Abstract
Fine root (<2 mm in diameter) biomass and nutrient concentrations were examined by a sequential core method at a soil depth of the top 15 cm in a Korean pine (Pinus koraiensis) plantation and a sawtooth oak (Quercus acutissima) stand in the Forest Practice Research Center, Gyeonggi-do, Korea. Live and dead fine root biomass showed a seasonal variation in pine and oak stands. Live fine root biomass generally decreased in the late growing season (September–November) with a seasonal peak in the early growing season (April–June), while dead fine root biomass in both stands showed high values in summer (July or August). Mean fine root (live, dead, total) biomass (g m−2) during the study period was higher for the Korean pine (142, 266, 408) than for the sawtooth oak stands (124, 193, 317). Nitrogen and phosphorus concentrations of live fine roots were little changed during the growing season, while other nutrient concentrations showed seasonal fluctuations during the study period. Nutrient concentrations were generally higher in the Korean pine than in the sawtooth roots except for calcium and phosphorus concentrations. The results indicate that fine roots could be attributed to below-ground nutrient cycling by the difference of biomass and nutrient concentrations induced from each stand type.
Introduction
Fine roots (<2 mm in diameter) in forest stands represent a large and dynamic portion of the below-ground biomass and nutrient capital because significant amounts of organic matter and nutrients in the soils can be transferred during root decomposition (Jackson et al. Citation1997; Silver and Miya Citation2001; Majdi et al. Citation2005; Yuan and Chen Citation2010). Although fine roots represent only a small proportion of total plant biomass in forest ecosystems, fine roots play important roles as a source and sink for nutrients in forest ecosystems (Gill and Jackson Citation2000). The fine roots biomass depends on several ecological factors and forest management activities such as forest types, climate, site quality, stand age, stand density, fertilization, and thinning (Chen et al. Citation2001; Borken et al. Citation2007; Hwang et al. Citation2007; Finér et al. Citation2011). In addition, tree species can have a significant influence on organic matter and nutrient inputs by roots because of the difference in quantity and quality of root litters produced by different species which have different nutrient requirements (Bolte and Villanueva Citation2006; Hwang et al. Citation2007; Finér et al. Citation2011).
Although numerous studies to evaluate the effect of forest types on root biomass are reported (Park et al. Citation2006; Hwang et al. Citation2007; Finér et al. Citation2011), major uncertainties remain about the importance and behavior of roots biomass in forest ecosystems in Korea. In addition, few root studies were conducted to determine fine root biomass and nutrient dynamics in Korea (Son and Hwang Citation2003; Park et al. Citation2006; Hwang et al. Citation2007). Korean pine (Pinus koraiensis) has become one of the most major planting species for forest regeneration throughout the country in the last 30 years. Also, a sawtooth oak (Quercus acutissima) is one of the dominant hardwood species in natural stands in Korea. Better understanding of biomass and nutrient dynamics in fine roots of both tree species is necessary because reliable information on root biomass is limited. The objectives of this study were: (1) to examine seasonal patterns of fine root biomass; (2) to determine nutrient (N, P, K, Ca, Mg) concentrations from fine roots in a Korean pine and a sawtooth oak stands on a similar environmental condition.
Materials and methods
The study was conducted in the Forest Practice Research Center (National Forest Experiment Station in Gwangnung), Gyeonggi-do, Korea. Annual precipitation in this site averages 1365 mm and is higher than the average of the country (1274 mm). The study site consisted of an approximately 70-year-old natural sawtooth oak stand and a 31-year-old Korean pine plantation. Dominant understory species were Capinus laxiflora, Viburnum dilatatum, Stephanandra incisa, Disporum smilacinum, and Syneilesis palmata in the sawtooth oak stand. C. laxiflora, V. dilatatum, and D. smilacina were dominant in the Korean pine plantation. Treatment plots were divided into three 20 × 10 m plots in each stand. The mean tree density was 1850 trees ha−1 in the Korean pine plantation and 550 trees ha−1 in the sawtooth oak stand. The mean basal area was approximately 28 m2 ha−1 in the Korean pine plantation and 33 m2 ha−1 in the sawtooth oak stand, respectively (). The soil in the study site is a well-drained, slightly wet, brown forest soil (mostly Inceptisols) originating from granite, with a loamy texture ().
Table 1. General characteristics of the study site.
Table 2. Soil properties in the top 15 cm of mineral soil.
