Abstract
The biomass and carbon (C) storage of the main ecosystem components were examined in an age-sequence of six Japanese red pine forest stands in central Korea. The tree biomass was determined by the destructive method, and the C storage of the tree biomass, forest floor and mineral soil was estimated by analyzing the C concentration of each component. The above-ground and total tree biomass increased from 21.76 and 28.82 Mg ha−1 in the 17-year-old stand to 308.83 and 385.74 Mg ha−1 in the 73-year-old stand. The comparisons of above-ground tree and tree root biomass in replicate stands indicated that stand density has an effect on tree biomass partitioning for Japanese red pine, especially on the biomass allocation of above-ground tree and tree roots. The C concentrations were lowest in the tree roots while the highest concentrations were found in the foliage across the six Japanese red pine forest stands. The C storage in the forest floor and mineral soil were age-independent. The above-ground and total ecosystem C stocks increased from 19.40 and 43.49 Mg ha−1 in the 17-year-old stand to 162.72 and 247.39 Mg ha−1 in the 73-year-old stand. Although the total tree biomass C showed considerable accumulation with stand age, the relative contribution of the below-ground ecosystem to the total ecosystem C storage demonstrated large variation. The results of this study assist in understanding C storage and its change with stand development in Japanese red pine forests, which makes this species a large sink for atmospheric C at the regional scale.
Introduction
Forest biomass and soils are particularly important because both have considerable potential for temporary and long-term carbon (C) storage in terrestrial ecosystems (Houghton Citation2007; Houghton et al. Citation2009; Li et al. Citation2010a). Although the greatest potential for above-ground biomass and C storage in pine forest ecosystems is usually found within the tree biomass, the forest floor and mineral soil also provide large C stocks (Peichl and Arain, Citation2006; Taylor et al. Citation2007; Noh et al. Citation2010b; Li et al. Citation2011). Evaluating biomass C storage in forests is normally based on the carbon concentration factor (50%), which is used directly to convert the carbon storage from the biomass. However, recent analyses have shown that the C concentration of tree components or tree species might be either above or below 50% (Laiho and Laine Citation1997; Lamlom and Savidge Citation2003; Bert and Danjon Citation2006; Zheng et al. Citation2008). The accurate assessment of the forest carbon storage thus can be improved by estimating the C concentration in its each component.
Japanese red pine (Pinus densiflora Sieb. et Zucc.) is one of the most important tree species in Korea, both in terms of the high-value wood products and cultural significance, covering approximately 27% of the total forest area in Korea and playing a very important role in the national and regional ecosystem C cycles (Lee et al. Citation2004; Chun et al. Citation2006; Kim Citation2008; Li et al. Citation2010b). Previous studies on allometric equations, biomass, and biomass expansion factors for this species in Korea have been reported (Lee Citation1985; Park and Kim Citation1989; Park and Lee Citation1990; Lee et al. Citation2004; Park et al. Citation2005; Li et al. Citation2010b), including a description of the stand C dynamics following forest tending works (Kim et al. Citation2009) and the first estimates of C and nitrogen stocks in a chronosequence of Japanese red pine stands (Noh et al. Citation2010a). However, there is still a lack of information on the biomass and C storage in Japanese red pine stands, especially in replicated stands with same age class.
The aims of this study were (1) to develop site-specific allometric equations to estimate tree biomass and (2) to assess the C stocks and changes of the main ecosystem components across an age-sequence of six Japanese red pine stands in central Korea by analyzing the C concentration of each component.
