
ABSTRACT
High-value hardwood species such as monarch birch (Betula maximowicziana) and castor aralia (Kalopanax septemlobus) are important economic and ecological elements of cool-temperate mixed forests in northern Japan. This article presents the single-tree management system for high-value timber species as practised for 50 years at the University of Tokyo Hokkaido Forest. Nearly 2000 valuable broad-leaved trees meeting the size and quality criteria have been registered as ‘superior trees’, and their status is periodically monitored for timing of harvest. A case study was conducted using 2105 inventory plots to characterize the stand types in which superior trees occur. A total of 57 superior trees of 11 broad-leaved species was found in 2.2% of the inventory plots. The results indicated that superior trees generally grew in mature species-rich stands. Superior trees of some species may have promoted their abundance by dispersing relatively more seeds to the surroundings. Single-tree management facilitates the sustainable use of high-value timber species by explicitly monitoring the numbers, attributes and locations of superior trees, and contributes to conserving stand structural diversity through protection of these large-sized canopy trees, which promotes ecological values such as biomass and carbon storage, species diversity, seed abundance and bird habitat. The production of fancy wood from superior trees earns significant income through extremely high log prices (maximum > 20,000 USD m–3).
EDITED BY Nicholas Brokaw
Introduction
High-value timber species are those used for products with extremely high commercial value. The monarch birch (Betula maximowicziana Regel) and castor aralia (Kalopanax septemlobus (Thunb.) Koidz) are the most valuable timber species of cool-temperate mixed forests in northern Japan, fetching log prices of up to 20,000 USD m–3. Other high-value species include big-leaf mahogany (Swietenia macrophylla King) and ipê (Tabebuia spp.) in Latin America (Kometter et al. Citation2004; Grogan et al. Citation2010), African mahogany (Entandrophragma spp.) in Africa (Romeiras et al. Citation2014) and rosewood (Dalbergia cochinchinensis Pierre) in Southeast Asia (So et al. Citation2010). In Europe, valuable broad-leaved genera (Acer, Alnus, Betula, Fraxinus, Juglans, Prunus, Sorbus, Tilia and Ulmus spp.) produce high-value timber that is used in the veneer and furniture industries (Hemery et al. Citation2008; Oosterbaan et al. Citation2009).
Even though high-value timber species typically occur at very low densities (Schulze et al. Citation2008), they play important economic and ecological roles in forest management (Oosterbaan et al. Citation2009). Trees with high economic value are often large and old. Large old trees are key structural attributes in forest ecosystems (Lindenmayer et al. Citation2012), with regard to biomass, carbon storage (Lutz et al. Citation2012; Sist et al. Citation2014; Kauppi et al. Citation2015) and structural heterogeneity (Lutz et al. Citation2013). Their social and cultural values are also significant (Blicharska & Mikusiński Citation2014). Management of valuable large old trees may help conserve both the economic value and the biodiversity of the forests they inhabit.
Due to their economic value, high-value timber species are often intensively harvested with little attention to the impacts on their population structures and dynamics (Schulze et al. Citation2008). They suffer from illegal logging (Ward et al. Citation2008) and are becoming increasingly scarce in heavily logged forests (Prates-Clark et al. Citation2008). The most valuable timber species are often the most threatened ones (So et al. Citation2010). Thus, fundamental changes in forest management practices are urgently needed to conserve existing trees of high-value timber species, as well as forest biomass and structural heterogeneity. Lindenmayer et al. (Citation2014) suggested creating a tree registry system that aims at promoting their protection. He suggested that single-tree management of valuable tree individuals should replace unplanned harvests by explicitly identifying tree attributes and locations in forests.
In this article, we describe single-tree management of high-value timber species in a cool-temperate mixed forest in northern Japan. A case study was conducted at the University of Tokyo (UTokyo) Hokkaido Forest, where single-tree management for high-value hardwoods has been practised since the mid-1960s, and nearly 2000 trees with superior timber quality have been registered as ‘superior trees’ and monitored on an individual-tree basis. Our specific objectives were (1) to identify how stands in which superior trees occur differ from stands without such trees, in terms of tree density, basal area (BA) and species richness and (2) based on differences between stands with and without superior trees, to consider how single-tree management for superior trees might help to conserve forest biodiversity. We also present the economic benefits of single-tree management for high-value trees.
