
Abstract
This study was carried out in order to explore and compare the correlation between each measured item of the above-ground and below-ground biomass of 13-year-old Pinus densiflora S. et Z. and prepare regression equations to identify differences of early growth characteristics on each slope aspect in the stand. The south-facing slope showed the highest above-ground and below-ground biomass, followed by the north-facing slope and ridge area. The root collar diameter showed the highest correlation between the biomass of the above-ground part and the below-ground part. In the linear regression equation on the root collar diameter and the biomass of the above-ground part and the below-ground part, the growth gradient was high in the south-facing slope and north-facing slope and low in the ridge area such that the biomass in the ridge area may well continue to be low in the future as the trees grow. The result of the study demonstrated that the differences in the growth environments associated with differences in the slope aspect had a significant influence on the early growth of trees. The results of this study further manifested the importance of the concept of slope direction in artificial forestation and could be used as preliminary data to support artificial forestation measures in the future.
Introduction
The relative balance between the above-ground and below-ground biomass of a tree is decided by the internal hereditary characteristics and the influence of external environmental factors (Köstler et al. Citation1968, Santantonio Citation1990, Lacointe Citation2000). In particular, the biomass of species with the same hereditary characteristics may also change according to environmental factors. This has been proven by preceding studies on the change of root weight according to light conditions (Röhrig Citation1966, Ledig et al. Citation1970), and the diverse influence of other environmental factors (Gruber Citation1994).
The biomass of the above-ground and the below-ground parts can be estimated easily using the diameter at breast height and root collar diameter as supply variables, and the relative growth-estimating equation using such a statistical method can apply to actual forestry fields easily (Lee, Citation2004a). However, regional characteristics according to lumbering conditions, financial burdens and the distribution of relevant tree species should be considered when investigating the biomass of a tree (Whittaker & Marks Citation1975, Alban et al. Citation1978, Lee et al. Citation1985, Park & Lee Citation1990, Bartelink Citation1998, Drexhage & Gruber Citation1999, Le Goff & Ottorini Citation2001).
Harrington (Citation1979) estimated the regression equation that could calculate the relevant carbon dioxide absorption volume using the tree height or diameter, which could be measured easily by paying attention to the fact that the biomass of the above-ground and below-ground parts had a very close correlation with the tree height and diameter, and various regression equations regarding diverse tree species have been studied since then (Na et al. Citation2011). The method for estimating biomass according to the correlation between the above-ground and below-ground biomass using the regression equation has the disadvantage that it requires a lot of time and effort to study, but due to its high reliability and economic feasibility, it is being used frequently (Lee Citation2004a).
Pinus densiflora S. et Z. is highly adaptable to the environmental conditions of Korea such that it has been distributed widely throughout the country, but it shows differences in biomass even within the same age-class according to the environmental conditions (Byun et al. Citation2010, Kim et al. Citation2012). Lee (Citation2004b) reported that the development of vertical roots slowed down and the development and growth of horizontal roots was enhanced when P. densiflora S. et Z. grew in poor soil conditions, which significantly reduced the biomass of the above-ground part. Lee et al. (Citation2005) reported that significant differences in annual ring width occurred in Pinus rigida Mill. of similar age and soil conditions due to differences in slope direction.
It has been reported that the slope direction is an important factor which determines the light conditions, temperature, humidity and physical and chemical characteristics of soil, and it is closely related to the growth of biota and trees so that the biomass of the same species of trees vary 5–30 times according to the slope direction (Kutiel Citation1992, Allan & Ian Citation2003), but the studies regarding such relationship between the slope direction and the growth of trees have been carried out mostly based on north-facing and south-facing slopes (Stage Citation1976, Atsushi et al. Citation1993, Scott & Hawkins Citation2005). Especially, studies regarding the correlation between the above-ground and below-ground biomass based on the concept of slope direction in Korea are significantly insufficient.