Materials and methods
Site description
The study was carried out in a natural Japanese red pine forest in the Inge area (38°59′N, 128°11′E) in central Korea. Japanese red pine purely dominates in the overstorey vegetation. There has been no commercial logging, fires, or artificial management such as thinning in the forest. The climate in the study area is humid continental with hot, humid summers and cold, dry winters. The mean annual precipitation of 1172 mm is distributed mainly in summer. The average temperatures in January and July are −2.7 and 20.6 °C, respectively. The soil texture is sandy clay loam. Given that the ratios of the total area and total stem volume of two age classes (20–30 and 30–40 years old) to that of all age classes for Korean forests were 67.7% and 71.5% in 2007 (Statistical Yearbook of Forestry Citation2008), it was possible to find replicate stands for the age classes of 20–30 and 30–40 years old with similar microclimate conditions in this region (Table ). The study design consisted of six stands with four age classes. Each sample plot (20 m × 20 m) was established, and the distances between stands were less than 500 m.
Table 1 Stand characteristics of the six Japanese red pine stands in central Korea
Destructive tree sampling
Trees were sampled based on the destructive methods, which were the same as those reported by Li et al. (Citation2010b). Briefly, five Japanese red pine trees representing the stand-specific diameter at breast height (DBH) range were selected and sampled destructively in each stand in early September 2007 (Table ). The trees were cut at a height of 20 cm above the ground. Before removing the branch, the diameter of each branch was measured, and five representative branches from the smallest to the largest throughout the crown were sampled. All branches were then clipped from the tree, and fresh weights were determined using a balance in the field. Those sampled branches were separated into foliage and branches, and subsamples of each component were taken to the laboratory to determine the moisture storage. The stem of each tree was cut in 2 m sections and weighed on a balance. A disc (5 cm wide) was cut from the stump to the top of each stem section to determine the ratio of dry weight to green weight. The dry weight of each component (foliage, branch and stem) was calculated for each sample tree. For estimating tree root biomass, two trees were harvested based on a stand-specific DBH range, and the root system was washed lightly to remove soil particles, oven-dried and weighed. The total dry weight for each component (foliage, branch, stem and root) was calculated, and the weights were related to the DBH in a logarithmic regression equation (Park and Lee Citation1990; Noh et al. Citation2010a): Log10 Y = a + b Log10 (DBH), where a and b are the equation parameters, and Y is the biomass (g) of each tree component (Table ). Correction factors were used in calculating the biomass of each tree component to eliminate any systematic bias in the log transformation (Sprugel Citation1983).
Table 2 Regressions of dry weights (g) in different tree components on DBH (cm) for Japanese red pine in central Korea
Tree component samples including stem wood, bark, foliage, branch, and root were collected from each stand. The component samples were dried, ground with a laboratory grinder, and then analyzed for the C concentration. The C storage was obtained by multiplying each tissue C concentration by the total dry weight of each component.
Forest floor and soil sampling
Three forest floor (LFH-layer) and three mineral soil samples were taken from each stand.
The forest floor was sampled by collecting the entire organic material within a 0.09 m2 quadrant placed at the central point of each stand, as well as in two replicate quadrants located within a radius of 5 m. The samples were oven dried separately at 85°C to a constant weight. The three samples were mixed and ground to approximately 4 mm to reduce the particle size. The subsamples were ground in a ring grinder to produce a fine powder with a particle size of approximately 1 μm. This powder was used to determine the C concentration.
Mineral soil samples were extracted from a depth up to 50 cm with three replicates in each stand. At each sampling point, soil samples were excavated from four depths (0–10, 10–20, 20–30, and 30–50 cm) using an improved double-cylinder hammer-driven soil corer (Hwang et al. Citation2001). The bulk density for each soil depth was measured by weighing the entire sample and drying the subsamples at 105°C. After measuring the bulk density, the samples were sieved with a 2 mm sieve to separate the sample into fine and coarse fractions. The total soil C concentration was analyzed.
The C concentration of all samples (tree tissue, forest floor and mineral soil) was analyzed using a Vario Macro Elemental Analyzer (Elementar Analysensysteme GmbH, Germany).
The common software package SPSS (v16) was used to examine stand means and within-stand variations. One-way analysis of variance with Tukey's test was used to evaluate the significant differences between the C concentrations of tree components and forest floor. Unless stated otherwise, a p value <0.05 was considered significant.
