Abstract
Herbicide application on permanent grassland to reduce weeds and improve forage quality is common agricultural practice. However, it still remains unclear how long it takes for the herbicide-disturbed swards to recover in terms of yield and forage quality. In a removal experiment in the Solling Uplands (Germany), the sward composition of permanent grassland had been manipulated by herbicides in order to obtain either relatively pure grass swards or swards with comparatively large amounts of forbs and legumes, in addition to untreated control swards. The short-term resilience of these sward types was examined under a gradient of management intensity regulated by both cutting regime and fertilizer supply. In the next growing season, the yield did not differ among any of the three sward types regardless of the management regime. All disturbed swards showed a complete recovery in terms of biomass. Yield was only influenced by functional sward characteristics across all disturbance treatments; the growth form of the dominant species determined the yield in fertilized plots. For the variation in forage quality (crude protein, water-soluble carbohydrates and fibre content), the functional group identity of the remaining vegetation was important, but management had a much larger influence than vegetation.
1. Introduction
Herbicide application on grassland is a common agricultural management practice. It aims at a reduction of less valuable forage species in the grass sward and consequently achieving higher yields and improved fodder values. These herbicide treatments are often combined with over-sowing of desired species (Frame and Laidlaw Citation2011). However, their establishment and effectiveness in improving the sward performance usually require 12 years given that weather and management are optimal for the establishment of grass seedlings (Opitz v. Boberfeld Citation1994). On grassland, herbicides have usually been applied against certain groups of species, e.g. broadleaved or annual dicots (Lewis and Hopkins Citation2000). Due to environmental concerns, herbicide use has decreased since the 1990s (Lewis and Hopkins Citation2000), but remains common practice: in Britain, for example, up to 7% of grassland is treated annually (Frame and Laidlaw Citation2011).
Reducing weeds means reducing biodiversity at least in the short term (Virágh Citation1987), no matter whether dicot weeds or monocot weeds are targeted. According to the results of biodiversity investigations, where predominantly positive relationships between biodiversity and many important ecosystem functions were found in field experiments (Quijas et al. Citation2010), this loss of biodiversity could have negative consequences at least for parts of the grassland ecosystem. However, most of these experiments lack direct applicability to agricultural systems (Isselstein Citation2005; Wrage et al. Citation2011). If the positive relationship is indeed dominating, why do farmers ‘improve’ their grasslands by herbicide application?
The impact of a disturbance like herbicide treatment on a grassland plant community mainly depends on the disturbance's duration and intensity and on the resistance and resilience of the original vegetation (Touzard and Clement Citation2001; Renne and Tracy Citation2007). The original biodiversity and the traits (mainly in terms of re-colonization abilities) of the remaining species are important for the vegetation's resilience (Virágh Citation1987; Lavorel Citation1999; Roxburgh and Wilson Citation2000; Symstad and Tilman Citation2001; McLaren and Turkington Citation2010). In addition, the environmental conditions at the disturbed site play an important role for the speed and completeness of the recovery of different vegetation characteristics like cover, biomass and composition (Fowler Citation1981; Virágh Citation1989; Symstad and Tilman Citation2001; Touzard and Clement Citation2001; Renne and Tracy Citation2007; Slocum and Mendelssohn Citation2008).
In diverse grassland, only a severe loss in species numbers is expected to completely disrupt the grassland ecosystems, as ecosystem processes seem to saturate at small (around five) numbers of species (Roy Citation2001; Wrage et al. Citation2011). Compared to ley farming systems, even intensively managed permanent grasslands host higher species numbers (Dykmans et al. Citation1999; Klimek et al. Citation2007; Pywell et al. Citation2007). Consequently, herbicide application as a management method should not have detrimental effects on agriculturally important ecosystem services such as productivity.
In contrast to productivity, forage quality should react to changes in diversity more severely: it is supposed to improve if less palatable weeds are gone. Thus, besides the main drivers of quality changes, maturity and nutrient status (Bruinenberg et al. Citation2002), forage quality is also influenced by biodiversity and species composition (White et al. Citation2004). The presence of forbs and legumes can improve forage quality in terms of crude protein (CP) content as Seng et al. (Citation2008) and Seip et al. (Citation2011) found when comparing pure grass swards (made up either from pure Lolium perenne or different grass species) with mixed permanent grassland. Some dicot species contain higher amounts of Mg and Na than grasses (Wilman and Riley Citation1993).
In this article, we report first results of the grassland management experiment (GrassMan), an experiment on managed permanent grassland manipulated once by herbicides to vary species composition and functional diversity in the swards (Petersen et al. Citation2012). To analyse the recovery process under different environmental conditions, the resulting swards were subjected to four different combinations of fertilization and cutting regimes. Here, we examined the resilience of forage yield and quality focussing on the short-term effects of the herbicide disturbance, that is, whether the manipulated swards recovered until the next growing season or whether yield and quality were changed. We hypothesized that:
| • | herbicide application and strong reduction of any functional group does not affect the yield in the following growing season; | ||||
| • | the remaining vegetation's ability to compensate the losses depends on the traits of the remaining species and the management and | ||||
| • | any substantial change in the functional composition (distribution of functional groups and species composition) affects forage quality. | ||||
2. Methods
2.1. Site and experimental design
The experimental site is located between Silberborn and Neuhaus, in the Solling Uplands, Germany (51° 44′ 53″ N, 9° 32′ 42″ E, 490 m a.s.l.) on long-term permanent grassland managed by the experimental farm of the University of Goettingen at Relliehausen since 1966. The mean annual temperature is 6.9°C and mean annual rainfall amounts to 1028 mm (Deutscher Wetterdienst, “German Meteorological Service” 1960–1991, station Silberborn-Holzminden, 440 m a.s.l.). The soil of the experimental area was determined as a shallow (40–60 cm), stony haplic Cambisol (Keuter et al. Citation2012) on middle Bunter (Triassic sandstone) (Hedemann Citation1950). The grassland had been used as summer pasture for cattle and had received an annual fertilizer amount of 80 kg N ha−1 yr−1 until two years prior to the start of the experiment. Its vegetation was classified as a nutrient poor, moderately wet Lolio-Cynosuretum with high abundances of Festuca rubra and Agrostis capillaris.