Therefore, the purpose of this study was to expose the influence of environmental differences such as growth environments and slope directions on the biomass growth of the above-ground and below-ground parts of P. densiflora S. et Z. The aim was to explore a comparative analysis of correlation between the above-ground and below-ground parts of a planted 13-year-old P. densiflora S. et Z. stand according to the slope direction.
Materials and methods
Study area and selection of the target trees stand
This study was carried out in Geombongsan Mountain, Imwon-ri, Wonduk-eup, Samcheok, Gangwon-do (37°13ʹ04.08ʹʹN, 129°18ʹ05.77ʹʹE), which is among the areas damaged by forest fire in April 2000 on the east coast of Korea. The study targeted the stand of II age-class P. densiflora S. et Z. (13 years old). The investigation was carried out separately on the south-facing slope, the north-facing slope and the ridge area in order to compare the growth increment of trees in the stand of P. densiflora S. et Z. in the target area. The target trees were 1-0 P. densiflora S. et Z. containerized seedlings planted in 2001, from which seven 13-year-old seedlings on each slope were selected in October 2013, which was the resting stage. At the time, only the target trees with the average height and diameter which represented the stand were selected, and a distance of 5 m between target trees was given to prevent the interference of growth between target trees.
The general conditions of the target area are shown in . The sea-level altitude was 270–277 m and there was a significant difference in slope inclination between slopes. The pH of soil on each slope ranged between 4.8 and 5.2 and showed no major difference. The main positive ions in the soil were shown in order of Ca2+, Mg2+, K+ and Na+, which were similar to the average distribution of positive ions in forest soils in Korea (Lee Citation1981). However, there was a major difference in rock volume between slopes ().
Table 1. General condition of the study site.
Table 2. Physical and chemical properties of soils in the studied stands.
Pinus densiflora S. et Z. was the dominant species on the target stand due to artificial forestation after the forest fire, and it was the 13-year-old P. densiflora S. et Z. with low average height which was the II age-class stand. The main understorey vegetation in the target areas included Quercus mongolica, Q. acutissima, Q. dentata, Lespedeza cyrtobotrya, Pteridium aquilinum and Smilax china.
Data collection and instrumentation
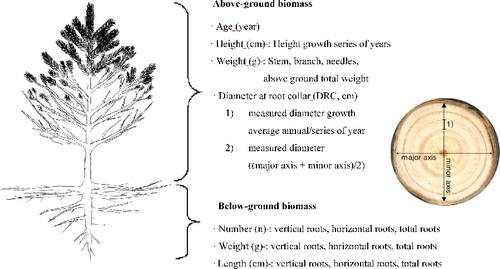
In this study, the above-ground and below-ground parts of all selected trees were investigated in order to accurately estimate the biomass even though it was costly and time-consuming. At each slope, the tree height, root collar diameter, volume growth and dry weight of needle leaf, stem and branch were harvested and measured and represented the above-ground biomass. For the below-ground biomass, the number of roots, weight and length of vertical and horizontal roots for each layer, and the length and weight of the main vertical root and the weight of fine roots were harvested within a 1.5 m radius and measured. All specimens used in the measurement were dried in a drying machine for 72 h at 104 °C, and a digital scale was used for the weight measurement.
The height of the selected trees was measured from the base to the top of the stem using a tape measurement device, and the root collar diameter was measured from the diameter of 0 plate of stem using the isoangle method. The volume growth was measured using Smalian’s formula by collecting a circular plate from the step at a 50-cm unit. The below-ground part was collected with soil within a 1.5 m radius from the root collar diameter in consideration of tree height, and all roots were collected after completely removing the soil. Then, the number, weight and length of roots, excluding roots stock and fine roots, were measured by dividing roots into a 10-cm unit for verticality and a 20-cm unit for horizontality. In addition, the weight was measured up to 0.1 g unit and the length was measured up to 0.1 cm unit ().
Figure 1. Schematic picture of the traits that were measured for the above- and below-ground biomass.