Results and discussion
Allometric equations, biomass and C concentration
Using a simple linear regression analysis with a log-transformation of the independent and dependent variables, the allometric equations were produced for the tree component biomass (Table ). For all equations, the correction factors were calculated using the standard error of the estimate, which was converted from base 10 to e-base to obtain the correct value (Sprugel Citation1983). The allometric equations that were developed explained more than 90% of the variability for the tree components except for branch. The equations were most fitted for the above-ground and total tree biomasses among the various tree components (p < 0.0001). The results from previous studies using a single diameter-based equation for Japanese red pine in Korea have shown that foliage and branch biomass are more difficult to calculate compared with other above-ground tree components (Park and Kim Citation1989, Park and Lee Citation1990; Noh et al. Citation2010b). For example, R2 values ranging from 0.764 to 0.986 were reported for branch biomass in four local forms of Japanese red pine throughout Korea (Park and Kim Citation1989; Park and Lee Citation1990), whereas the R2 value of 0.887 was reported for foliage biomass in a chronosequence of Japanese red pine stands (Noh et al. Citation2010a).
Using the logarithmic regression equations, above-ground tree and total tree biomass in the six Japanese red pine stands were estimated (Table ). The above-ground tree and total tree biomass ranged from 21.76 and 28.82 Mg ha−1 in the 17-year-old stand to 308.83 and 385.74 Mg ha−1 in the 73-year-old stand, demonstrating a rapid increase with stand age. The proportion of stem biomass to the above-ground tree biomass ranged from 52% for the 17-year-old stand to 78% for the 73-year-old stand, whereas the proportions between the replicate stands were different. Tree root biomass in the 26-year-old stand was approximately twice the amount of that in the 28-year-old stand while above-ground tree biomass seemed to be similar in both stands. In contrast, above-ground tree and tree root biomasses in the 38-year-old stand were nearly twice the amount of those in the 36-year-old stand. Stand densities differed obviously in the 26- and 28-year-old stands whereas there was no main difference in the stand characteristics of the 38- and 36-year-old stands, which may indicate that stand density has an effect on tree biomass partitioning for Japanese red pine. Site-specific conditions, such as the stand density, altitude and slope, affect the future biomass allocation to above-ground tree and tree root in the Japanese red pine stands with the same age. Although our findings lack replication information from the 17- and 73-year-old stands, previous studies have reported similar results (Park and Lee Citation1990; Kim et al. Citation2009; Noh et al. Citation2010b). Noh et al. (Citation2010b) designed a chronosequence of Japanese red pine stands in Korea, and reported that the above-ground and total tree biomass increased steadily with stand age. Park and Lee (Citation1990) reported that above-ground tree biomass in 33- and 31-year-old stands of Japanese red pine forests with different stand characteristics were 93.55 and 116.61 Mg ha−1 while tree root biomass in the 33- and 31-year-old stands were 15.35 and 15.00 Mg ha−1. Changes in stand density following forest tending works in a 40-year-old Japanese red pine forest influenced significantly tree biomass C dynamics (Kim et al. Citation2009). Stand density, as one of the key factors, changed biophysical and environmental factors in Japanese red pine stands (Noh et al. Citation2010a).
Table 3 Tree biomass and forest floor (Mg ha−1) in the six Japanese red pine stands
Since most forest inventories focus on the stem biomass or timber volume, the varied proportion of stem biomass to the above-ground tree biomass in the six Japanese red pine stands may indicate that above-ground tree biomass, especially in a young stand, may be underestimated by the forest inventories, which suggests that there is a need to develop age-related biomass expansion factors in order to estimate other tree components, such as twig, branch and foliage (Park et al. Citation2005; Li et al. Citation2010b). Although the tree root biomass increased steadily over time (Table ), the ratio of tree root to above-ground tree biomass varied significantly from 0.14 to 0.32 across the six Japanese red pine stands. The biomass allocation of tree roots was strongly influenced by different site conditions and ecotypes (Park et al. Citation2005; Noh et al. Citation2010b).