Methods
UTokyo Hokkaido Forest and management policies
The UTokyo Hokkaido Forest was established in 1899, when national forests were transferred from the Ministry of Home Affairs to the UTokyo. It is located in the centre of Hokkaido Island, in northernmost Japan (43°10′–20′ N, 142°18′–40′ E, 190–1459 m asl), and has an area of 22,716 ha. The mean annual temperature and precipitation at the arboretum of the Forest (230 m asl) during 2001‒2008 were 6.3°C and 1210 mm, respectively (The University of Tokyo Hokkaido Forest Citation2012). Snow (maximum 83 cm in depth) usually covers the ground from late November to early April. The typical substrate and soil type are welded tuff and dark-brown forest soil. The Forest is situated in the pan-mixed forest zone (Tatewaki Citation1958), which is transitional between deciduous forests in the cool-temperate zone and coniferous forests in the Sub-Boreal zone. Uneven-aged mixed forests with coniferous and broad-leaved tree species are the main vegetation cover. As of 2011, the total and mean ha–1 growing stock of the Forest were 4.8 million m3 and 211 m3 ha–1, respectively (The University of Tokyo Hokkaido Forest Citation2012).
A large-scale and long-term experiment on the stand-based silvicultural management system (Takahashi Citation1961; Watanabe & Sasaki Citation1994) has been conducted at the UTokyo Hokkaido Forest since 1958. The idea behind the system is that management should be adapted to the conditions of each stand to maximize the multiple public and economic functions of forest ecosystems (Takahashi Citation2001). Single-tree selection harvest has been implemented as the main silvicultural system at the Forest, in which trees are periodically selected and harvested individually from a large area (Zingg Citation1999). The cutting cycle is either 15 or 20 years, depending upon stand growth rates and accessibility. In each managed area, the rate of tree removal is 10–17% of the growing stock per cutting cycle, which is lower than the stock growth rate (The University of Tokyo Hokkaido Forest Citation2012). Logging operations typically use felling with chainsaws and skidding of the felled stems by a winch-equipped crawler tractor (Tatsumi et al. Citation2014). The mean annual tree removal and timber sales (stumpage and logs) during 2006–2010 were 23,100 m3 and 120 million Japanese Yen (JPY) (approximately 1 million USD), respectively (The University of Tokyo Hokkaido Forest Citation2012).
Single-tree management for ‘superior trees’
To maintain and sustainably produce high-value timber in managed stands, the UTokyo Hokkaido Forest has been practicing the single-tree management system since 1965 (Shibata Citation1988; Yamamoto et al. Citation1989; Yamamoto Citation1990). The system has three components: (1) single-tree selection and registration, (2) measurement and assessment and (3) periodic monitoring for optimal harvest.
Under the single-tree management system, large-sized canopy trees with high timber quality and value have been individually selected and registered as ‘superior trees’ (The Tokyo University Forest in Hokkaido Citation2012). Operational guidelines for selecting superior trees include diameter at breast height (DBH ≥ 40 cm), crown height (≥4 m) and estimated length of butt log (≥4 m) (The University of Tokyo Hokkaido Forest Citation2012). There are also qualitative criteria for single-tree selection: the trunk should be straight, the cross section should be a circle and the stem should have few defects such as bending, twisting, knots from branches or dormant buds, and decay.
All superior trees are numbered, and DBH, species name, tree vitality (based on visual assessment of tree crown) and stem damage (if any) are recorded. The geographic locations have been partly obtained using a global navigation satellite system (GNSS) receiver with positional accuracy < 5 m (Owari et al. Citation2011; Tokuni et al. Citation2016). As shown in , registered superior trees are sparsely distributed over the management area. This precise information on individual tree positions is, therefore, crucial for single-tree management practices. The status of registered superior trees is periodically monitored by tree-marking crews for harvest at the optimal time (see below).