Data analysis
The correlation analysis (Pearson Bivariate Correlation) was carried out for the collected data using the statistical program SPSS (v. 21.0). The linear regression equation was estimated by setting the root collar diameter for the independent variable (X) and stem weight, branch weight, needle leaf weight, total biomass of above-ground part and total root biomass of below-ground part for dependent variables (Y). Y = β1 X + β0 (linear regression); where β0 is the Y intercepts and β1 is a slope.
Results and discussion
Analysis of biomass
The results for each part of the above-ground biomass that was investigated for three slopes showed different growth characteristics (). The average tree height of the selected trees was highest in north-facing slope (286 ± 4.82 cm), but it was not significantly different from that of the south-facing slope (275 ± 7.66 cm; p = 0.199), while the average tree height in the ridge area was 212 ± 5.36 cm and was significantly different from those of the other two slopes (p < 0.01). The average root collar diameter was shown in the order of 7.4 ± 0.45 cm on the south-facing slope and 7.1 ± 0.38 cm on the north-facing slope, but the difference was not statistically significant (p = 0.124), while the average root collar diameter in the ridge area of 5.7 ± 0.34 cm was significantly different from those of the other two slopes (p < 0.01). In consideration of the characteristics that the soil depth on the ridge area is relatively shallower and drier and contains more pore spaces than the other slopes (Kong & Lim Citation2008, Cho et al. Citation2012), it is expected that the trees growing on the ridge area under such soil conditions have low development of roots which led to lower height growth. This result is similar to that of previous studies by Lee et al. (Citation2009) and Park et al. (Citation2010), which showed that the height growth of a stand of P. densiflora S. et Z. planted on the ridge area was relatively lower compared to other slope conditions.
Table 3. Investigated variables of the above-ground biomass on different slope conditions.
The weight of the stem was shown to be of the order of 2908.6 ± 276.40 g on the south-facing slope, 2322.4 ± 224.60 g on the north-facing slope and 1478.1 ± 182.88 g in the ridge area, and among these, the ridge area showed a statistically significant difference in comparison to the other two slopes (p < 0.01). The weight of the branch was shown to be of the order of 888.9 ± 72.12 g on the south-facing slope, 775.3 ± 95.62 g on the north-facing slope and 578.8 ± 87.62 g in the ridge area. Similarly, the difference in branch weight between the north-facing and south-facing slopes was not statistically significant (p = 0.058), but both showed a significant difference in comparison to that of the ridge area at 5% level of significance. The weight of needle leaf was shown to be of the order of 747.7 ± 44.87 g on the south-facing slope, 607.6 ± 52.20 g on the north-facing slope and 472.9 ± 39.74 g in the ridge area, and the ridge area showed a statistically significant difference in comparison to the other two slopes (p < 0.01). The total biomass of the above-ground parts was shown to be of the order of 4545.2 ± 365.00 g on the south-facing slope and 3705.3 ± 339.03 g on the north-facing slope, and showed no statistically significant difference between these two slopes (p = 0.090); that of the ridge area was 2529.7 ± 284.04 g and was significantly different from those of the other two slopes (p < 0.01). It is considered that such a difference in the biomass of the above-ground parts was caused by differences in habitat environments such as light condition and soil condition associated with the slope direction.
Meanwhile, the result for each part of the biomass of the below-ground part investigated in the three slope directions is presented in . The number, weight and length of roots, excluding stump are shown in the order of south-facing slope, north-facing slope and ridge area, and the ridge area showed a significant difference at 1% significance level for the measured weight of root in comparison to the other two slopes (p < 0.01). The length and weight of stump is also shown in the order of south-facing slope, north-facing slope and ridge area, and the south-facing slope showed a significantly higher value at 1% significance level (p < 0.01). Average total biomass of the below-ground part including stump was shown to be of the order of 1473.8 ± 95.79 g on the south-facing slope, 1138.9 ± 120.49 g on the north-facing slope and 674.4 ± 68.62 g in the ridge area and a significant difference was shown between slopes (p < 0.01). Such difference in the results of the biomass of the below-ground part is of the same order as that of the biomass of the above-ground part, and is in line with the result of a previous study (Harrington Citation1979; Seo et al. Citation2010), which showed that the biomass of the above-ground part is closely related to the biomass of the below-ground part.