The C concentrations of the tree components were significantly different (p < 0.05) (Table ). On average, the C concentrations were lowest in the tree roots while the highest concentrations were found in the foliage with a mean value of 51.88% across the six Japanese red pine stands. Similar results of C concentration analysis from a chronosequence stand of same species showed that foliage had higher values than tree roots (Noh et al. Citation2010b). The C concentrations varied largely among tree components, and may be affected by a range of factors, such as sample chemical composition, stand age and analysis means (Bert and Danjon Citation2006).
Table 4 Carbon concentration (%) of tree and forest floor in the six Japanese red pine stands (stand mean ± within-stand S.D.)
Tree biomass C, forest floor C and mineral soil C
Using the results of the biomass and mean C concentration (Tables and ), the C stocks in the tree components can be calculated. The above-ground and total tree biomass C stocks increased from 10.83 and 14.15 Mg ha−1 in the 17-year-old stand to 154.38 and 190.64 Mg ha−1 in the 73-year-old stand. The biomass C storage in each tree component increased steadily with stand age. The contribution of stem biomass C storage to total tree biomass C storage increased from 39% for the 17-year-old stand to 63% for the 73-year-old. For a better understanding of accurate C assessment in tree biomass, the carbon concentration factor (50%) was used for estimating the predicted C storage from the tree biomass to compare it with the observed C storage by analysis. The relative difference to compare the values between the predicted and observed C storage varied from one tree component to another. Overall, the predicted C stocks were generally higher than the observed C stocks in the tree components except for foliage, which indicated that the 50% value is an oversimplification when dealing with the C storage in forests (Lamlom and Savidge Citation2003).
The C stocks in the forest floor for the 17-, 28-, 26-, 36-, 38-, and 73-year-old stands was 8.57, 6.54, 8.21, 15.33, 9.58 and 8.34 Mg ha−1, respectively (Table ). The C stocks of the forest floor in our 28-, 26- and 73-year-old stands were lower than 10.0 and 11.2 Mg ha−1 of the forest floor in the 27- and 71-year-old stands reported for the same species in Korea (Noh et al. Citation2010b). However, there was no age-related increase in our results, which were in accordance with the results from a chronosequence of Japanese red pine stands (Noh et al. Citation2010b). The C stocks of the forest floor in two ∼ 40-year-old Japanese red pine stands were not changed by forest tending works (Kim et al. Citation2009). The discrepancy between the different findings was due mainly to the large inter- and intra-stand spatial variation (Peilchl and Arain Citation2006; Taylor et al. Citation2007).
Table 5 Above-ground and below-ground ecosystem C stocks (Mg ha−1) in the six Japanese red pine stands
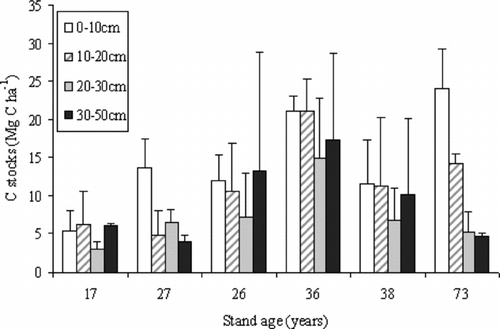
With increasing soil depth, the C concentrations of the mineral soil for each stand decreased from approximately 2.32% at 0–10 cm depth to 0.62% at 30–50 cm depth (data not shown). The total C storage of mineral soil in the 17-, 28-, 26-, 36-, 38-, and 73-year-old stands was 20.77, 29.03, 43.21, 74.46, 39.99 and 48.41 Mg ha−1, respectively (Table ). The total C storage of mineral soil in this study did not show any age-related patterns (Figure ), which was close to the results from a chronosequence of Japanese red pine stands (Noh et al. Citation2010b). The total C storage of mineral soil in each stand was lower than the 82.3 Mg ha−1 in the overall soil of temperate forests across all age classes (Pregitzer and Euskirchen Citation2004). The upper 20 cm of mineral soil seemed to sequestrate more C stocks than a deeper depth for each stand. Although an improved soil corer was used to obtain better bulk densities in this study, soil samples from a deeper depth may be disturbed by the coring compaction, which would further affect the absolute values of mineral soil C.
