34 tree species. The shade-loving primary tree species such as Myristica dactyloides, Hopea canarensis, Calophyllus polyanthus and Agrostachis indica were recorded only in undisturbed forest plots. However, light-demanding tree species such as Mallotus philippensis, Macaranga peltata and Callicarpa tomentosa were only recorded in disturbed plots ().
Table 1. Pioneer index (PI) and importance value index (IVI) of tree species in three different areas of Kudremudh National Park, South India
Overall tree species richness data indicate that higher numbers of species or all size classes were found in undisturbed forest, whereas tree density of the regenerative class (seedlings) was higher in disturbed forest plots, indicating the presence of more light-demanding species (). Moreover, the diversity index value for mature trees (DBH > 30 cm) was higher in tribal and non-tribal forest (). Higher number and diversity of seedlings was recorded in the undisturbed forest. The Simpson's index was higher in the undisturbed forest; tribal and non-tribal sites had a lower index ().
Table 2. Vegetation characteristics of tree species of three areas of Kudremukh National Park, South India
Stem density of trees at sapling stage was low in tribal and non-tribal plots, while seedling density remained high. The RISQ value of the disturbance index () for mature trees, saplings and seedlings ranged from 1.16 to 1.19, with a low value in the undisturbed forest. The RISQ values in tribal and non-tribal forest ranged from 1.406 to 2.221, with the highest value for the sapling stage in non-tribal forest.
Stand quality
The percentage of primary species (shade species) such as M. dactyloides, Palaquium ellipticum, P. indicum and so on among tree, sapling and seedling stages was higher in the undisturbed forest (), while the percentage of light-demanding tree species such as M. peltata, Terminalia bellirica, M. philippensis and so on was higher in the tribal and non-tribal forest. In the undisturbed forest, primary tree species accounted for 81.86% and occupied a basal area of 86.83%, while such values did not exist in the tribal forest where primary tree species accounted for 49.5% and occupied a basal area of 84.63%.
Table 3. Contribution of primary, late secondary and early secondary trees to total basal area and number in Kudremukh National Park, South India
Discussion
It has been shown that human activities influence the quality of stands, microclimate, nutrient cycling and composition of forest species (Kappelle et al. Citation1995). One of the characteristic features of the humid tropical forest ecosystem is its high species richness (Parson and Cameron Citation1974). The number of species recorded here is comparable to that found in Swamy and Proctor (Citation1994) for species richness in the forest surrounding the KNP. The stem density for the mature trees in the undisturbed forest appears to be quite low (440 trees ha–1) compared with the study of Ghate et al. (Citation1998), which recorded a mean density of 419 trees ha–1 for the Western Ghats in a closed-canopy evergreen forest. However, the range of stem density recorded in the tribal and non-tribal plots for sapling and mature tree phases is closer to the observations of Pascal (Citation1988), Chandrashekara and Ramakrishnan (Citation1994), varying from 604 to 4566 trees ha−1, which is also more than in undisturbed plots. This is due to the lower canopy cover in the tribal and non-tribal plots, which causes higher light transmission, stimulating germination and rapid growth of pioneer species (Wales Citation1972; Levenson Citation1981; Ranney et al. Citation1981; Lopez de casenave et al. Citation1995; Jose et al. Citation1996).