The GrassMan experiment was set up in the summer of 2008. It is a three-factor experiment incorporating the factors sward type, utilization and fertilization (). It has got six replicates resulting in 72 experimental plots, each 15 m by 15 m large. To account for potential spatial heterogeneity due to the location on a shallow slope bordering a forest on the upper part, the experimental layout was a Latin rectangle design with the six replicates (blocks) arranged in 6 rows and 12 columns, two columns forming one block.
Table 1. Experimental factors and treatment levels of the GrassMan experiment
Since we wanted to test disturbances in both directions, i.e. to obtain dicot-reduced (–Dic) and monocot-reduced (–Mon) swards on top of the untreated control sward (Co), herbicides against dicots (active ingredients Fluoroxypyr + Triclopyr and Mecoprop-P; 3 l ha−1 each) and monocots (Clethodim 0.5 l ha−1) were applied each on a third of the experimental plots. The dead plant material was not removed, to mirror common practice in managed grasslands. The herbicide application followed 4 weeks after the first harvest of the whole area, at the end of July 2008. In spring 2009, three initially distinct sward types had developed on the experimental area. They differed in species numbers (7–16 per m2), species composition and functional group composition (Petersen et al. Citation2012). Starting with a grass:forb:legume ratio of 76:22:2 (June 2008), a gradient ranging from 39:52:9 (–Mon) up to 93:7:0 (–Dic) was generated. The fertilized plots (acronym NPK) received 50 kg N ha−1 in 2008 and the plots belonging to the three times cut regime were harvested a second time at the end of October that year. The complete management with NPK fertilization (90 kg N ha−1 at the end of April, the remaining N, P and K at the end of May) and scheduled harvests did not start until spring 2009.
2.2. Measurements
Species composition and functional diversity were monitored by means of vegetation relevés on permanent sub-plots where the dry matter (DM) yield proportions of each species were estimated according to Klapp and Stählin (Citation1936). Two 9 m2 sub-plots per plot, each with an additional 1 m2 quadrant in its centre for different scales of diversity measures, were recorded twice a year at the beginning of May before the first and in mid-August 4 weeks after the second harvest. The yield proportions of the species occurring in at least one-third of the plots with proportions larger than 5% were included in the statistical analyses as factor ‘species identity’ (ID). Additionally, the mass proportions of grasses, forbs and legumes and the amount of dead plant material were determined by the sorting of sub-samples of each plot taken at harvest. Another functional grouping comprised the three functional groups, grasses, forbs and legumes, comparable to the classification used by Roscher et al. (Citation2004) with further subdivisions into species with tall and with small stature. Besides growth height and stature, the criterion for this subdivision into six groups was also the sensitivity to defoliation. These criteria closely follow the classification of grasses into understory grasses and tall meadow grasses according to Klapp (Citation1954) and Klapp and Opitz von Boberfeld (Citation2006). This classification was adopted for legumes and forbs. Relative growth rates of single species according to Grime et al. (Citation1988) were not included in this classification as almost all common species had the same growth rate (1–1.4 g g−1 week−1). The only fast growing species, Urtica dioica and Holcus lanatus, were only present in small amounts on the experimental area. All traits needed for this classification, as well as information on the main propagation type of the species (vegetative vs. by seeds), were acquired from Schrader and Kalthofen (Citation1974), Grime et al. (Citation1988) and the database BiolFlor Version 1.1 (Klotz et al. Citation2002).
The DM yield per hectare was projected from the average yield of two 1.50 m × 15 m stripes per plot harvested with a Haldrup® forage combine harvester (cutting height 7 cm). For forage quality analyses, mixed herbage samples per harvested plot were taken and dried on the same day at 60°C (for 48 hours) in a forced-air oven. The material was then ground to pass through a 1-mm screen. We estimated CP, water-soluble carbohydrates (WSCs), neutral detergent fibre (NDF) and acid detergent fibre (ADF) contents by near-infrared reflectance spectroscopy (NIRS). The spectra were analysed using the large dataset of calibration samples from different kinds of grasslands by the Institute VDLUFA Qualitätssicherung NIRS GmbH, Kassel, Germany (Tillmann Citation2010). As described in Kesting et al. (Citation2009), samples showing a lack of accuracy (H value exceeding 3) were excluded from further statistical analyses (1 out of 144 samples). For the calculation of the energy content (metabolizable energy, ME) of the forages, we determined the dry ash content by incinerating samples of the ground material in a muffle furnace (550°C) for 5 hours. ME was calculated according to the equation by Gesellschaft für Ernährungsphysiologie (‘Society for Nutrition Physiology’, Citation2008) incorporating crude ash, CP, crude fat and ADF contents among others.