Table 4. Investigated variables of the below-ground biomass in different slope conditions.
The results obtained on the biomass of the above-ground and below-ground parts showed that the south-facing slope had the highest total biomass while the ridge area had the lowest total biomass. It is determined that this was caused by differences in soil condition, light condition and degree of inclination inherently associated with differences in habitat environments and slope directions.
Correlation between biomass of the above-ground and the below-ground parts
The derived correlation between the biomass of the above-ground and the below-ground parts is presented in . The correlation between each item of the above-ground part (tree height, root collar diameter, weight of stem, weight of branch, weight of needle leaf, total weight of above-ground part) and each item of the below-ground part (number, length, weight of roots, length and weight of stump, total weight of below-ground part) was mostly positive (r > 0).
Table 5. Correlation coefficient among above- and below-ground traits in different slope conditions.
The correlation between the total biomass of the above-ground part and the weight of stem and branch was significantly high in all slope directions, but the correlation between the total biomass of above-ground part and the weight of needle leaf was relatively low on the south-facing slope. This means that the weight of needle leaf accounted for a small portion in the biomass of above-ground part on the south-facing slope. According to a previous study (Qishui & John Citation1995), the amount of 2-year-old fallen needle leaves and branches on the south-facing slope was larger in comparison to other slopes and varied depending on the amount of sunshine.
Various correlations between the measurement items of the above-ground and the below-ground parts were shown to be dependent on the slope. Generally, tree height and the diameter of the tree have a high correlation with total biomass (Harrington Citation1979, Lee Citation2004a, Na et al. Citation2011), but the tree height had a low correlation with most other measurement items in this study. This may be because the selected trees of 13-year-old P. densiflora S. et Z. had height growth that was not yet well advanced. It is known that the ratio of underground to above-ground biomass decreases as the age of a tree increases, but a significant difference is shown according to the type of tree and growing environments (Marler & Wilis Citation1996). In our study, the age of the trees could not be used to estimate the ratio of underground biomass to the above-ground biomass because the sample trees were of the same age and species, but the different growing environments of the trees provided appreciable information on differences in biomass increment associated with different environmental and slope conditions.
On the other hand, the root collar diameter had a high correlation with all measurement items of the below-ground part, including the measurement items of the above-ground part such as the weights of stem, branch and needle leaf ( and ). Therefore, because in this study the root collar diameter showed a stronger positive correlation with all the measurement parts of the below-ground biomass compared to the well-documented tree height and diameter at breast height variables, we suggested that it was appropriate to calculate the regression equation for estimating the biomass of P. densiflora S. et Z. through the root collar diameter and biomass.
Figure 2. Relationship between diameter at root collar and above-ground biomass for the 13-year-old Pinus densiflora on different slope conditions.
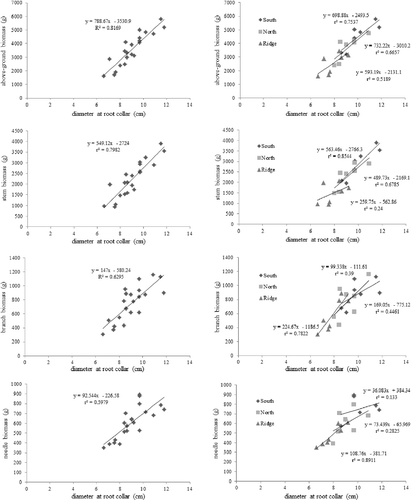
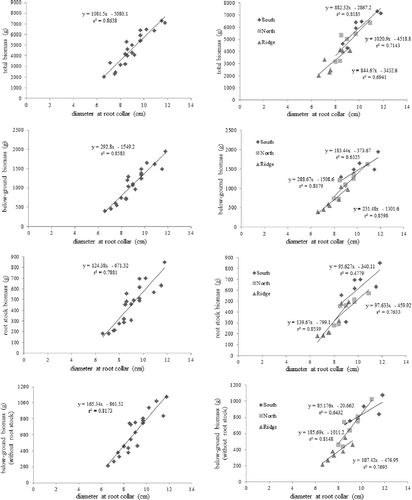
Figure 3. Relationship between diameter at root collar and below-ground biomass for the 13-year-old Pinus densiflora on different slope conditions.
