
ABSTRACT
The size growth patterns of major coniferous trees species Pinus densiflora (Pd), Pinus koraiensis (Pk), and Larix kaempferi (Lk) in South Korea were found and compared using stem analysis. A dominant or co-dominant tree selected from each of 131 sites (39 sites for Pd, 47 sites for Pk, and 45 sites for Lk) was stemmed and analyzed for the study. Total increment (TI), periodic annual increments (PAI), mean annual increments (MAI), and growth percentage of diameter at breast height (DBH), height, and volume were numerically and statistically compared among the three tree species. As the trees age the total DBH growths of Pk and Lk were similar to each other but higher than that of Pd. A noticeable difference of total height growth was shown among the trees and the total height increment was Lk > Pk > Pd. Like the total height growth, the volume difference was noticeably different among the trees and the growth was Lk > Pk > Pd. Given no difference of DBH between Lk and Pk, the volume difference between the two species was due to height. Unlike DBH and height, MAI and PAI of volume did not meet together.. This inferred that the volume growth was still vigorous to reach the rotation age. Growth percentages of DBH, height, and volume tended to decrease with age and no difference was shown among the tree species. The growth percentage of volume was higher than that of DBH and height, while there was no difference between DBH and height.
Introduction
The principle reason for analyzing the past growth of trees is to establish a pattern for predicting future growth (Avery and Burkhart Citation2002). For sustainable forest management the growth patterns for individual trees with information on the size increment are very important factors primarily to log and conserve trees, to estimate cutting cycles and climatic effects, and to prescribe silvicultural treatments (da Silva et al. Citation2002; Brienen and Zuidema Citation2006; Dang et al. Citation2010; Soliz-Gamboa et al. Citation2010). Complete stem analysis is the most accurate method to obtain the growth information including accumulated tree volume growth through a record of the past size growth of individual trees (Husch et al. Citation2003). Also, a relationship between size (diameter, height, or volume) and age of trees can infer age structures and enable assessment of the population dynamics of forests through establishing size class frequency distributions (Harcombe and Marks Citation1978; Lorimer Citation1980; Stewart Citation1986; Bergeron Citation2000; Baker et al. Citation2005; Xing et al. Citation2012).
In South Korea many studies using stem analysis have been carried out on modeling growth parameters of diameter, height, stem volume, site index, or stand density (Ma Citation1997; Park and Chung Citation1999; Lee Citation2002; Lee Citation2003; Choi and Yoo Citation2006; Jeon et al. Citation2007; Lee et al. Citation2009), while, even with limited stem data, there have been only a few studies (Seo et al. Citation2014, Citation2015) conducted on the size growth patterns of diameter, height, and volume for trees.
Meanwhile coniferous tree species, especially Pinus densiflora (Pd), Pinus koraiensis (Pk), and Larix kaempferi (Lk), have been intensively planted as they are grow fast and are used commercially for lumber in South Korea. Of the total forest land in 2015 coniferous species covered about 37% (2,339,022 ha), of which Pd, Pk, and Lk occupied, respectively, about 67% (1,562,843 ha), 7.3% (170,905 ha), and 12% (272,800 ha) (Korea Forest Service Citation2016). As the importance of domestic wood supply and utilization increases, there are greater concerns over how to grow and manage these tree species as well as on how much timber to supply in the future.
Therefore this study aimed to find the size growth patterns of Pd, Pk, and Lk in South Korea. Specifically the study questions were: (1) what kind of size growth patterns exist in the tree species in this study; and (2) whether there is any difference between the growth patterns among the tree species focusing on diameter at breast height (DBH), height, and volume growth.
Materials and methods
Study area
This study was conducted in Pd, Pk, and Lk plantations of the national forest of Gangwon-do, Gyeonggi-do, and Gyeongsangbuk-do in South Korea. During 2012–2015 a total of 131 permanent monitoring plots (20 m × 20 m) in 16 cities and counties were installed: 39 plots for Pd, 47 plots for Pk, and 45 plots for Lk ().
Figure 1. Study area and distribution of stemmed trees by tree species.
The summery statistics of the 131 permanent plots for the tree species were shown in . The mean DBHs were 30.5 cm for Pd, 29.8 cm for Pk, and 27.8 cm for Lk, and the mean heights were 17.0 m for Pd, 17.2 m for Pk, and 22.8 m for Lk. The mean numbers of trees were 843 trees/ha for Pd, 871 trees/ha for Pk, and 782 trees/ha for Lk, the mean basal areas were 36.3 m2/ha for Pd, 36.9 m2/ha for Pk, and 30.6 m2/ha for Lk, and the mean site indices 14.2 m for Pd, 15.3 m for Pk, and 21.4 m for Lk.