2.3. Statistical analyses
Univariate statistical analyses were conducted with R (version 2.12.2) (R Development Core Team Citation2011). ANOVAS, linear contrasts and linear models (lm) were used to compare the importance of treatment factors and functional diversity influencing the measured parameters as well as their means. For the comparison of means of all treatments, Tukey's ‘Honest Significant Difference’ method (Miller Citation1981; Yandell Citation1997) was used with a confidence level of 0.95. To take into account the environmental heterogeneity and its influence on forage yield and quality, the rows and blocks of the experimental area were added to the models first of all factors. The best fitting models including only row, block and the significant independent predictive variables and interactions were obtained by the comparison of the Akaike Information Criteria of the full and the reduced models (Zuur et al. Citation2009). The collinearity of continuous variables like forb and grass content, or individual species yield proportions, was checked beforehand in a special version of the pairs plot following Zuur et al. (Citation2009). If vegetation characteristics like species number, proportion of tall species, amounts of legumes and so on were included, the factor sward type was excluded from the models since it was strongly correlated with all the vegetation characteristics. To obtain homoscedasticity and normal distribution of the residuals, the data were fitted in general least square (gls) models including the ‘varIdent’ variance structure (different variance per factor level, ‘nlme’ package Pinheiro et al. Citation2009) if necessary. Additionally, a part of the data were transformed according to the results of the boxcox analysis (MASS package in R; Venables and Ripley Citation2002) as indicated in the results. To analyse the influence of management and vegetation characteristics on forage quality as a whole, all NIRS-measured quality parameters were included as dependent variables in an ordination (they took the place of the ‘species’ in ordination of vegetation relevés), whereas management (fertilization and utilization frequency) and vegetation characteristics (sward type, proportions of functional groups or tall and small grasses, forbs and legumes and species ID) represented the independent, ‘environmental’ variables. For statistics, vegetation characteristics of the three-cut regime were averaged first over all harvest dates. Since the spatial distribution of several species depended on rows and blocks of the Latin rectangle, these two spatial variables were excluded from direct ordinations. The program Canoco for Windows version 4.5 (ter Braak and Šmilauer Citation1997–2004) was used for redundancy analysis (RDA, constrained ordination) including Monte Carlo permutation tests for assessing importance and significance of the environmental variables (ter Braak and Šmilauer Citation2002). We used the variance partitioning procedure (Borcard et al. Citation1992; Vandvik and Birks Citation2002) to quantify the proportions of variance in quality explained by management, vegetation characteristics or environmental influences (date, percentage of open soil and position within the Latin rectangle). The quality of the constrained ordination was judged by comparison of its ordination diagrams with those from an unconstrained ordination (here a principal component analysis, PCA) and the amount of residual variance in the RDA.
3. Results
3.1. Yields
The yield proportions of single species as well as their frequency in the vegetation relevés were affected by the herbicide application. Particularly the dicot species became less frequent () in the –Dic swards; the grasses were mainly reduced in yield proportions, and their frequency hardly changed. Several species showed seasonal fluctuations. For example, Poa trivialis, Poa pratensis and Cardamine pratensis decreased later in the year, while Dactylis glomerata and Taraxacum Sec. Ruderalia had higher yield proportions in August. The herbicides against dicots did not completely eliminate all forbs, but were very effective in removing legumes. The most common forbs were mainly reduced in yield proportion, not in frequency. The same applied to the grasses. Red fescue (F. rubra) was hardly affected by the herbicide against monocots; it even slightly increased its frequency. The available space was colonized mainly by species belonging to the functional group spared by the herbicides. In the –Mon swards, seed propagating as well as vegetatively spreading dicots gained more space. In the –Dic swards, however, the seed propagating tuft grasses could not expand significantly (data not shown) and still had the same yield proportion as in the control plots.
Table 2. Relative change of degree of presence of the 20 most frequent species in 9 m2 before (June 2008) and after (May, August 2009) the herbicide application within the three sward types
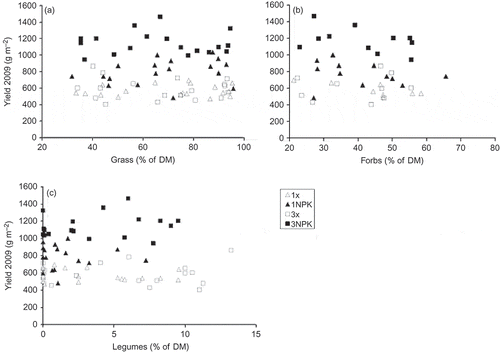
The 12 experimental treatments resulted in widely spread annual yields ranging from an average of 550 g m−2 in the –Mon3x up to 1220 g m−2 in the Co3NPK treatment. Within none of the four management categories, the yields differed significantly among the three sward types. In the plots without fertilization cut three times, the –Mon-swards tended to have the largest yields, but the yield increase due to fertilization was significantly smaller than that in the control swards (interaction fertilization × sward type –Mon, P = 0.019). In the fertilized swards cut only once, the –Dic-swards tended to produce more forage than the other sward types. The untreated control swards gained most from the intensive treatment with fertilization and three cuts. The management in terms of fertilization and utilization frequency was mainly responsible for yield differences in overall yields in 2009 (). However, the influence of the experimental factors on single yields was not constant, as shown in During the course of the year 2009, fertilization gained importance, whereas the sward type did not explain significant amounts of yield variation apart from the very first cut after herbicide application in October 2008. We did not find any significant relationship between the proportions of grasses, forbs and legumes and yield within the four management categories ((a)–(c)). The variance explained by the lm did not differ regardless of whether one of the functional groups or the sward type was included.
Table 3. Importance of the experimental factors in explaining the differences in annual yield 2009
Table 4. Proportion of variation in single and total yields 2008–2009 explained by the experimental factors (%)
Figure 1. Relationships between total yield 2009 and proportions of grasses (a), forbs (b) and legumes (c) under the four management regimes (legend: middle right). None of the correlations between yield and independent variable within one management category were significant. For abbreviations of experimental treatments please refer to Table 1.
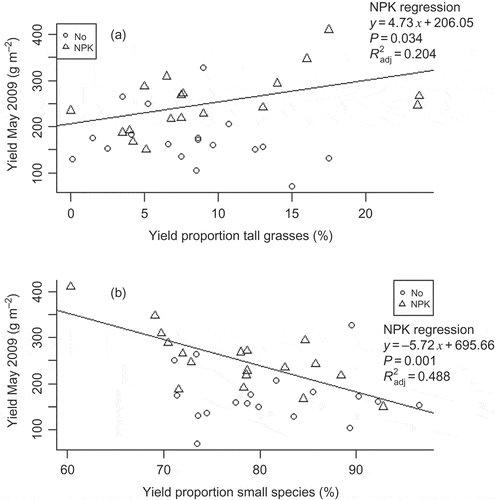
However, using the finer division into tall and small grasses, forbs and legumes, we found significant effects on yield in May 2009 (), when we had recorded both vegetation and yield data. A larger proportion of tall grasses () or tall species altogether (data not shown) increased yields significantly in May 2009 – yet, only in the fertilized plots. The number of tall species or grasses was irrelevant. The higher the number of small species present, the less forage was harvested on NPK-plots. This trend was also visible in the yield × species ID correlation (data not shown). Achillea millefolium (classified as a tall forb) showed a tendency to increase total yield in fertilized plots, whereas A. capillaris (a small grass) seemed to decrease total yield in fertilized plots when it was present in large amounts. However, the correlations were not significant at the 5% level and none of the other frequent species showed any visible correlation with yield at all. The variability in yield at the experimental area was best explained by a lm containing the amounts of tall grasses, small species, the factor fertilization and all interaction terms (R 2 adj = 0.529, P < 0.001). Still, fertilization alone (excluding interactions with vegetation) accounted for 21.3% of all variation in the yield.