Correlation between biomass of the above-ground and the below-ground parts and root collar diameter
In the analysis of correlation between the above-ground and the below-ground parts, the root collar diameter showed a higher correlation with all other measured items in comparison to the other measured items. Therefore, the linear regression equation was calculated using the root collar diameter as the independent variable (X) and the above-ground and below-ground parts as the dependent variables (Y).
The linear regression equation on the root collar diameter and the measured items of above-ground part is shown in , and the linear regression equation of the root collar diameter, the measured items of the below-ground part and the total biomass of the below-ground part is shown in . In the linear regression equation of the root collar diameter and the total biomass of the above-ground part, the growth gradient on each slope showed no significant difference between the south-facing and the north-facing slopes (). It is determined and suggested that the biomass may increase continuously at a similar rate in the future, but the observed difference between the biomass of the ridge area and the biomass of the other two slopes may increase with time as the trees grow. In the linear regression equation on the root collar diameter and the total biomass of the below-ground part, the growth gradient on each slope was higher in the order of north-facing slope, ridge part and south-facing slope. This was because the diameter growth was more promoted than the height growth in the south-facing slope with best light conditions. Therefore, in order to obtain more accurate linear regression equation, it is necessary to measure and analyze the trees when the stand becomes older.
Also, when the regression equations of the above-ground and below-ground parts and on the root collar diameter were compared, the above-ground part showed a higher growth gradient than the below-ground part. This means that the above-ground part has more biomass than the below-ground part when the diameter grows, and this result was similar to that of previous studies on Abies koreana WILS (Lee et al. Citation2007), P. densiflora S. et Z. (Lee Citation2004a) and P. densiflora and P. koraiensis (Jo et al. Citation2013).
On the other hand, in the linear regression equation on the root collar diameter and the total biomass of the above-ground and below-ground parts, the growth gradient on each slope was higher in the order of north-facing slope, south-facing slope and the ridge area. According to Kim and Lee (Citation2000) and Cho et al. (Citation2012), when a plant grows in low luminous intensity, height growth is promoted to secure the amount of light received. Our result was seen because the diameter growth on the north-facing slope has not been promoted yet.
The result of this study shows that the root collar diameter had a high correlation with the total biomass. This result is in line with that of previous studies (Harrington Citation1979, Lee Citation2004a, Na et al. Citation2011), which reported that the height and diameter of a tree such as P. densiflora S. et Z. have a high correlation with total biomass, and the method using the root collar diameter for estimating the biomass of tree at an early growth stage of less than II age-class is the most appropriate method.
Conclusions
This study was carried out in order to explore and compare the early growth characteristics and correlation between the above-ground and below-ground biomass of 13-year-old P. densiflora S. et Z. associated with three slope aspects. The study result showed that among the measured items of the above-ground biomass, the root collar diameter had the highest correlation with the below-ground part, and was thus used as the independent variable to calculate the regression equation of the biomass of the above-ground and below-ground parts.
Various regression equations were shown according to the measured items, but the biomass shown was generally highest in the order of south-facing slope, north-facing slope and ridge area. Also, the regression equation of total biomass according to the diameter of root collar showed no significant difference in the growth gradient on all slopes. Thus, the study suggests that the observed biomass increment highest in the order of south-facing slope, north-facing slope and ridge area may continue in the future as the trees grow.
However, the ridge area showed a high growth gradient in most regression equations, and this means that the development of below-ground biomass in the ridge area was more promoted in comparison to the growth of root collar diameter, and this may be because of the young age of the study stand. Therefore, it is expected that more accurate analysis of the growth gradient could be obtained when the trees in the different stand aspects must have developed properly according to the environmental conditions in the future.