Figure 1 The C stocks of the different mineral soil depth in the six Japanese red pine stands (error bars indicate within-stand S.D.)
Total ecosystem C
The above-ground and total ecosystem C stocks increased with stand age (Table ). The increasing trend of total ecosystem C storage in our stands was different from the sigmoidal pattern examined for the same species (Noh et al. Citation2010b). Different patterns were mainly due to deeper soil excavation depth and smaller chronological stages in our case compared with the previous chronological studies (Taylor et al. Citation2007; Noh et al. Citation2010b). The ratio of below-ground to above-ground ecosystem C storage decreased from 1.24 in the 17-year-old stand to 0.52 in the 73-year-old stand, whereas the ratios in replicate stands were different. The mean ratio was 1.04 for the 28- and 26-year-old stands, and 1.01 for the 36- and 38-year-old stands. If using the mean ratios, this decreasing ratio may suggest the fast C accumulation of the tree biomass in the 28- and 26-year-old stands, and a large C sequestration potential in the 73-year-old stand with the stand's maturing development. Similar decreasing ratios were also reported in an age-sequence of four White pine plantation stands (Peilchl and Arain Citation2006).
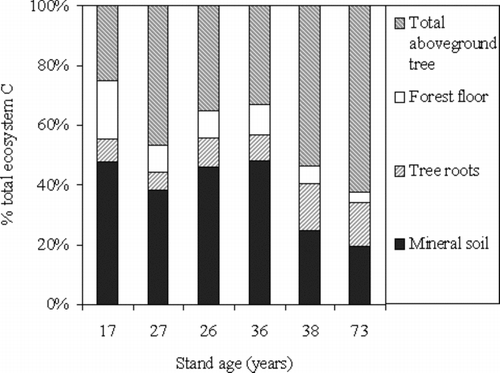
The contribution of mineral soil to total ecosystem C storage decreased from 48% in the 17-year-old stand to 20% in the 73-year-old stand (Figure ). The contribution of above-ground tree biomass to total ecosystem C storage increased from 25% in the 17-year-old stand to 62% in the 73-year-old stand, whereas the contribution in replicate stands was different. The forest floor C storage comprised 3–20% of the total ecosystem C storage. With increasing stand age, the relative contribution of below-ground ecosystem (tree roots plus mineral soil) to total ecosystem C storage seemed to be variable with a range of 34–55% in this study. Similar results were also found in other chronosequence studies. For example, a fairly variable range of 35–90% was found in a chronosequence of Korean pine plantation stands (Li et al. Citation2011), whereas a slightly unstable range of 45–50% was reported in the four different-aged boreal mixed-wood stands (Martin et al. Citation2005). Different comparisons indicated that total ecosystem C storage may depend on the below-ground C storage because of the instability, whereas above-ground tree biomass may make major contribution to total ecosystem C development (Peilchl and Arain Citation2006; Li et al. Citation2011). Therefore, a more accurate estimation of C storage of mineral soil is needed in future chronosequence studies.