Figure 2. Yield May 2009 dependent on proportion of tall grasses (a) and proportion of small species (b) grouped by fertilization level (with regression lines). The corresponding linear model includes a significant interaction between nutrients and tall grasses (P = 0.032) and nutrients and small species (P = 0.017), response variable untransformed.
3.2. Forage quality
The energy content of the forages at the first and third harvests (May: 11.6 ± 0.2 MJ (kg DM)−1, September: 11.6 ± 0.2 MJ (kg DM)−1) was not influenced by any of the experimental factors. At the second harvest in July, the cutting regime (1-cut regime, 10.6 ± 0.2 MJ (kg DM)−1; 3-cut regime, 11.1 ± 0.1 MJ (kg DM)−1) was responsible for most of the variation in energy content (ANOVA based on gls including one variance term per level of the factor utilization, P < 0.001, data not shown). However, in unfertilized plots, the forage samples from the –Mon-swards had significantly lower energy contents than those from the other two sward types (linear contrasts in gls model including one variance term per utilization type, P = 0.005). This distinction was not visible in fertilized plots (significant interaction sward type –Mon × fertilization, P = 0.014).
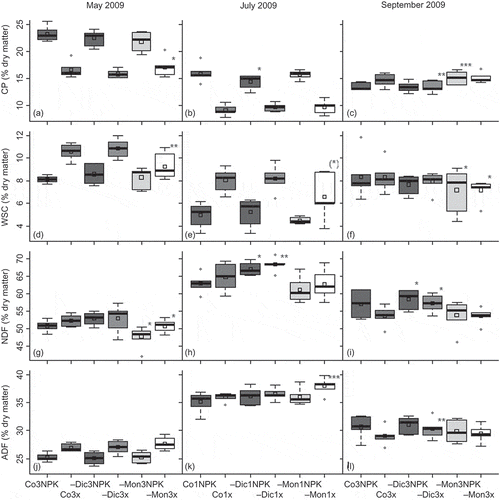
For four analysed forage quality parameters (CP, WSC, NDF and ADF), we found significant sward effects at different harvest dates (). However, these effects were not consistent across the fertilization levels; we found significant interactions between sward type and fertilization ((a), (c), (d)). When comparing the forage quality of the first growth cycles (i.e. forages cut in May and the ones cut for the first time in July), apart from a general loss of quality due to senescence of the sward, we detected a significant time × sward interaction. From May to July, the swards rich in forbs increased in ADF contents significantly more (P = 0.043) than the other swards.
Figure 3. Box and whisker plots showing the four quartiles, the median (calculation of outliers by the default method in R) and the means, denoted by small squares, of selected forage quality characteristics in the six different sward type × fertilization level combinations at each of the three harvests. Second cut (July) of the three cut regime not presented. Asterisks denote significant differences from the control sward of the same fertilization level (***P < 0.001, **P < 0.01, *P < 0.05, (*)P = 0.0506). Linear contrasts in linear models; either models with different variances per fertilization level (graphs (c), (k)) or sward type (graph (f)) or response variable either square root (graph (h)) or not transformed (all others). For abbreviations of experimental treatments please refer to Table 1.
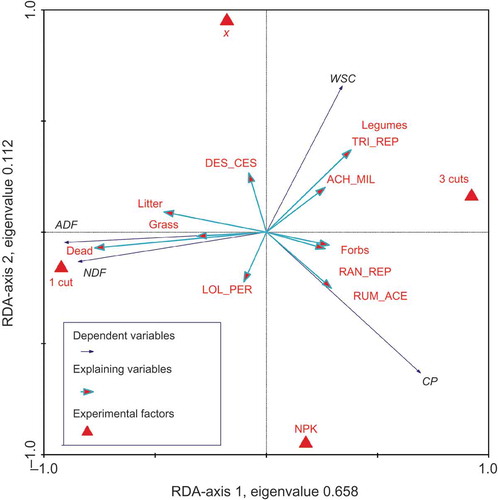
The unconstrained ordination (PCA) of the quality parameters showed the same main gradients determining forage quality as the constrained ordination (RDA) (). The environmental variables selected for RDA explained 81.7% of the total variation of the forage quality parameters in 2009. The cutting regime, or rather the age of the swards at harvest, explained most of the variation (57%, P < 0.010, Monte Carlo permutation test). The variation in sugar and protein contents made up the second gradient of forage quality variation. It was mainly determined by the fertilization regime, which explained 12% of the total variation (P < 0.010). After accounting for management influences, only forbs, the three species Rumex acetosa, Trifolium repens and A. millefolium, the amount of tall forbs or the sward type –Mon (not included in the ordination diagram in ) explained significant (P < 0.050, Monte Carlo permutation test) conditional proportions of the remaining variance if added to the ordination model as separate groups. The remaining species and functional groups shown in only accounted for significant parts of the remaining variation (after extracting the influences of management) if added as single extra independent variables. The variable dead material in forage samples was the only one that did not explain any extra variation, since it was highly correlated with cutting regime. The results of the variance partitioning emphasized the influence of management on forage quality. Just a small part (up to 8.6%) of variation in forage quality was solely due to sward characteristics.