Fine root samples were estimated monthly during the growing season by sequential soil coring method except for the winter season at three random locations in each treatment plot for 2 years (April 1995–October 1996). Sequential soil coring technique was the most widespread approach to quantify fine root biomass in forest stands (Hendricks et al. Citation2006). Three cores in each plot (total 9 cores in each stand) were collected at 15 cm soil depth by a stainless steel corer (5 cm diameter and 15 cm long). Soil core samples were returned to the laboratory and stored at 4°C until they were processed. To evaluate the relationship of fine root dynamics and soil environmental factors, subsoils from a soil core sampler were sampled to measure soil pH and soil moisture content. The subsoil samples were dried in an oven for 48 h at 105°C to quantify the soil gravimetric water content. Soil pH (1:5 soil:water suspension) was determined by glass electrode.
Roots from the soil core sample were separated into <2 mm in diameter class and classified as live or dead on the basis of morphology and color with careful observation. For this study the fine root system was defined as woody and non-woody root <2 mm in diameter and their associated root tips. Roots were then dried to a constant mass at 65°C and weighed to the nearest 0.01 g. Root nutrients (N, P, K, Ca, Mg) in live and dead roots sampled for 1995 were analyzed by the standard method of National Institute of Agricultural Science and Technology (1988). All data were analyzed using the General Linear Models procedure of Statistical Analysis System (SAS Institute Inc. Citation2003).
Results and discussion
Seasonal patterns of fine root biomass
Seasonal variations in live and dead fine root biomass were found in Korean pine and sawtooth oak stands (). Both stands showed a similar seasonal pattern in live fine root biomass. Live fine root biomass generally decreased in the late growing season (September–November) with a seasonal peak in the early growing season (April–June). For example, live fine root biomass in both stands was significantly higher in April (Korean pine) or May (sawtooth oak) than in other months in 1995. The result supported that live fine root biomass in temperate forests peaks with foliage development in late spring or early summer and decreases rapidly in fall with decreasing soil temperature (Burke and Raynal Citation1994; Park et al. Citation2006; Zewdie et al. Citation2008), although other study showed no seasonal pattern of live root biomass in larch and pitch pine plantations in Korea (Hwang et al. Citation2007).
Figure 1. Monthly biomass variation of live and dead roots in a Korean pine plantation (a) and a sawtooth oak (b) stand. Vertical bar indicates one standard error.
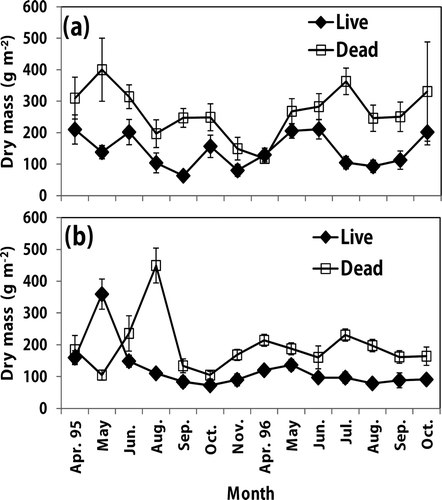
Dead fine root biomass had less seasonal pattern compared with live fine roots, while dead fine roots biomass in both stands showed generally high values in summer (July or August). Other study suggests that the seasonal pattern of fine root biomass was influenced by abiotic factors, such as soil temperature, soil moisture, and nitrogen availability (Makkonen and Helmisaari Citation1998) as well as carbon allocation mechanisms of the tree.
Fine root biomass
Fine root biomass was significantly greater for the Korean pine than for the sawtooth oak stands. Mean fine root (live, dead, total) biomass (g m−2) during the study period was 142, 266, 408 for the Korean pine plantation and 124, 193, 317 for the sawtooth oak stands, respectively (). The high live root biomass in the Korean pine plantation could be due to the difference of tree density () or the species-specific differences in carbon allocation with different stand age (Yuan and Chen Citation2010), while high dead fine roots in the Korean pine plantation may be due to a high turnover rate. In addition, high dead fine roots in the Korean pine plantation could be partly related to low soil pH values () because low soil pH could increase fine root mortality and necromass (Leuschner et al. Citation2004). Also, Kim (Citation2002) reported that fine root decompositions in Korean pine were more rapid compared with fine roots of sawtooth oak in the same study site.
Table 3. Mean biomass of live and dead fine roots, soil pH and soil moisture content in a Korean pine plantation and a sawtooth oak stand.
Fine root biomass from this study was similar to that for pine stands (377 g m−2) in temperate zone (Finér et al. Citation2011) and for the Quercus spp. (332–366 g m−2) at soil depth of 0–30 cm in Korea (Park et al. Citation2006). However, mean fine root biomass in this study was greater than the other result for the 40-year-old Japanese larch plantation (96 g m−2) in central Korea (Son and Hwang Citation2003). These differences among the studies could be due to the differences in soil sampling depth, tree species, age, and assessment methodology (Hendricks et al. Citation2006; Finér et al. Citation2011) with a large spatial and temporal variation in fine root biomass (O’Grady et al. Citation2005).