Figure 2 The C stocks of above-ground tree biomass, forest floor, tree roots, and mineral soil in the six Japanese red pine stands expressed as a percentage
Conclusions
Using the allometric equations developed from limited samples in this study, above-ground and total tree biomass increased with stand age. The comparisons of above-ground tree and tree root biomass in replicate stands indicated that stand density has an effect on tree biomass partitioning for Japanese red pine, especially on the biomass allocation of above-ground tree and tree roots. The C concentrations were lowest in the tree roots while the highest concentrations were found in the foliage. The C storage in the forest floor was no age-related increase, which was due mainly to the large inter- and intra-stand spatial variation. The total mineral soil C storage was age-independent in our study. The upper 20 cm of mineral soil sequestrated more C stocks than a deeper depth for each stand, whereas the absolute values of mineral soil C storage from a deeper depth may be affected by the coring compaction. The above-ground and total ecosystem C stocks increased with stand age, from 19.40 and 43.49 Mg ha−1 in the 17-year-old stand to 162.72 and 247.39 Mg ha−1 in the 73-year-old stand. The total tree biomass C showed considerable accumulation with stand age. However, the relative contribution of the below-ground ecosystem to the total ecosystem C storage demonstrated large variation. Priority should be focused on the accurate estimation of mineral soil C storage in future chronosequence studies.
Acknowledgement
This study was carried out with the support of “Forest Science & Technology Projects (No. S1107L0101)” provided by Korea Forest Servce.
Notes
aMSR is the mean square residuals. bCF is the correction factor (Sprugel Citation1983). Equations follow the form Log10 Y = a+b Log10 X, where X is DBH, Y is the dry weight of different components.
Notes: Means with different letters within columns are statistically different at p < 0.05.
References
- Bert , D and Danjon , F . 2006 . Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.) . Forest Ecol Manag , 222 : 279 – 295 . doi: 10.1016/j.foreco.2005.10.030
- Chun , Y M , Lee , H J and Lee , C S . 2006 . Vegetation trajectories of Korean red pine (Pinus densiflora Sieb. et Zucc.) forests at Mt. Seorak, Korea . J Plant Biol , 49 : 141 – 152 . doi: 10.1007/BF03031010
- Houghton , R A . 2007 . Balancing the global carbon budget . Annu Rev Earth Pl Sci. , 35 : 313 – 347 . doi: 10.1146/annurev.earth.35.031306.140057
- Houghton , R A , Hall , F and Goetz , S J . 2009 . Importance of biomass in the global carbon cycle . J Geophys Res , 114 : G00E03 doi: 10.1029/2009JG000935
- Hwang , J H , Son , Y H and Kim , J S . 2001 . An improved soil core sampler . J Korean Forest Soc. , 90 : 788 – 791 .
- Kim , C S . 2008 . Soil CO2 efflux in clear-cut and uncut red pine (Pinus densiflora S. et. Z.) stands in Korea . Forest Ecol Manag , 255 : 3318 – 3321 . doi: 10.1016/j.foreco.2008.02.012
- Kim , C S , Son , Y H , Lee , W-K , Jeong , J Y and Noh , N-J . 2009 . Influences of forest tending works on carbon distribution and cycling in a Pinus densiflora S. et Z. stand in Korea . Forest Ecol Manag , 257 : 1420 – 1426 . doi: 10.1016/j.foreco.2008.12.015
- Laiho , R and Laine , J . 1997 . Tree stand biomass and carbon storage in an age sequence of drained pine mires in southern Finland . Forest Ecol Manag , 93 : 161 – 169 . doi: 10.1016/S0378-1127(96)03916-3
- Lamlom , S H and Savidge , R A . 2003 . A reassessment of carbon storage in wood: variation within and between 41 North American species . Biomass Bioenerg , 25 : 381 – 388 . doi: 10.1016/S0961-9534(03)00033-3
- Lee , S W . 1985 . Biomass and net primary productivity of Pinus densiflora natural ecosystem in Kangwon, Korea . J Korean Forest Soc. , 71 : 74 – 81 .