Figure 4. Ordination diagram of a redundancy analysis (RDA) with forage quality characteristics (contents of CP, WSC, ADF, NDF in 2009, untransformed) as dependent variables and the experimental factors utilization and fertilization (cf. ) and vegetation characteristics (instead of sward types) as explaining variables. Quality characteristics of the three-cut variant are averaged across harvest dates. Explaining variables with a correlation coefficient with the first and second ordination axis <–0.2 and >0.2 are included.
4. Discussion
4.1. Yields
The main objective of this study was to investigate the effects of different herbicides on forage yield and quality in managed permanent grassland under different management categories. We hardly found any influences of disturbance by herbicides, of functional diversity or of botanical composition on yield in this experiment: most of the yield variation was due to the management category, supporting our first hypothesis.
The sward effect found in October 2008 was the only sign of the previous disturbance, since both treated swards had lower yields than the control sward, indicating the ongoing recovery process after the herbicide application. By May 2009, however, in the following growing season, the treated swards had fully recovered in terms of biomass production: their yields did not differ from those of the control swards (Petersen et al. Citation2012). In other removal experiments (e.g. Symstad and Tilman Citation2001; McLaren and Turkington Citation2010; Šmilauer and Šmilauerová Citation2012), it took a year or more for the remaining vegetation to reach the biomass production of the control plots after the removal had taken place.
This may be due to the moderate removal of biomass and the intermediate species number in the different sward types analysed here. Only about 20% of total biomass (the yield proportion of dicots before herbicide application) was removed in the –Dic plots. Due to the insensitivity of some grasses, the biomass removal in the –Mon plots cannot be estimated from their pre-treatment proportions. In other removal experiments, the loss of biomass due to the disturbance was higher, e.g. 30–50% depending on the distribution of the functional groups (Symstad and Tilman Citation2001; McLaren and Turkington Citation2010) or around 50% (Virágh Citation1987; Šmilauer and Šmilauerová Citation2012). Consequently, biomass production levels took longer to recover, especially if the remaining vegetation had recruitment limitations as encountered by Symstad and Tilman (Citation2001); Roxburgh and Wilson (Citation2000) found a recovery rate comparable to ours, when they removed all graminoids (approximately 50–60% of total biomass) by herbicide in a dicot-dominated lawn. Six weeks after the removal, the cover values they recorded in their herbicide treatment were similar to those of the control plots.
After the disturbance by herbicides in our experiment, sufficient species remained to compensate for the losses, no matter which groups of species were missing. As Schwartz et al. (Citation2000) and Waide et al. (Citation1999) pointed out, most of the ecosystem processes are saturating at low species numbers; Roy (Citation2001) summarized that, in many studies, five plant species were enough to reach 90% of the productivity effect. Since the remaining vegetation in our experiment consisted of at least seven species per m2, the ecosystem functions, in this case the yield, should not have shown a lower rate of functioning. In accordance with this, we did not find any effects, or the existing effects were too small compared to the variability in our treatments as is often the case in experiments with no controlled environments (Balvanera et al. Citation2006).
Of crucial importance for productivity are the traits of the remaining species after a disturbance. As reported in a review on observational studies (Jiang et al. Citation2009), the abundance of the dominant species along with their special traits (here in all swards A. capillaris and F. rubra as low- to medium-yielding grasses) may influence yields more than a shifted monocot:dicot ratio. We likewise did not find any influence of dicot or monocot proportions on yield, only the presence of productive bunch grasses made a difference to yields in the first cut. This is consistent with the findings of Mokany et al. (Citation2008), who showed that it was not the ‘diversity’ (Tilman Citation1997) but rather the ‘mass ratio’ (Grime Citation1998) hypothesis that better explained ecosystem process variation in natural communities. Thus, the more of the taller grasses were present in the plots, the higher yields we got in the first cut, underlining the importance of special traits as hypothesized.
The effects of functional diversity on ecosystem functions also depend on management (e.g. Bernhardt-Römermann et al. Citation2011). In line with this, we only found effects of special traits (growth characteristics) in fertilized plots, i.e. dependent on agricultural management as we had anticipated in our second hypothesis. Silletti et al. (Citation2004) likewise found different growth responses of one of the remaining species after neighbour removal to depend on the nutrient status of the soil. In our experiment, the faster-growing tall grasses gained more biomass from the fertilization than the smaller species.
Not only productivity related traits, but also dispersal abilities are of importance. Since the tall tuft grasses exhibited poor re-colonization abilities, they could not increase their yield proportions in the –Dic plots; so their influence on total yield did not lead to any differences between control and –Dic-swards. In the disturbed plots, the traits of the fastest-colonizing (A. capillaris, Ranunculus repens and R. acetosa) or most resistant (F. rubra in the –Mon swards) species dominated. Since these are not the highest yielding species, the treated plots did not have higher yields than the control.
4.2. Forage quality
In line with the results for yield, the influence of sward characteristics on special forage quality characteristics depended on management regime. We only found significantly reduced energy contents in unfertilized –Mon swards cut once in July. Fertilization did not increase energy contents. Since the energy content of forages depends on all cell contents (protein, carbohydrates and fats) and the fibre content determining their digestibility (Voigtländer and Jacob Citation1987), the equal energy contents in fertilized versus unfertilized swards can be explained by their contrasting contents of CP and WSC in May and July (, (b), (d), (e)). Low CP contents in unfertilized swards were compensated for by higher WSC proportions and vice versa. In September, CP and WSC did not differ among the fertilization regimes, leading to equal energy contents again.
The senescence of the swards combined with higher amounts of structural tissue (= higher amounts of dead plant material, see ) reduced forage quality in terms of digestibility (Duru et al. Citation2008) in the swards of the 1-cut regime. The –Mon-swards contained less energy due to increased levels of ADF and decreased levels of WSC (). This is consistent with Lopez et al. (Citation1991), Bruinenberg et al. (Citation2004) and Andueza et al. (Citation2010) who also reported increased ADF values in forbs compared with grasses due to higher lignin contents.