Nutrient concentration of fine roots
Nitrogen and P concentrations of live roots in both stands were little changed during growing season, while other nutrient concentrations showed seasonal fluctuations ( and ). Potassium, Ca and Mg concentrations in live fine roots were significantly higher in summer (August) than in spring (April or May) except for K in the sawtooth oak. Nutrient concentrations of dead fine root also showed seasonal fluctuations during the growing season except for P and N in the sawtooth oak stands. In addition, K, Ca and Mg concentrations in dead fine roots were significantly higher in summer or fall (August or September) than in spring (April or May). This seasonal variation of nutrients in fine roots could be due to the change of mineralization, immobilization, leaching loss and retranslocation during the growing season (Chen et al. Citation2001; Borken et al. Citation2007).
Figure 2. Monthly nutrient variation of live and dead roots in a Korean pine plantation. Vertical bar indicates one standard error.
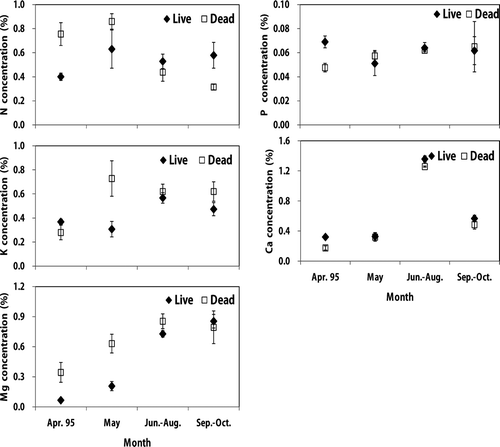
Figure 3. Monthly nutrient variation of live and dead roots in a sawtooth oak stand. Vertical bar indicates one standard error.
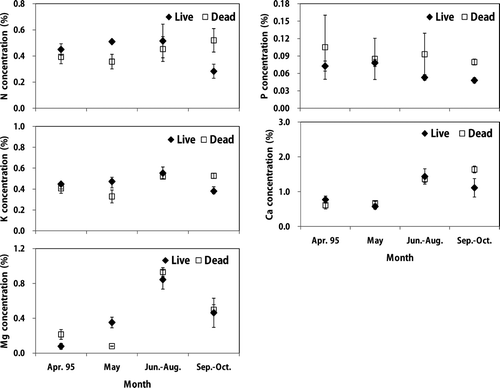
Mean nutrient concentrations between live and dead fine roots were not significantly different between the Korean pine plantation and the sawtooth oak stand (). Similarly, Gordon and Jackson (Citation2000) reported that mean N concentration by analyzing data from 56 published studies were similar between live and dead fine roots. The nutrient concentrations in live fine roots of the Korean pine plantation were N (0.59%) > Ca (0.5%) > K (0.47%) > Mg (0.42%) > P (0.07%), while the concentrations in dead fine roots were K (0.60%) > Mg (0.58%) > N (0.55%) > Ca (0.42%) > P (0.07%). The mean nutrient concentrations of live fine roots in the sawtooth oak stand followed the order: Ca (0.82%) > N (0.44%) > K (0.43%) > Mg (0.36%) > P (0.07%), while the concentrations in dead fine roots were Ca (0.80%) > K (0.47%) > N (0.45%) > Mg (0.44%) > P (0.09%). Other root studies showed nutrient concentration of N (0.81%) and P (0.05%) in pitch pine, and N (1.01%) and P (0.07%) in larch plantations (Hwang et al. Citation2007), and N (0.66%), P (0.04%), K (0.33%), Ca (0.83%), and Mg (0.16%) in oak stands in Korea (Park et al. Citation2006). Mean fine root nutrient concentrations in this study were generally within the range of values reported elsewhere for forest ecosystems (Gordon and Jackson Citation2000; Park et al. Citation2006).
Table 4. Mean nutrient concentrations of live and dead fine roots in a Korean pine plantation and a sawtooth oak stand.
Root nutrient concentrations during the study period were generally higher in the Korean pine than in the sawtooth roots except for Ca and P (). In addition, N concentration was significantly higher in the Korean pine (0.56%) than in the sawtooth oak roots (0.43%), while Ca concentration was significantly lower in the Korean pine (0.61%) than in the sawtooth oak roots (1.02%). High Ca concentrations in the oak roots may be attributed to a structure component of cell wall within plant tissues. In addition, the specific gravity of green wood in a Korean pine was 0.68 g cm−3, while that in a sawtooth oak was 1.05 g cm−3 (Kim Citation2002). In contrast, Gordon and Jackson (Citation2000) reported that nutrient concentrations in fine roots were not differed significantly between coniferous and broad-leaved trees except for Mg.