Figure 1. An example of location map for registered superior trees (Compartment no. 56) at the University of Tokyo Hokkaido Forest.
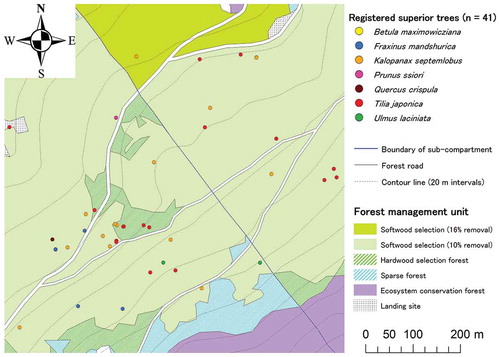
A total of 1952 superior trees from 19 valuable broad-leaved species was listed on the registry as of September 2014. Major species in the registry and included in this study were K. septemlobus (Thunb.) Koidz, Quercus crispula Blume, Tilia japonica (Miq.) Simonk., B. maximowicziana Regel and Fraxinus mandshurica Rupr. (). Their mean DBH was 64.3 cm, with a range of 40–110 cm. Shibano et al. (Citation1995) found that superior B. maximowicziana trees that had been harvested during 1988–1993 ranged between 105 and 316 years in age and 48–108 cm in DBH.
Figure 2. Major high-value hardwood species at the University of Tokyo Hokkaido Forest: (a) monarch birch (B. maximowicziana), (b) castor aralia (K. septemlobus), (c) Japanese oak (Q. crispula) and (d) Japanese ash (F. mandshurica).

According to the 10-year forest management plan for 2011–2020, 100 m3 of superior trees are to be harvested annually (The University of Tokyo Hokkaido Forest Citation2012). Superior trees are harvested on the basis of tree vitality assessment. Trees with ≥50% of branch death in the crown are usually considered candidates for harvest (Kuboyama Citation1994). In practice, more than half of the superior trees have been logged within a few years after death. A dead B. maximowicziana tree can be sold at an extremely high price if it is large enough, because after death, the heartwood often turns an elegant pink that buyers prefer (Kuboyama Citation1993). On the other hand, K. septemlobus should be harvested while alive, because the species is prone to rapidly losing timber quality and market value after the death (Kuboyama Citation1994).
Data collection and analysis
A total of 2906 inventory plots was established and measured during 2006–2013 within the UTokyo Hokkaido Forest (Owari Citation2013). Excluding plots located in plantation forests, we used the data from 2105 inventory plots for our preliminary analysis. These plots had an aggregated area of 452.4 ha, and the plot size was normally 0.25 ha (50 m × 50 m). The species and DBH classes (2 cm intervals) for all live trees with DBH ≥ 5 cm were recorded in each plot. We determined whether registered superior trees occurred in each plot from the measurement record. For this study, we focused on T. japonica, K. septemlobus, F. mandshurica, B. maximowicziana and Q. crispula.
To characterize the stands where superior trees of these five species occur and that are under single-tree management, tree density (number of tree individuals per hectare), BA and the number of tree species per inventory plot with superior trees were determined. A nonparametric Brunner–Munzel test (Brunner & Munzel Citation2000) was then performed to examine the differences in these parameters between inventory plots with and without superior trees. Stand parameters (including all species) were computed for plots with superior trees of each of the five focal species.
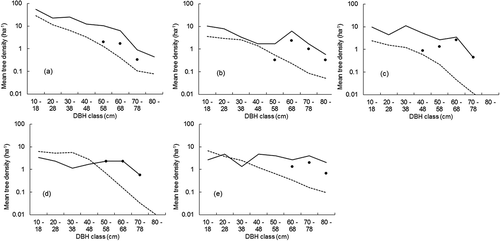
We examined the abundance (number of tree individuals observed) for each of the five focal species in the inventory plots by graphing rank-species abundance diagrams, with log-abundance for mean tree density or BA plotted on the y-axis, and species ranked consecutively by abundance on the x-axis (McGill et al. Citation2007). Species mean abundances based on tree density () and BA () were computed for the inventory plots where superior trees of each of the focal species were found and also for all inventory plots. Relative abundance (number of trees of a species as a percentage of the total number of trees) was calculated for each of the five focal species. A Wilcoxon signed-rank test was used to compare the relative abundance in tree density and BA for each focal species, from plots with superior trees for that species and from all plots. Average diameter class densities () were also determined for inventory plots with superior trees and also for the inventory plots without superior trees. A Brunner–Munzel test was performed to compare the diameter class densities of large-sized trees (≥50 cm DBH) between inventory plots with and without superior trees.