For the summer re-growth, Bruinenberg et al. (Citation2002) reported higher digestibility in forb species than in grass species. This is comparable with our results. The –Dic-swards had higher NDF and ADF contents than the –Mon-swards in the second (data not shown) and the third cuts ((i), (l)), whereas the CP contents were highest in the –Mon-swards. High CP contents in forbs were similarly found by Wilman and Riley (Citation1993); Bruinenberg et al. (Citation2004) and Andueza et al. (Citation2010). These study results clearly support our third hypothesis.
However, there may be other crucial factors influencing forage quality far more than sward composition does. Especially the cutting date along with the phenological development of the swards can be responsible for the largest changes in quality (van Soest et al. Citation1978; Bruinenberg et al. Citation2002; Čop et al. Citation2009; Bochi-Brum et al. Citation2011). This is due to a declining leaf:stem ratio when the grasses start their generative growth (Duru et al. Citation2008). Further, the accumulated biomass needs to be supported by structural tissues. Contents of soluble carbohydrates, mainly found in the leaf tissue, are in decline, reducing digestibility. This is also mirrored in our data. The forage of the late first cut in July had the highest NDF and ADF contents ((h), (k)). The high ADF contents in the –Mon1x swards could be due to higher proportions of fruiting Rumex acetosa, R. repens and red fescue. The other sward types hosted larger amounts of A. capillaris still in bloom.
In the redundancy analysis, most variation in forage quality of the whole year 2009 was explained by management, that is to say, fertilization and cutting regime. As also reported by Čop et al. (Citation2009), who analysed the quality of forages by means of ordination as well, the main gradient in our redundancy analysis was the contrast in fibre contents induced by different cutting dates. The CP content as the second largest gradient showed a much smaller variability. Just in the third ordination axis, the sward characteristics were the determining factors but they hardly explained any (eigenvalue: 0.046) of the remaining variation in forage quality.
To sum up, the swards showed a high resilience towards a herbicide treatment in terms of yield; they had recovered until the following growing season. Yield was mainly influenced by fertilization and cutting regime; the disturbance was not visible any more. The propagation traits of the remaining species were important as those species able to spread more quickly and became dominant. If these fast colonizers had been high yielding species, we would probably have found higher yields in treated versus control plots – at least under high nutrient levels. Even a small proportion of high yielding grasses was able to increase yields in fertilized plots in the first cut. Forage quality was strongly influenced by phenological development of the sward, which hardly differed among our three sward types since they all belonged to the same plant community and had the same set of early and late flowering species, albeit in different proportions. Only at single harvest dates, the influences of forbs and grasses on quality characteristics were visible, although, most of the time, relatively small compared with fertilization effects. In the short term, the herbicide application did not lead to changes in yields contrary to the results of biodiversity experiments suggesting higher yields in swards hosting higher species diversity. However, it significantly affected forage quality. So, depending on the subsequent utilization of the forage, application of herbicides against either monocots or dicots might be useful to adapt forage quality.
Acknowledgements
This work was supported by the Ministry of Science and Culture of Lower Saxony and the ‘Niedersächsisches Vorab’ and is part of the Cluster of Excellence ‘Functional Biodiversity Research’. We would like to thank Uwe von Borstel and Bernd Gehlken, who recorded the vegetation relevés in 2009, and Peter Tillmann (VDLUFA), who was responsible for the analysis of the NIRS spectra.
References
- Andueza , D , Cruz , P , Farruggia , A , Baumont , R , Picard , F and Michalet-Doreau , B. 2010 . Nutritive value of two meadows and relationships with some vegetation traits . Grass Forage Sci. , 65 ( 3 ) : 325 – 334 .
- Balvanera , P , Pfisterer , AB , Buchmann , N , He , J-S , Nakashizuka , T , Raffaelli , D and Schmid , B. 2006 . Quantifying the evidence for biodiversity effects on ecosystem functioning and services . Ecol Lett. , 9 ( 10 ) : 1146 – 1156 .
- Bernhardt-Römermann , M , Römermann , C , Sperlich , S and Schmidt , W. 2011 . Explaining grassland biomass – the contribution of climate, species and functional diversity depends on fertilization and mowing frequency . J Appl Ecol. , 48 ( 5 ) : 1365 – 2664 .
- Bochi-Brum , O , Garcia , R , Bodas , R , Calleja , A , Andres , S and Lopez , S. 2011 . Nutritive value of herbage from mountain hay meadow managed under traditional and intensive harvest systems as affected by nitrogen fertilisation and time of cutting . Anim Prod Sci. , 51 ( 6 ) : 549 – 556 .
- Borcard , D , Legendre , P and Drapeau , P. 1992 . Partialling out the spatial component of ecological variation . Ecology. , 73 ( 3 ) : 1045 – 1055 .
- Bruinenberg , MH , Valk , H , Korevaar , H and Struik , PC. 2002 . Factors affecting digestibility of temperate forages from seminatural grasslands: a review . Grass Forage Sci. , 57 ( 3 ) : 292 – 301 .
- Bruinenberg , MH , Van Gelder , AH , Gonzalez Perez , P , Hindle , VA and Cone , JW. 2004 . Estimating rumen degradability of forages from semi-natural grasslands, using nylon bag and gas production techniques . NJAS-Wagen J Life Sci. , 51 ( 4 ) : 351 – 368 .
- Čop , J , Lavrenčič , A and Košmelj , K. 2009 . Morphological development and nutritive value of herbage in five temperate grass species during primary growth: analysis of time dynamics . Grass Forage Sci. , 64 ( 2 ) : 122 – 131 .
- Duru , M , Cruz , PP , Raouda , AHK , Ducourtieux , C and Theau , JP. 2008 . Relevance of plant functional types based on leaf dry matter content for assessing digestibility of native grass species and species-rich grassland communities in spring . Agron J. , 100 ( 6 ) : 1622 – 1630 .
- Dykmans , A , Mack , H and Weissbach , F. 1999 . “ The effect of grassland extensification on yield, forage quality and botanical composition at different grassland locations ” . In Landbauforschung Völkenrode. Mitteilungen der Bundesforschunganstalt für Landwirtschaft. [Farming research Völkenrode. Announcements of the Federal Agricultural Research Center (FAL)]. 206 125 – 139 .