- Lee , W K , Gadow , K V , Chung , D J , Lee , J L and Shin , M Y . 2004 . DBH growth for Pinus densiflora and Quercus variabilis mixed forests in central Korea . Ecol Model , 176 : 187 – 200 . doi: 10.1016/j.ecolmodel.2003.11.012
- Li , X D , Yi , M J , Son , Y H , Jin , G Z and Han , S S . 2010a . Forest biomass carbon accumulation in Korea from 1954 to 2007 . Scand J Forest Res. , 25 : 554 – 563 . doi: 10.1080/02827581.2010.524892
- Li , X D , Yi , M J , Son , Y H , Park , P S , Lee , K H , Son , Y M , Kim , R H and Jeong , M J . 2010b . Biomass expansion factors of natural Japanese red pine (Pinus densiflora) forests in Korea . J Plant Biol , 53 : 381 – 386 . doi: 10.1007/s12374-010-9134-7
- Li , X D , Yi , M J , Son , Y H , Park , P S , Lee , K H , Son , Y M , Kim , R H and Jeong , M J . 2011 . Biomass and carbon storage in an age-sequence of Korean pine (Pinus koraiensis) plantation forests in central Korea . J Plant Biol , 54 : 33 – 42 . doi: 10.1007/s12374-010-9140-9
- Martin , J L , Gower , S T , Plaut , J and Holmes , B . 2005 . Carbon pools in a boreal mixed wood logging chronosequence . Global Change Biol , 11 : 1883 – 1894 .
- Noh , N J , Son , Y H , Lee , S K , Seo , K W , Heo , S J , Yi , M J , Park , P S , Kim , R H , Son , Y M and Lee , K H . 2010a . Carbon and nitrogen storage in an age-sequence of Pinus densiflora stands in Korea . Sci China Life Sci , 53 ( 7 ) : 822 – 830 . doi: 10.1007/s11427-010-4018-0
- Noh , N J , Son , Y H , Lee , S K , Yoon , T K , Seo , K W , Kim , C S , Lee , W K , Bae , S W and Hwang , J H . 2010b . Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea . J Plant Res. , 123 ( 4 ) : 411 – 419 . doi: 10.1007/s10265-010-0331-8
- Park , I H and Kim , J S . 1989 . Biomass regression of Pinus densiflora natural forests of four local forms in Korea . J Korean Forest Soc. , 78 : 323 – 330 . (in Korean with English abstract)
- Park , I H and Lee , S M . 1990 . Biomass and net production of Pinus densiflora natural forests of four local forms in Korea . J Korean Forest Soc. , 79 : 196 – 204 . (in Korean with English abstract).
- Park , I H , Park , M S , Lee , K H , Son , Y M , Son , J H , Son , Y H and Lee , Y J . 2005 . Biomass expansion factors for Pinus densiflora in relation to ecotype and stand age . Jour Korean Forest Soc. , 94 : 441 – 445 . (in Korean with English abstract)
- Pregitzer , K S and Euskirchen , E S . 2004 . Carbon cycling and storage in world forests: biome patterns related to forest age . Global Change Biol , 10 ( 12 ) : 2052 – 2077 . doi: 10.1111/j.1365-2486.2004.00866.x
- Peichl , M and Arain , M A . 2006 . Above- and belowground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests . Agr Forest Meteorol , 140 ( 1-4 ) : 51 – 63 . doi: 10.1016/j.agrformet.2006.08.004
- Sprugel , D G . 1983 . Correcting for bias in log-transformed allometric equations . Ecology , 64 ( 1 ) : 209 – 210 . doi: 10.2307/1937343
- Statistical Yearbook of Forestry . 2008 . Seoul: Korea Forest Service
- Taylor , A R , Wang , J R and Chen , H YH . 2007 . Carbon storage in a chronosequence of red spruce (Picea rubens) forests in central Nova Scotia, Canada . Can J Forest Res , 37 ( 11 ) : 2260 – 2269 . doi: 10.1139/X07-080
- Zheng , H , Ouyang , Z , Xu , W , Wang , X , Miao , H , Li , X and Tian , Y . 2008 . Variation of carbon storage by different reforestation types in the hilly red soil region of southern China . Forest Ecol Manag. , 255 : 1113 – 1121 . doi: 10.1016/j.foreco.2007.10.015