Figure 3. Rank-species abundance diagram using mean tree density, for inventory plots in which superior trees of particular high-value species were observed, compared to all inventory plots, at the University of Tokyo Hokkaido Forest: (a) T. japonica, (b) K. septemlobus, (c) F. mandshurica, (d) B. maximowicziana and (e) Q. crispula. Log-abundance represented by the mean tree density (number of tree individuals per hectare) is plotted on the y-axis, while species are ranked consecutively on the x-axis. The solid line indicates the abundance of each species for the inventory plots in which the superior trees were observed (see ), while the dashed line represents all inventory plots (n = 2105). The ranks of the particular high-value tree species are marked by a solid circle (the inventory plots in which superior trees were observed) and an open circle (all inventory plots).
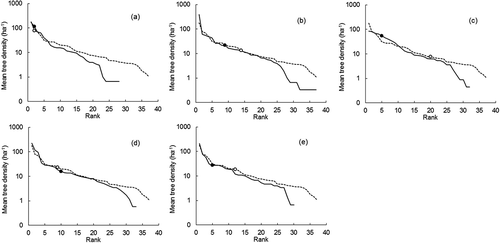
Figure 4. Rank-species abundance diagrams using mean basal area, for inventory plots in which superior trees of particular high-value species were observed, compared to all inventory plots, at the University of Tokyo Hokkaido Forest: (a) T. japonica, (b) K. septemlobus, (c) F. mandshurica, (d) B. maximowicziana and (e) Q. crispula. Log-abundance represented by the mean basal area is plotted on the y-axis, while species are ranked consecutively on the x-axis. The solid line indicates the abundance of each species for the inventory plots in which the superior trees were observed (see ), while the dashed line represents all inventory plots (n = 2105). The ranks of the particular high-value tree species are marked by a solid circle (the inventory plots in which superior trees were observed) and an open circle (all inventory plots).
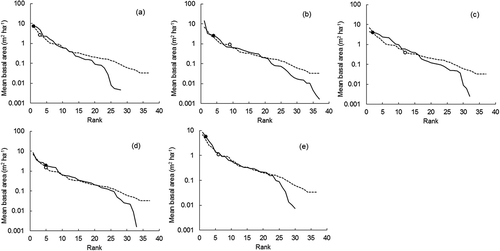
Figure 5. Average diameter class densities for particular high-value species in the inventory plots at the University of Tokyo Hokkaido Forest: (a) T. japonica, (b) K. septemlobus, (c) F. mandshurica, (d) B. maximowicziana and (e) Q. crispula. The solid line indicates the diameter class densities for the inventory plots in which superior trees for the particular species were observed (see ), and the densities of the superior trees in each DBH class are marked by solid circles. The dashed line represents the diameter class densities for the particular species in the inventory plots in which the superior trees for the particular species were not found.
The economics of high-value timber species was also examined. The UTokyo Hokkaido Forest has been selling fancy hardwood logs produced from superior trees at the auction market in Asahikawa, central Hokkaido (Okamura & Goto Citation2004), once or twice a year since 1987 (Kuboyama Citation1994). The unit prices and total sales of fancy logs for each species in January 2014 and January 2015 were calculated using the sales record at the auction market.