Acknowledgements
This work was supported by Gyeongnam National University of Science and Technology Grant (2012).
References
- Bolte , A and Villanueva , I . 2006 . Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies(L.) Karst.). Eur . J For Res , 125 : 15 – 26 .
- Borken , W , Kossmann , G and Matzner , E . 2007 . Biomass, morphology and nutrient contents of fine roots in four Norway spruce stands . Plant Soil , 292 : 79 – 93 .
- Burke , M and Raynal , D J . 1994 . Fine root growth phenology, production, and turnover in a northern hardwood forest ecosystem . Plant Soil , 162 : 135 – 164 .
- Chen , H , Harmon , M E and Griffiths , R P . 2001 . Decomposition and nitrogen release from decomposing woody roots in coniferous forests of the Pacific Northwest: a chronosequence approach . Can J For Res , 31 : 246 – 260 .
- Finér , L , Helmisaari , H S , Lõhmus , K , Majdi , H , Brunner , I , Børja , I , Eldhuset , T , Godbold , D , Grebenc , T , Konôpka , B , Kraigher , H , Möttönen , M R , Ohashi , M , Oleksyn , J , Ostonen , I , Uri , V and Vanguelova , E . 2011 . Variation in fine root biomass of three European tree species: beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.) . Plant Biosystems , 141 : 394 – 405 .
- Gill , R A and Jackson , R B . 2000 . Global patterns of root turnover for terrestrial ecosystems . New Phytol , 147 : 13 – 31 .
- Gordon , W S and Jackson , R B . 2000 . Nutrient concentrations in fine roots . Ecology , 81 : 275 – 280 .
- Hendricks , J J , Hendrick , R L , Wilson , C A , Mitchell , R J , Pecot , S D and Guo , D . 2006 . Assessing the patterns and controls of fine root dynamics: an empirical test and methodological review . J Ecol , 94 : 40 – 57 .
- Hwang , J , Son , Y , Kim , C , Yi , M J , Kim , Z S , Lee , W K and Hong , S K . 2007 . Fine root dynamics in thinned and limed pitch pine and Japanese larch plantation . J Plant Nutr , 30 : 1821 – 1839 .
- Jackson , R B , Moony , H A and Schulze , E D . 1997 . A global budget for fine root biomass, surface area, and nutrient contents . Proc Natl Acad Sci USA , 94 : 7362 – 7366 .
- Kim , C . 2002 . Mass loss rates and nutrient dynamics of decomposing fine roots in a sawtooth oak and a Korean pine stands . Korean J Ecol Sci , 1 : 101 – 105 .
- Leuschner , C , Hertel , D , Schmid , I , Koch , O , Muhs , A and Holscher , D . 2004 . Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility . Plant Soil , 258 : 43 – 56 .
- Makkonen , K and Helmisaari , H S . 1998 . Seasonal and yearly variations of fine-root biomass and necromass in a Scots pine (Pinus sylvestris L.) stand . For Ecol Manage , 102 : 283 – 290 .
- Majdi , H , Pregitzer , K , Morén , A S , Nylund , J E and Ågren , G I . 2005 . Measuring fine root turnover in forest ecosystems . Plant Soil , 27 : 1 – 8 .
- National Institute of Agricultural Science and Technology . 1988 . Methods of soil chemical analysis , Suwon : Sammi Press . (in Korean)
- O’Grady , A P , Worledge , D and Battaglia , M . 2005 . Temporal and spatial changes in fine root distributions in young Eucalyptus globules stand in southern Tasmania . For Ecol Manage , 214 : 373 – 383 .
- Park , G S , Lim , J G , Kim , D U and Ohga , S . 2006 . Net nine root carbon production in Quercus natural forest ecosystems of Korea . J Fac Agr Kyusu Univ , 51 : 57 – 61 .
- SAS Institute Inc . 2003 . SAS/STAT statistical software , Version 9.1. Cary , NC : SAS Publishing .
- Silver , W L and Miya , R K . 2001 . Global patterns in root decomposition: comparisons of climate and litter quality effects . Oecologia , 129 : 407 – 419 .
- Son , Y and Hwang , J H . 2003 . Fine root biomass, production and root turnover in a fertilized Japanese larch plantation in central Korea . Ecol Res , 18 : 339 – 346 .
- Yuan , Z Y and Chen , H YH . 2010 . Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: literature review and meta-analyses . Cri Rev Plant Sci , 29 : 204 – 221 .
- Zewdie , S , Fetene , M and Olsson , M . 2008 . Fine root vertical distribution and temporal dynamics in mature stands of two enset (Enset ventricosum Welw Cheesman) clones . Plant Soil , 305 : 227 – 236 .