Table 1. Summary statistics of 131 permanent plots for the study.
Data collection and analysis
For collecting stem profile data, a dominant or co-dominant tree was felled from each of 131 sites where a plot was installed and the stem sectioned at 2 m intervals beginning at 20 cm with last disc at 1 m or 2 m intervals (Seo et. al. Citation2014, Citation2015). After sanding all discs every fifth ring was marked and cross checked between radii for identifying errors in ring counting. Using a tree ring measuring system with a stereomicroscope, two diameters outside bark (dob) and two diameters inside bark (dib) perpendicular to each other were measured to the nearest 0.001 mm and averaged for all discs. Smalian's formula was used to compute the volume of each section and the volume of a cone was calculated for the portion from the last cut to the tip of the tree.
The summery statistics of the 131 sample trees species in this study are shown in . For each Pd, Pk, and Lk the mean ages were 47, 40, and 38 years old, the mean DBH were 30.4 cm, 29.9 cm, and 27.6 cm, and the mean heights were 17.7 m, 17.3 m, and 22.6 m, respectively. The mean stem volume inside bark and outside bark were 0.6074 m3 and 0.6634 m3 for Pd, 0.6509 m3 and 0.6939 m3 for Pk, and 0.6663 m3 and 0.7498 m3 for Lk.
Table 2. Summary statistics of stemmed trees for this study.
Total increment (TI), mean annual increment (MAI), periodic annual increment (PAI), and growth percentage (GP) of DBH, height, and stem volume were analyzed every 5 years. They were numerically compared among the tree species to calculate the growth patterns of individual tree in terms of age. The analysis of variance (ANOVA) was carried out to statistically verify the growth difference of DBH, height, and volume among the tree species, and Duncan's multiple range test was used to analyze the size growth differences.
Result and discussion
DBH growth pattern
Total DBH growths of Pk and Lk were similar to each other and higher than that of Pd (). The total DBH growth curves of Pk and Lk showed quite similarly increasing pattern each other and were located upper than the total DBH growth curves of Pd at each age. There was a significant difference in total DBH increment among the tree species (p < 0.05). By Duncan's multiple range test (α = 0.05) the difference was not found to be significant between total DBH increments of Pk and Lk, but was definitely significant between Pk (or Lk) and Pd. The DBHs of Pk and Lk grew to 4.7 cm and 6.8 cm at age 10, 22.6 cm and 21.7 cm at age 30, and 32.1 cm and 34.3 cm at age 50, respectively, while DBH of Pd grew to 4.5 cm at age 10, 18.8 cm at age 30, and 28.8 cm at age 50.
Figure 2. Comparing DBH growth patterns by species as the trees age.
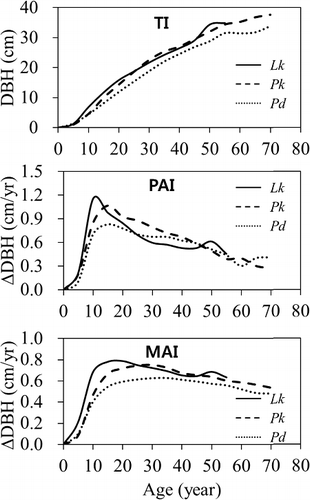
It was shown in all the tree species that the PAI of DBH culminated at early ages and then slowly decreased. The culmination of PAI was Lk > Pk > Pd in order; PAIs of Lk, Pk, and Pd peaked at 1.15 cm at age 10, 1.07 cm at age 15, and 0.83 cm at age 15, respectively. The MAI culmination was smaller than the PAI but had a similar pattern; MAI of Lk, Pk, and Pd peaked at 0.79 cm at age 20, 0.75 cm at age 30 and 0.63 cm at age 35, respectively, where MAI and PAI came together. There was a significant difference among the tree species in both PAI and MAI of DBH (p < 0.0001). By Duncan's multiple range test PAI and MAI of DBH were found not to be significantly different between Pk and Lk, but were significantly different between Pk (or Lk) and Pd.