Results and discussion
Stand type of ‘superior trees’
A total of 327,198 naturally grown trees ≥ 5 cm DBH of 53 species was found within the 2105 inventory plots. The most common species was the conifer Abies sachalinensis (F. Schmidt) Mast. (80,758 trees; 25% of the total), followed by T. japonica (34,495 trees; 11%), Acer pictum Thunb. (31,279 trees; 10%) and Picea jezoensis (Siebold & Zucc.) Carrière (22,960 trees; 7%). High-value timber species of B. maximowicziana (9947 trees; 3%), Q. crispula (7980 trees; 2%), K. septemlobus (5995 trees; 2%) and F. mandshurica (3467 trees; 1%) were relatively rare in the inventory plots.
We found 57 superior trees of 11 broad-leaved species in 2.2% of the inventory plots (46 out of 2105 plots; ). The mean density of superior trees over the inventory plots was calculated at 0.13 trees ha–1. The mean DBH of superior trees was 62.3 cm, and ranged between 40 and 94 cm. Among superior trees of all species, the species in this study (T. japonica, K. septemlobus, F. mandshurica, B. maximowicziana and Q. crispula) had the most individual trees. Other species with superior trees included A. pictum Thunb., Betula ermanii Cham., Cercidiphyllum japonicum Sieb. & Zucc, Phellodendron amurense Rupr., T. maximowicziana Shiras. and Ulmus laciniata (Trautv.) Mayr.
Table 1. Stand parameters of the inventory plots in which superior trees were found (mean ± s.d.) at the University of Tokyo Hokkaido Forest.
Within the inventory plots in which superior trees were found, the mean density of all tree species was 725 trees ha–1. The difference in total tree density between plots with and without superior trees was not significant (p = 0.38). The mean BA (31.2 m2 ha–1) and the mean number of species observed (17.0 species) were both significantly larger in inventory plots with superior trees than in inventory plots without superior trees (p < 0.001). These results indicated that superior trees of all species generally grew in mature and species-rich stands. As the understorey develops, the number of plant species increases, and the habitat for animals is enhanced in the mature stage of stand development (Fujimori Citation2001). Since BA has a linear relation to the above-ground biomass (Chiba Citation1998), there were greater biomass and carbon stocks in stands where superior trees grew. By carefully managing superior trees and the surroundings, tree species diversity and high carbon storage may have been maintained.
Although stand parameters varied widely for each species, superior K. septemlobus trees tended to occur in plots with relatively high tree density (862 trees ha–1) and large BA (34.1 m2 ha–1) (). The number of tree species was relatively large in plots with superior B. maximowicziana trees (19.1 species; ). Superior trees of F. mandshurica and T. japonica were found in plots having relatively lower tree density (660 and 648 trees ha–1) and fewer tree species (14.2 and 16.1 species) (), probably because these two species are often found in mixed stands dominated by broad-leaved species, which typically have lower density than stands dominated by coniferous species (The Tokyo University Forest in Hokkaido Citation2007).
Relative species abundance and size distributions
shows rank-species abundance for inventory plots in which superior trees of the focal species were observed, compared to the rank-species abundance for all inventory plots. There were more trees of K. septemlobus in inventory plots with superior trees of this species than in plots without superior trees of K. septemlobus. The same was true for F. mandshurica and Q. crispula. The seed size is relatively large in F. mandshurica (>100 mg) and Q. crispula (>2000 mg) (Seiwa Citation1994). Although K. septemlobus produces small seeds (>1 mg), the dispersal depends on birds. For these species, superior trees may have promoted their own abundance within inventory plots by dispersing more seeds to the immediate surroundings. The presence of a same-species superior tree did not affect the ranks of species abundance in T. japonica and B. maximowicziana, which have wind-dispersed seeds, and whose seeds were probably dispersed mostly outside of the inventory plots. We obtained similar results when the species abundance distribution in terms of mean BA was examined (). For the five high-value tree species focused on in this study, the mean relative abundance was highest in T. japonica (18.3% in tree density and 23.2% in BA) and lowest in B. maximowicziana (2.1% in tree density and 6.3% in BA). In the five species investigated, the relative abundance was significantly higher in BA than in tree density (p < 0.05), indicating the relative dominance of large-sized trees within these species.