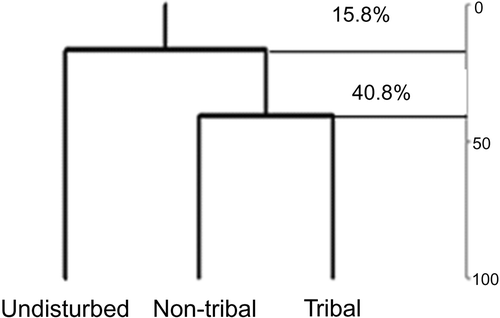
More variation in vegetation composition is observed between the undisturbed, non-tribal and tribal plots. Only 15.8% of species are similar between the undisturbed and non-tribal forest plots, where tribal and non-tribal forest plots had 40.8% species similarity (). The species similarity is less than 50% in the human-dominated plots, which implies that anthropogenic pressures had a serious impact on primary forest species, viz. C. travancorensis, Euonymus indicum, H. canarensis, P. ellipticum, P. indicum and Scheflera rostrata in the national park. These species were only found in undisturbed plots (). The density of saplings (20–30 cm DBH) was comparatively lower in the tribal and non-tribal inhabited areas than in the undisturbed forest due to the utilisation of saplings by inhabitants in these areas for fencing around settlements (protection from wildlife), roofing, preparing agricultural implements, firewood, green foliage (for Areca plantations and other cash crops) and so on.
Figure 1. Tree species similarity among sites in Kudremukh National Park, South India.
Although early- and late-secondary seedlings are regenerating in the undisturbed forest, these seedlings do not prevent native species regeneration because they do not establish into mature trees (Primack et al. Citation1985; Silva et al. Citation1995). There were more light-demanding species in the tribal forest. This could be attributed to vegetation changes following extensive canopy disturbance in the past and the recent past. A similar result was recorded in Kibale National Park, Uganda (Chapman and Chapman Citation1997), where creation of larger gaps facilitated colonisation by the light-demanding species.
The mature tree diversity index was higher in tribal and non-tribal forests than in the undisturbed forest. The presence of light-demanding species contributed to the increased diversity in the tribal and non-tribal forests. However, the values were lower than those recorded in tropical rain forest of Barro Colorado Island (Knight Citation1975) and the Silent Valley (Singh et al. Citation1981), India; but higher than those in Kerekatte forest near Sringeri, near the park (Swamy and Proctor Citation1994). Significantly higher diversity values were observed for the seedling phase in the undisturbed forest, indicating an increase in regeneration. The diversity of the seedling stage was comparatively lower in tribal and non-tribal forest, indicating low regeneration. This could be attributed to activities such as leaf litter collection, livestock grazing and so on which disturb the regeneration of primary forest tree species. Values of the dominance index of seedlings in tribal plots were higher than in non-tribal forests due to the establishment of light-demanding species through disturbance. In undisturbed forest, light-demanding species were comparatively less abundant in all stages.
The two experimental plots had a higher disturbance index, as they are located within a gap of 1–2 km around human settlements. Anthropogenic intervention has affected the regeneration strength of primary species (shade-loving, light-sensitive species) in the human-affected areas. A higher RISQ value for saplings and seedlings than mature trees indicates repeated disturbance or failure of shade-tolerant species to establish (Chandrashekara and Sankar Citation1998). Due to the canopy openness and more light availability the late-secondary saplings and seedlings are more common in tribal and non-tribal plots, resulting in a loss of the regeneration capacity of primary forest species. Hence it is recommended to plant primary forest species such as Garcinia gummi-gutta, Calophyllum apetalum, M. dactyloides and so on in degraded forest fringes close to tribal and non-tribal settlements. It is also important to undertake awareness programmes among local people on the importance of conserving this biodiversity hotspot and restoring forest fringe areas in the protected areas. The Forest and Wildlife Departments, along with other government departments, should implement programmes such as providing energy- efficient cooking devices, sustainable harvesting of forest products and so on to reduce pressure on the protected area, which may have a direct ecological impact on the forest ecosystem in the future.
Conclusion
This study highlights the distribution of plant resources in the dynamic landscapes of the Western Ghats region. The forest ecosystem in both settlement areas supported higher plant diversity than the undisturbed forest, with less shade-loving primary forest species. This clearly indicates that human settlements close to rain forests directly impact regeneration of primary forest species, resulting in changes in the composition of the forest. Our observations will help forest functionaries to prepare site-specific restoration plans. Primary forest species, such as M. dactyloides, P. ellipticum, G. gummi-gutta and P. indicum, could be used in the restoration process. These species help to maintain forest ecosystem stability as well as improving the provision of forest ecosystem services.