The result of this study shows that the difference in the growth environments according to the slope aspects continuously influences the growth of stands in the early stage, as well as at later stages of development. This result provides compelling evidence that the concept of slope direction is invaluable in artificial forestation such that it is expected that the results could be used as preliminary data for designing future forestation plans.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Alban DH, Pelata DA, Schlaegel BE. 1978. Biomass and nutrient distribution in aspen, pine and spruce stands on the same soil type in Minnesota. Canadian Journal of Forest Research. 8:290–299.
- Allan W, Ian A. 2003. Landscape-scale species richness of earthworms in the Porongurup range, Western Australia: influence of aspect, soil fertility, and vegetation type. Biology and Fertility of Soils. 39(2):94–102.
- Atsushi O, Tomomi N, Michihiro W. 1993. Relationship between root growth angle of seedlings and vertical distribution of roots in the field in wheat cultivars. Japanese Journal Crop Science. 62(4):565–570.
- Bartelink HH. 1998. A model of dry matter partitioning in trees. Tree Physiology. 18:91–101.
- Byun JG, Lee WK, Nor DK, Kim SH, Choi JK, Lee YJ. 2010. The relationship between tree radial growth and topographic and climatic factors in red pine and oak in central regions of Korea. Journal of Forest Research 99(6): 908–913.
- Cho MG, Chung JM, Jung HR, Moon HS, Kang MY. 2012. Vegetation structure of Picea jezoensis communities in Mt. Deogyu and Mt. Gyebang. Institute of Agriculture & Life Science. 46(6): 33–41. In Korean with English abstract.
- Drexhage M, Gruber F. 1999. Above- and below stump relationships for Picea abies –estimation root system biomass from breast height diameter-. Scanadinavian Journal of Forest Research. 14:328–333.
- Gruber F. 1994. Morphology of coniferous trees: possible effects of soil acidification on the morphology of Norway spruce and Silver fir. Effects of Acid Rain on forest Processes. 8:265–324.
- Harrington G. 1979. Estimation of above-ground biomass of trees and shrubs in a Eucalyptus populnea F. Muell. woodland by regression of mass on trunk diameter and plant height. Australian Journal of Botany. 27(2): 135–143.
- Jo HK, Kim JY, Park HM. 2013. Carbon storage and uptake by evergreen trees for urban landscape for Pinus densiflora and Pinus koraiensis. Korean Journal of Environment and Ecology 27(5): 571–578. In Korean with English abstract.
- Kim IS, Ryu KO, Lee JW. 2012. Climatic factors affecting bud flush timing of Pinus densiflora provenances. Korean Journal of Agricultural and Forest Meteorology 14(4): 229–235. In Korean with English abstract.
- Kim YH, Lee DG. 2000. Growth characteristics and physiological adaptation of Pinus densiflroa seedling in the canopy gap. Journal of Korean Forestry Society 89(3): 452–460. In Korean with English abstract.
- Kong WS, Lim JH. 2008. Distinctive distribution of Vaccinium vitis-idaea and thermal condition. The Korean Geographical Society. 43(4): 495–510. In Korean with English abstract.
- Köstler JN, Bruckner E, Bibelriether E. 1968. Die Wurzeln der Waldbaume. hamburg, Germany: Verlag Paul Parey. 284 pp.
- Kutiel P. 1992. Slope aspect effect on soil and vegetation in a Mediterranean ecosystem. Israel Journal of Plant Sciences. 47(4–6):243–250.
- Lacointe A. 2000. Carbon allocation among tree organs: A review of basic processes and representation in functional-structural tree models. Annals of Forest Science. 57:521–533.
- Le Goff N, Ottorini JM. 2001. Root biomass and biomass increment in a beech (Fagus sylvatica L.) stand in North-East France. Annals of Forest Science. 58:1–13.