shows the mean tree diameter class densities in inventory plots in which superior trees were observed for each of the five focal species, compared to the mean diameter class densities of the same species in the inventory plots in which the superior trees for the particular species were not observed. In the inventory plots having superior trees, large-sized trees were mostly registered as superior trees. For all five species, the diameter class densities of large-sized trees (≥50 cm DBH) were significantly greater in the inventory plots with superior trees than the inventory plots without superior trees (p < 0.001). Under the single-tree management system, these superior trees have been excluded from periodic selection harvests. This result indicates that the single-tree management contributes to forest structural diversity through the protection of large-sized high-value timber trees, which are often preferred by cavity nesting birds (Suzuki et al. Citation1983) and perching raptors (Berkelman et al. Citation2002). A relatively few trees in the largest size classes dominate stand-level above-ground biomass (Enquist & Niklas Citation2001; Lutz et al. Citation2012).
Economics of high-value timber
The high-value hardwoods B. maximowicziana, K. septemlobus, F. mandshurica and C. japonicum were sold at the auction market in January 2014 and January 2015 (). The total sales from fancy hardwood logs (41.6 million JPY) accounted for 25% of the total income (168.3 million JPY) during the period for the UTokyo Hokkaido Forest. Among the species sold, B. maximowicziana was the most expensive, and for this species the highest log price in January 2015 was 2,370,600 JPY (20,090 USD) m–3. K. septemlobus had the second highest log price, which was 880,200 JPY (7459 USD) m–3 in the same year. These two species are mainly used to produce face veneer for decorative plywood. For F. mandshurica, which is mainly used for furniture, the average price in 2014 was 73,351 JPY (622 USD) m–3. Market preference, resource scarcity, as well as the type of final products may have affected the market price of high-value timber species. Since the implementation of single-tree management requires costs and efforts for searching for, marking and periodically monitoring individual trees, it is feasible only when log price is very high.
Table 2. Sales of fancy hardwood logs by auction from the University of Tokyo Hokkaido Forest between January 2014 and January 2015, in Asahikawa, central Hokkaido.
Conclusions
At the UTokyo Hokkaido Forest, the single-tree management system facilitates the sustainable use of high-value timber species by explicitly listing the numbers, attributes and locations of superior trees in the forests. The system also maintains large-sized canopy trees in the forest, which, in turn, maintains the high carbon storage and structural heterogeneity that supports species diversity by providing abundant seed sources and diverse wildlife habitat. The production of fancy wood from high-value timber species brings significant income through extremely high log prices.
Although the risks of illegal logging are currently negligible in the country, the precise locations of superior trees may need to be kept secret (Lindenmayer et al. Citation2014). Due to budget and labour constraints, however, the periodic field assessment of superior trees across the management area may become difficult in the near future. Single-tree management is most likely to be applicable to large-sized tree individuals of valuable species having extremely high timber prices. To expand its applicability to other trees and species, innovative single-tree management techniques such as species distribution modelling with remote sensing data (Prates-Clark et al. Citation2008) may be useful as they can provide more cost-effective alternatives.
Acknowledgements
We would like to thank current and former technical staff members at the UTokyo Hokkaido Forest for their substantial ongoing efforts for management practices. We are grateful to two anonymous reviewers and editors for their helpful and useful comments on an earlier version of this article.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Berkelman J, Fraser JD, Watson RT. 2002. Nesting and perching habitat use of the Madagascar Fish-Eagle. J Raptor Res. 36:287–293.
- Blicharska M, Mikusiński G. 2014. Incorporating social and cultural significance of large old trees in conservation policy. Conserv Biol. 28:1558–1567.
- Brunner E, Munzel U. 2000. The nonparametric Behrens-Fisher problem: asymptotic theory and a small-sample approximation. Biom J. 42:17–25.
- Chiba Y. 1998. Architectural analysis of relationship between biomass and basal area based on pipe model theory. Ecol Model. 108:219–225.
- Enquist BJ, Niklas KJ. 2001. Invariant scaling relations across tree-dominated communities. Nature. 410:655–660.
- Fujimori T. 2001. Ecological and silvicultural strategies for sustainable forest management. Amsterdam: Elsevier.