Acknowledgements
The authors thank the Karnataka Forest Department for granting permission to carry out the work in Kudremukh National Park and their invaluable help in site selection. The valuable comments and suggestions from anonymous reviewers are gratefully acknowledged.
References
- Bawa , KS , Setty , S and Joseph , G. 2007 . Poverty, biodiversity and institutions in forest-agriculture ecotones of the Western Ghats and Eastern Himalaya ranges of India . Agric Ecosyst Environ , 121 : 287 – 295 .
- Bourgeon , G. 1989 . Explanatory booklet on the reconnaissance soil map of forest area , Pondicherry , India : French Institute .
- Chandrashekaran , C. 1960 . “ Forest types of Kerala state ” . In Special report submitted for Diploma in Forestry , Dehradun , , India : New Forest .
- Chandrasekhara , UM. Ramakrishna Index of stand quality (RISQ): an indicator for the level of forest disturbance . Proceedings of the 10th Kerala Science Congress State Committee on Science . Thiruvananthapuram , India . Edited by: Damodaran , AD . pp. 398 – 400 . Technology and Environment .
- Chandrashekara , UM and Ramakrishnan , PS. 1994 . Vegetation and gap dynamics of tropical wet evergreen forests in the Western Ghats of Kerala, India . J Trop Ecol , 10 : 337 – 354 .
- Chandrashekara , UM and Sankar , S. 1998 . Ecology and management of sacred groves in Kerala, India . For Ecol Manage , 112 : 165 – 177 .
- Chapman , CA and Chapman , LJ. 1997 . Forest regeneration in logged and unlogged forests of Kibale National Park, Uganda . Biotropica , 29 ( 4 ) : 396 – 412 .
- Gadgil , M and Subhas Chandran , ND . 1988 . “ On the history of Uttara Kannada forests ” . In Changing tropical forests , Edited by: Dargavel , J , Dixon , K and Semple , N . Canberra , Australia : Centre for Resource and Environmental Studies .
- Gamble , JS. 1928 . Flora of Presidency of Madras , Vol. 1–3 , London , UK : Adlard and Son Ltd .
- Ghate , U , Joshi , NV and Gadgil , M. 1998 . On the patterns of tree diversity in the Western Ghats of India . Curr Sci , 75 : 594 – 603 .
- Hill , DA , Fasham , M , Tucker , G , Shewry , M and Shaw , P. 2005 . Handbook of biodiversity methods: survey, evaluation and monitoring , 219 – 222 . Cambridge , UK : Cambridge University Press .
- Hussain , SA. 2003 . Kudremukh National Park, Karnataka: a profile and a strategy for the future . J Bombay Nat Hist Soc , 100 ( 2&3 ) : 202 – 213 .
- Jose , S , Gillespiie , AR , George , SJ and Kumar , BM. 1996 . Vegetation responses along edge-to-interior gradients in a high altitude tropical forest in peninsular India . For Ecol Manage , 87 : 51 – 61 .
- Kappelle , M , De Vries , RAJ and Kennis , PAF. 1995 . Changes in diversity along a successional gradient in a Costa Rican . Biodivers Conserv , 4 : 10 – 34 .
- Kershaw , KA. 1973 . Quantitative and dynamic plant ecology , 286 London , UK : Edward Arnold .
- Kimmins , H. 1992 . Balancing acts: environmental issues in forestry , Vancouver , BC : University of British Columbia Press .
- Knight , DH. 1975 . A phytosociological analysis of species-rich tropical forest: Barro Colorado Island, Panama . Ecol Monogr , 45 : 259 – 284 .
- Krishnamurthy , SV , Reddy , AHM and Gururaja , KV. 2001 . A new species of frog in the genus Nyctibatrachus (Anura: Ranidae) from Western Ghats, India . Curr Sci , 80 ( 7 ) : 887 – 891 .