- Fowler , N. 1981 . Competition and coexistence in a North Carolina grassland: II. The effects of the experimental removal of species . J Ecol. , 69 ( 3 ) : 843 – 854 .
- Frame , J and Laidlaw , AS. 2011 . Improved grassland management. , New , Ramsbury , UK : Crowood .
- Gesellschaft für Ernährungsphysiologie . 2008 . Neue Gleichungen zur Schätzung der umsetzbaren Energie für Wiederkäuer von Gras- und Maisprodukten [New equations for estimation of metabolisable energy for ruminants from grass and maize products] . Proc Soc Nutr Physiol. , 17 : 191 – 198 .
- Grime , JP. 1998 . Benefits of plant diversity to ecosystems: immediate, filter and founder effects . J Ecol. , 86 ( 6 ) : 902 – 910 .
- Grime , JP , Hodgson , JG and Hunt , R. 1988 . Comparative plant ecology: a functional approach to common British species , London , UK : Unwin Hyman .
- Hedemann , H-A. 1950 . Entwicklung und Struktur des Sollinggewölbes [Development and structure of the Solling Uplands] [dissertation] , Clausthal-Zellerfeld , Germany : Bergakademie Clausthal .
- Isselstein , J. 2005 . “ Enhancing grassland biodiversity and its consequences for grassland management and utilisation ” . In McGilloway, DA, editor. Grassland: a global resource , 305 – 320 . Wageningen , The Netherlands : Wageningen Academic Press. p .
- Jiang , L , Wan , SQ and Li , LH. 2009 . Species diversity and productivity: why do results of diversity-manipulation experiments differ from natural patterns? . J Ecol. , 97 ( 4 ) : 603 – 608 .
- Kesting , S , Wrage , N and Isselstein , J. 2009 . Herbage mass and nutritive value of herbage of extensively managed temperate grasslands along a gradient of shrub encroachment . Grass Forage Sci. , 64 ( 3 ) : 246 – 254 .
- Keuter A Hoeft I Veldkamp E Corre MD. Nitrogen response efficiency of a managed and phytodiverse temperate grassland Plant Soil. 2012 [Internet]. [cited 2012 Dec 11]; 14 http://dx.doi.org/10.1007/s11104-012-1344-y (http://dx.doi.org/10.1007/s11104-012-1344-y)
- Klapp , E. 1954 . Wiesen und Weiden. Behandlung, Verbesserung und Nutzung von Grünlandflächen , Berlin , Germany : Paul Parey .
- Klapp , E , Opitz , von and Boberfeld , W . 2006 . Taschenbuch der Gräser. , 13th , Stuttgart , Germany : Ulmer .
- Klapp , E and Stählin , A. 1936 . Standorte, Pflanzengesellschaften und Leistung des Grünlandes , Stuttgart , Germany : Ulmer .
- Klimek , S , Richter , gen , Kemmermann , A , Hofmann , M and Isselstein , J. 2007 . Plant species richness and composition in managed grasslands: the relative importance of field management and environmental factors . Biol Conserv. , 134 ( 4 ) : 559 – 570 .
- Klotz , S , Kühn , I and Durka , W. 2002 . BIOLFLOR – Eine Datenbank zu biologisch-ökologischen Merkmalen der Gefäßpflanzen in Deutschland . Schriftenreihe für Vegetationskunde. , 38 : 41 – 281 .
- Lavorel , S. 1999 . Ecological diversity and resilience of Mediterranean vegetation to disturbance . Divers Distrib. , 5 ( 1–2 ) : 3 – 13 .
- Lewis , GC and Hopkins , A. 2000 . “ Weeds, pests and diseases of grassland ” . In Hopkins A, editor. Grass its production & utilization. , 3rd , London , UK : Blackwell Science Ltd .
- Lopez , S , Carro , MD , Gonzalez , JS and Ovejero , FJ. 1991 . Rumen degradation of the main forage species harvested from permanent mountain meadows in North-western Spain . J Agr Sci. , 117 ( 03 ) : 363 – 369 .
- McLaren , JR and Turkington , R. 2010 . Ecosystem properties determined by plant functional group identity . J Ecol. , 98 ( 2 ) : 459 – 469 .
- Miller , RG. 1981 . Simultaneous statistical inference , New York , NY : Springer .
- Mokany , K , Ash , J and Roxburgh , S. 2008 . Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland . J Ecol. , 96 ( 5 ) : 884 – 893 .
- Opitz , von and Boberfeld , W . 1994 . Grünlandlehre [Grassland science] , Stuttgart , Germany : Ulmer .
- Petersen , U , Wrage , N , Köhler , L , Leuschner , C and Isselstein , J. 2012 . Manipulating the species composition of permanent grasslands – a new approach to biodiversity experiments . Basic Appl Ecol. , 13 ( 1 ) : 1 – 9 .
- Pinheiro , J , Bates , D , DebRoy , S and Sarkar , D . 2009 . The R Core Team . NLME: linear and nonlinear mixed effects models. R package version , 3 : 1 – 96 .
- Pywell , RF , Bullock , JM , Tallowin , JB , Walker , KJ , Warman , EA and Masters , G. 2007 . Enhancing diversity of species-poor grasslands: an experimental assessment of multiple constraints . J Appl Ecol. , 44 ( 1 ) : 81 – 94 .
- Quijas , S , Schmid , B and Balvanera , P. 2010 . Plant diversity enhances provision of ecosystem services: a new synthesis . Basic Appl Ecol. , 11 ( 7 ) : 582 – 593 .
- R Development Core Team . 2011 . R: a language and environment for statistical computing , Vienna , Austria : R Foundation for Statistical Computing .
- Renne , I and Tracy , B. 2007 . Disturbance persistence in managed grasslands: shifts in aboveground community structure and the weed seed bank . Plant Ecol. , 190 ( 1 ) : 71 – 80 .