- Grogan J, Blundell AG, Landis RM, Youatt A, Gullison RE, Martinez M, Kómetter R, Lentini M, Rice RE. 2010. Over-harvesting driven by consumer demand leads to population decline: big-leaf mahogany in South America. Conserv Lett. 3:12–20.
- Hemery G, Spiecker H, Aldinger E, Kerr G, Collet C, Bell S. 2008. COST Action E42: growing valuable broadleaved tree species. Final Report. 40 pp. Available at http://www.valbro.uni-freiburg.de/
- Kauppi PE, Birdsey RA, Pan Y, Ihalainen A, Nöjd P, Lehtonen A. 2015. Effects of land management on large trees and carbon stocks. Biogeosciences. 12:855–862.
- Kometter RF, Martinez M, Blundell AG, Gullison RE, Steininger MK, Rice RE. 2004. Impacts of unsustainable mahogany logging in Bolivia and Peru. Ecol Soc. 9:12. [online].
- Kuboyama H. 1993. Koyoju yuryo zai no seisan hanbai ni kansuru keizai bunseki: Tokyo Daigaku Hokkaido Enshurin wo jirei to shite [An economic analysis of producing and selling high-quality hardwoods: a case study in the Tokyo University Forest in Hokkaido]. Ringyo Keizai Kenkyu [Journal of Forest Economics (Japan)]. 123:105–109. (in Japanese).
- Kuboyama H. 1994. On the price formation mechanism and the selling method of high-quality hardwoods: a case study in the Tokyo University Forest in Hokkaido. Buu Tokyo Univ For. 91:1–21. (in Japanese with English summary).
- Lindenmayer DB, Laurance WF, Franklin JF. 2012. Global decline in large old trees. Science. 338:1305–1306.
- Lindenmayer DB, Laurance WF, Franklin JF, Likens GE, Banks SC, Blanchard W, Gibbons P, Ikin K, Blair D, McBurney L, et al. 2014. New policies for old trees: averting a global crisis in a keystone ecological structure. Conserv Lett. 7:61–69.
- Lutz JA, Larson AJ, Freund JA, Swanson ME, Bible KJ. 2013. The importance of large-diameter trees to forest structural heterogeneity. PLoS ONE. 8:e82784.
- Lutz JA, Larson AJ, Swanson ME, Freund JA. 2012. Ecological importance of large-diameter trees in a temperate mixed-conifer forest. PLoS ONE. 7:e36131.
- McGill BJ, Etienne RS, Gray JS, Alonso D, Benecha HK, Dornelas M, Enquist BJ, Green JL, He F, Hurlbert AH, et al. 2007. Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework. Ecol Lett. 10:995–1015.
- Okamura K, Goto S. 2004. Evaluation and market price of 6 hardwood species logs in Asahikawa premium hardwood market. Trans Meet Hokkaido Branch Jpn For Soc. 52:171–173. (in Japanese).
- Oosterbaan A, Hochbichler E, Nicolescu V-N, Spiecker H. 2009. Silvicultural principles, goals and measures in growing valuable broadleaved tree species. Die Bodenkultur. 60:45–51.
- Owari T. 2013. Sustainable and adaptive forest management and data infrastructure under stand-based silvicultural system. Proc Inst Statist Math. 61:201–216. (in Japanese with English summary).
- Owari T, Kimura N, Shimizume M, Takuma R, Nakagawa Y, Tookuni M. 2011. Using global positioning system (GPS) technology for tree marking in a natural forest under a single-tree selection system. FORMATH. 10:105–121.
- Prates-Clark CDC, Saatchi SS, Agosti D. 2008. Predicting geographical distribution models of high-value timber trees in the Amazon Basin using remotely sensed data. Ecol Model. 211:309–323.
- Romeiras MM, Figueira R, Duarte MC, Beja P, Darbyshire I. 2014. Documenting biogeographical patterns of African timber species using herbarium records: a conservation perspective based on native trees from Angola. PLoS ONE. 9:e103403.
- Schulze M, Grogan J, Landis RM, Vidal E. 2008. How rare is too rare to harvest? Management challenges posed by timber species occurring at low densities in the Brazilian Amazon. For Ecol Manage. 256:1443–1457.