- Levenson , H. 1981 . “ Woodlots as biogeographic islands in South-eastern Wisconsin ” . In Forest islands dynamics in man-dominated landscape , Edited by: Burgess , R and Sharpe , D . 13 – 39 . New York , NY : Springer .
- Lopez de casenave , J , Pelotto , JP and Protomastro , J. 1995 . Edge-interior differences in vegetation structure and composition in a Chaco semi-arid forest, Argentina . For Ecol Manage , 72 : 61 – 69 .
- Parson , RF and Cameron , DS. 1974 . Maximum plant species diversity in terrestrial communities . Biotropica , 6 : 202 – 203 .
- Parthasarathy , N. 1999 . Tree diversity and distribution in undisturbed and human impacted sites of tropical wet evergreen forest in Southern Western Ghats, India . Biodivers Conserv , 8 : 1365 – 1381 .
- Pascal , JP. 1988 . Wet evergreen forest of the Western Ghats of India , Pondicherry , India : French Institute .
- Primack , RB , Ashton , PS , Chai , P and Lee , HS. 1985 . Growth rates and population structure of Moraceae trees in Sarawak, East Malaysia . Ecology , 66 ( 2 ) : 577 – 588 .
- Rai , SN. Gap regeneration in wet evergreen forest of Karnataka . Karnataka (India): Karnataka Forest Department. Karnataka Forest Department Research Report, KFD-2 . 1979 .
- Rai , SN and Proctor , J. 1986 . Ecological studies on four rainforest in Karnataka, India. Environmental, structure, floristics and biomass . J Ecol , 74 : 439 – 454 .
- Ranney , J , Bruner , M and Levenson , J. 1981 . “ The importance of edge in the structure and dynamics of forest islands ” . In Forest islands dynamics in man-dominated landscape , Edited by: Burgess , R and Sharpe , D . 67 – 75 . New York , NY : Springer .
- Shannon , CE and Wiener , W. 1963 . The mathematical theory of communication , Urbana , IL : University of Illinois Press .
- Silva , JMN , De Carvallho , JOP , Costa , DHM , De Oliveira , LC , De Almeida , BF , Lopes , JCA , Skovsgaard , JP and Van Clay , JK. 1995 . Growth and yields of a tropical rainforest in the Brazilian Amazon thirteen years, after logging. For Ecol Manage , 71 : 267 – 274 .
- Simpson , EH. 1949 . Measurement of diversity . Nature , 163 : 688
- Singh , JS , Singh , SP , Saxena , AK and Rawat , YS. 1981 . “ The Silent Valley forest ecosystem and possible impact of proposed hydroelectric project ” . In Reports on the Silent valley study , Nainital (India): Ecology Research Circle, Kumaun University .
- Swamy , HR and Proctor , J. 1994 . Rain forests and their soils in the Sringeri area of the Indian Western Ghats . Global Ecol Biodivers Lett , 4 : 140 – 150 .
- Venkatesh Nayak , SN , Ramachandra Swamy , H , Nagaraj , BC , Rao , U and Chandrashekara , UM. 2000 . Farmers attitude towards sustainable management of Soppina Betta forests in Sringeri area of the Western Ghats, South India . For Ecol Manage , 132 : 223 – 241 .
- Wales , B. 1972 . Vegetation analysis of north and south edges in a mature oak-hickory forest . Ecol Monogr , 42 ( 4 ) : 451 – 471 .
- Whitmore , TC. 1989 . Changes over twenty-one years in the Kolombangara rain forests . J Ecol , 77 : 469 – 483 .
- Whitmore , T and Sayer , J. 1992 . Tropical rainforests of the far east , 2nd , Oxford , UK : Clarendon Press .
- WRI, IUCN, UNEP] World Resources Institute, International Union for Conservation of Nature, United Nations Environment Programme . 1992 . Global biodiversity strategy: guidelines for action to save, study, and use earth's biotic wealth sustainability and equitably , Washington , DC : WRI, IUCN, UNEP .