- Roscher , C , Schumacher , J , Baade , J , Wilcke , W , Gleixner , G , Weisser , WW , Schmid , B and Schulze , ED. 2004 . The role of biodiversity for element cycling and trophic interactions: an experimental approach in a grassland community . Basic Appl Ecol. , 5 ( 2 ) : 107 – 121 .
- Roxburgh , SH and Wilson , JB. 2000 . Stability and coexistence in a lawn community: experimental assessment of the stability of the actual community . Oikos. , 88 ( 2 ) : 409 – 423 .
- Roy , J. 2001 . “ How does biodiversity control primary productivity? ” . In Terrestrial global productivity , Edited by: Saugier , B and Mooney , HA . San Diego , CA : Academic Press .
- Schrader , A and Kalthofen , H. 1974 . “ Gräser ” . In Biologie—Bestimmung—Wirtschaftliche Bedeutung , Berlin , Germany : VEB Deutscher Landwirtschaftsverlag .
- Schwartz , MW , Brigham , CA , Hoeksema , JD , Lyons , KG , Mills , MH and van Mantgem , PJ. 2000 . Linking biodiversity to ecosystem function: implications for conservation ecology . Oecologia. , 122 ( 3 ) : 297 – 305 .
- Seip , K , Breves , G , Isselstein , J and Abel , H. 2011 . Nitrogen excretion of adult sheep fed silages made of a mixed sward or of pure unfertilised grass alone and in combination with barley . Arch Anim Nutr. , 65 ( 4 ) : 278 – 289 .
- Seng , M , Bonorden , S , Nissen , J , Isselstein , J and Abel , H. 2008 . Fermentation patterns and nutrient contents of forb-containing silages and their effects on microbial fermentation in the artificial rumen system RUSITEC . J Agri Sci. , 146 ( 3 ) : 333 – 341 .
- Silletti , AM , Knapp , AK and Blair , JM. 2004 . Competition and coexistence in grassland codominants: responses to neighbour removal and resource availability . Can J Bot. , 82 ( 4 ) : 450 – 460 .
- Slocum , MG and Mendelssohn , IA. 2008 . Use of experimental disturbances to assess resilience along a known stress gradient . Ecol Indic. , 8 ( 3 ) : 181 – 190 .
- Šmilauer P Šmilauerová M. Asymmetric relationship between grasses and forbs: results from a field experiment under nutrient limitation Grass Forage Sci. 2012[Internet]. [cited 2012 Jul 3]; 13 http://dx.doi.org/10.1111/j.1365-2494.2012.00888.x (http://dx.doi.org/10.1111/j.1365-2494.2012.00888.x)
- Symstad , AJ and Tilman , D. 2001 . Diversity loss, recruitment limitation, and ecosystem functioning: lessons learned from a removal experiment . Oikos. , 92 ( 3 ) : 424 – 435 .
- ter Braak , CJF and Šmilauer , P. 1997–2004 . Canoco for windows 4.53. Wageningen , The Netherlands : Plant Research International .
- ter Braak , CJF and Šmilauer , P. 2002 . Canoco reference manual and CanoDraw for Windows User's guide: software for canonical community ordination (version 4.5) , Ithaca , NY : Microcomputer Power .
- Tillmann , P. 2010 . Anwendung der Nahinfrarotspektroskopie (NIRS) an Grünlandproben [Usage of near-infrared reflectance spectroscopy for grassland samples] . VDLUFA-Schriftenreihe. , 66 : 145 – 150 .
- Tilman , D. 1997 . Distinguishing between the effects of species diversity and species composition . Oikos. , 80 ( 1 ) : 185
- Touzard , B and Clement , B. 2001 . Plant diversity dynamics in an eutrophic alluvial reed bed after experimental small-scale disturbances . Bot Helv. , 111 ( 1 ) : 45 – 58 .
- van Soest , PJ , Mertens , DR and Deinum , B. 1978 . Preharvest factors influencing quality of preserved forage . J Anim Sci , 47 ( 3 ) : 712 – 720 .
- Vandvik , V and Birks , H. 2002 . Partitioning floristic variance in Norwegian upland grasslands into within-site and between-site components: are the patterns determined by environment or by land-use? . Plant Ecol. , 162 ( 2 ) : 233 – 245 .
- Venables , WN and Ripley , BD. 2002 . Modern applied statistics with S. , 4th , New York , NY : Springer .
- Virágh , K. 1987 . The effect of herbicides on vegetation dynamics: a five year study of temporal variation of species composition in permanent grassland plots . Folia Geobot. , 22 ( 4 ) : 385 – 403 .
- Virágh , K. 1989 . An experimental approach to the study of community stability – resilience and resistance . Acta Botanica Hungarica. , 35 ( 1–4 ) : 99 – 125 .
- Voigtländer , G and Jacob , H. 1987 . Grünlandwirtschaft und Futterbau , Stuttgart , Germany : Ulmer .
- Waide , RB , Willig , MR , Steiner , CF , Mittelbach , G , Gough , L , Dodson , SI , Juday , GP and Parmenter , R. 1999 . The relationship between productivity and species richness . Annu Rev Ecol Syst. , 30 : 257 – 300 .
- White , TA , Barker , DJ and Moore , KJ. 2004 . Vegetation diversity, growth, quality and decomposition in managed grasslands . Agric Ecosyst Environ. , 101 ( 1 ) : 73 – 84 .
- Wilman , D and Riley , JA. 1993 . Potential nutritive value of a wide range of grassland species . J Agri Sci. , 120 ( 1 ) : 43 – 50 .
- Wrage , N , Strodthoff , J , Cuchillo , MH , Isselstein , J and Kayser , M. 2011 . Phytodiversity of temperate permanent grasslands: ecosystem services for agriculture and livestock management for diversity conservation . Biodivers Conserv. , 20 ( 14 ) : 3317 – 3339 .
- Yandell , BS. 1997 . Practical data analysis for designed experiments , Boca Raton , FL : Chapman & Hall/CRC .
- Zuur , AF , Ieno , EN , Walker , N , Saveliev , AA and Smith , GM. 2009 . Mixed effects models and extensions in ecology in R , New York , NY : Springer .