- Seiwa K. 1994. Effects of seed size and seedling phenology on seedling establishment of deciduous broad-leaved tree species. Bull Hokkaido For Res Inst. 31:1–68. (in Japanese with English summary).
- Shibano S, Kasahara H, Kimura N, Fukushi K, Iguchi K, Okamura K, Takahashi Y. 1995. Processes of diameter growth for large trees of Betula maximowiczinana: growth declining and dead trees at the Tokyo University Forest in Hokkaido. Trans Annu Meet Jpn For Soc. 106:263–264. (in Japanese).
- Shibata S. 1988. Studies on the management of natural forests: an experiment of natural forest management in the Tokyo University Forest in Hokkaido. Bull Tokyo Univ For. 80:269–297. (in Japanese with English summary).
- Sist P, Mazzei L, Blanc L, Rutishauser E. 2014. Large trees as key elements of carbon storage and dynamics after selective logging in the Eastern Amazon. For Ecol Manage. 318:103–109.
- So T, Theilade I, Dell B. 2010. Conservation and utilization of threatened hardwood species through reforestation: an example of Afzelia xylocarpa (Kruz.) Craib and Dalbergia cochinchinensis Pierre in Cambodia. Pac Conserv Biol. 16:101–116.
- Suzuki T, Saito S, Saito M. 1983. Bird populations during the breeding season in a deciduous broad-leaved forest at Iwamizawa, central Hokkaido. Bull Hokkaido For Res Inst. 21:95–103. (in Japanese with English summary).
- Takahashi N. 1961. Rinbun segyo ho gairon: enshurin ni okeru rekishi to jikken [Introduction to the stand-based silvicultural management system: history and experiments at the University Forest]. Hoppo Ringyo [Northern Forestry]. 13:105–109. (in Japanese).
- Takahashi N. 2001. Rinbun segyo ho: sono kangae to jissen (kaitei ban) [The stand-based silvicultural management system: its theory and practices (Revised Ed.)]. Sapporo: Log Bee. (in Japanese).
- Tatewaki M. 1958. Forest ecology of the islands of the North Pacific Ocean. J Fac Agric Hokkaido Univ. 50:371–486.
- Tatsumi S, Owari T, Kasahara H, Nakagawa Y. 2014. Individual-level analysis of damage to residual trees after single-tree selection harvesting in northern Japanese mixedwood stands. J For Res. 19:369–378.
- The Tokyo University Forest in Hokkaido. 2007. The 12th management and experiment on the Tokyo University Forest in Hokkaido (2006–2015). Misc Inform Tokyo Univ For. 46:215–350. (in Japanese)
- The University of Tokyo Hokkaido Forest. 2012. The 13th education and research plan on the University of Tokyo Hokkaido Forest (2011–2020). Misc Inform Univ Tokyo For. 51:67–176. (in Japanese)
- Tokuni M, Owari T, Hirata M, Suzuki M, Takahashi K, Kasahara H, Shibano H. 2016. Positioning accuracy of mapping- and consumer-grade global navigation satellite system (GNSS) receivers in a forest environment. Bull Univ Tokyo For. 134:29–40. (in Japanese with English summary).
- Ward SE, Boshier D, Grogan J. 2008. Special issue: sustainable management of high-value timber species of the Meliaceae: introduction. For Ecol Manag. 255:265–268.
- Watanabe S, Sasaki S. 1994. The silvicultural management system in temperate and boreal forests: a case history of the Hokkaido Tokyo University Forest. Can J For Res. 24:1176–1185.
- Yamamoto H. 1990. Studies on an integrated computer-based system of forest management in natural selection forest. Bull Tokyo Univ For. 83:31–142. (in Japanese with English summary).
- Yamamoto H, Nagumo H, Watanabe S. 1989. The selection cutting system of high valued natural hardwoods: a new method of managing natural forests. J Jpn For Soc. 17:1–9. (in Japanese with English summary).
- Zingg A. 1999. English and German terminologies in forestry research on growth and yield: a few example. For Snow Landsc Res. 74:179–187.