- Ledig FT, Borman FH, Wenger KF. 1970. The distribution of dry matter growth between shoot and roots in Loblolly Pine. Botanical Gazette. 131(4):349–359.
- Lee DH. 2004a. Estimation above- and below-ground biomass from diameter of breast height and height for the Pinus densiflora Sieb. et Zucc. Journal of Korean Forest Society. 93(3):242–250. In Korean with English abstract.
- Lee DH. 2004b. Estimation above- and below-ground biomass for Norway Spruce (Picea abies): Estimating root system biomass from breast height diameter. Journal of Korean Forest Society. 90(3):338–345. In Korean with English abstract.
- Lee DH, Yon JH, Woo KS. 2007. Studies on biomass for young Abies koreana Wilson. Journal of Korean Forest Society. 96(2):138–144. In Korean with English abstract.
- Lee DS, Kim DG, Bea KH, Suh HM. 2005. The growth characteristics of the main afforestation species using the change of the annual ring in Uiseong area. Korean Journal of Agricultural and Forest Meteorology 7(4): 274–281. In Korean with English abstract.
- Lee KJ, Kim KD, Kim JS, Park IH. 1985. Distribution of biomass and production of Pinus rigida and Pinus rigida × teaeda plantation in Kwangju district. Journal of Korean Forest Society. 69:28–35. In Korean with English abstract.
- Lee SD, Lee KJ, Choi JW. 2009. Management plan to consider ecological characteristic of Pinus densiflora community in Seoul. Korean Journal of Environment and Ecology 23(3): 258–271. In Korean with English abstract.
- Lee SW. 1981. Studies on forest soils in Korea (II). Journal of Korea Forest Society. 54:25–35. In Korean with English abstract.
- Marler TE, Wilis D. 1996. Chemical or air root pruning containers improve carambola. Logan and mongo seedling root morphology and initial root growth after transplanting. Journal of Environment Horticulture. 14(2):47–49.
- Na SJ, Kim CS, Woo KS, Kim HJ, Lee DH. 2011. Correlation of above- and below-ground biomass between natural and planted stands of Pinus densiflora for. erecta of one age-class in Gangwon Province. Journal of Korean Forest Society. 100(1):42–51. In Korean with English abstract.
- Park IH, Chae KS, Kim KS, Kim MS. 2010. Biomass expansion factors and biomass equations of Pinus densiflora in Suncheon, Jeollanam-do. Proceeding of the Korean Forest Society of spring 2010 in Changwon, Korea. 66–68. In Korean.
- Park IH, Lee SM. 1990. Biomass and net production of Pinus densiflora natural forests of four local forms in Korea. Journal of Korean Forest Society. 79(2):196–204. In Korean with English abstract.
- Qishui Z, John CZ. 1995. Effect of gap size on litter decomposition and microbial activity in a subtropical forest. Ecological Society of America Ecology. 76(7):2196–2204.
- Röhrig E. 1966. Die Wurzelentwicklung der Waldbäume in Abhängigkeit von den Ökologischen Verhältnissen. Forstarchiv 34: 217–227, 237–249.
- Santantonio D. 1990. Modeling growth and production of tree roots. Process modeling of forest growth responses to environmental stress. Timber, Portland, Ore:124–141.
- Seo YO, Lee YJ, Pyo JK, Kim RH, Son YM, Lee KH. 2010. Above- and below-ground biomass and net primary production for Pinus densiflora stands of Cheongyang and Boryeong regions in Chungnam. Journal of Forest Research 99(6): 914–921.
- Scott GD, Hawkins CDB. 2005. Competitive interactions in sub-boreal birch-spruce forests differ on opposing slope aspects. Forest Ecology and Management. 214:1–10.
- Stage AR. 1976. Notes: an expression for the effect of aspect, slope, and habitat type on tree growth. Forest Science 22(4):457–460.
- Whittaker RH, Marks PL. 1975. Methods of assessing terrestrial productivity. In: Lieth H, Whittaker RH, editors. Primary productivity of the biosphere. Berlin Heidelberg: Springer. 55–118.