Height growth pattern
Unlike total DBH growth a noticeable difference was shown among total height growths of Pd, Pk, and Lk (p < 0.0001) and the total increment of height was Lk > Pk > Pd as the trees age (). By Duncan's multiple range test (α = 0.05) the difference was found to be significant among the tree species. The total heights of Pd, Pk, and Lk were 3.3 m, 4.0 m, 7.3 m at age 10, 12.5 m, 14.5 m, 20.2 m at age 30, and 17.0 m, 19.2 m, 26.4 m at age 50, respectively.
Figure 3. Comparing height growth patterns by species as trees age.
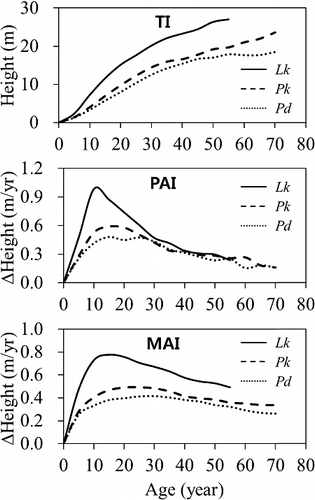
It was shown in all the tree species that the PAI of height culminated at early ages and then slowly decreased, as did DBH. There was a significant difference in PAI of height among the tree species. The culmination of PAI was in the order of Lk > Pk > Pd, getting closer after age 40; PAIs of Lk, Pk, and Pd peaked to 0.98 cm at age 10, 0.59 cm at age 15, and 0.48 cm at age 15, respectively.
The MAI of height pattern was similar to the PAI, but it was more clearly Lk > Pk > Pd. There was a significant difference in MAI of height among the tree species. MAI of Lk, Pk, and Pd peaked at 0.78 cm at age 15, 0.49 cm at age 25, and 0.42 cm at age 30, respectively, where MAI and PAI came together.
Volume growth pattern
DBH and height growth normally presented a concave curve (Liao et al. Citation2003> ; Kwak et al Citation2004; Son et al. Citation2004; Perez Citation2008; Zuidema et al. Citation2011), while volume growth presented a convex curve (Chiba Y and Shinozaki Citation1994; Brienen and Zuidema Citation2006; Jeon et al. Citation2007 ; Castagneri et al. Citation2013). Our study also showed that DBH and height growth curves were slightly concave while volume growth curves were convex ().
Figure 4. Total growth curves of height, DBH, and volume for each tree species.
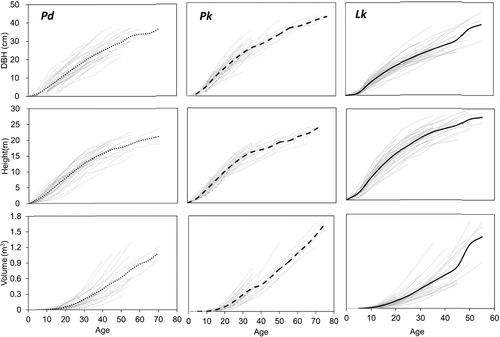
Total volume growth was similar to total height growth and the volume difference was noticeably shown among the trees: Lk > Pk > Pd (). There was a significant difference among the total volumes of the tree species (α = 0.05). The total volumes for Pd, Pk, and Lk grew to 0.0053 m3, 0.0056 m3, 0.0150 m3 at age 10, 0.1930 m3, 0.3144 m3, 0.3707 m3 at age 30, and 0.5803 m3, 0.8010 m3, 1.1402 m3 at age 50, respectively.
Figure 5. Comparing volume growth patterns by species as trees age.
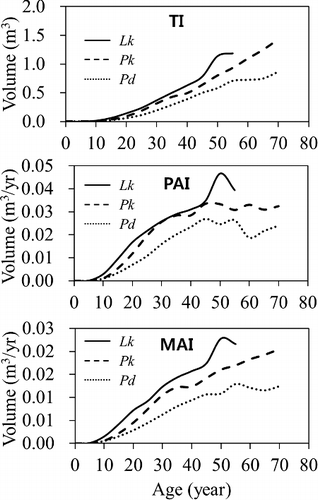
There was a significant difference in PAI and MAI of the volumes among the tree species. Also PAI and MAI of the volumes were differentiated from those of DBH and height in terms of shape and they did not meet together. In the production function its maximum is achieved at the age when MAI is equal to PAI and decides the rotation age. Therefore the volume growth in our study is inferred to be still vigorous to reach the rotation age (). This was consistent with the results from other studies; the PAI of the volume of Japanese cedar (Cryptomeria japonca) increased up to age 150 (Sawata et al. Citation2007) and aging did not reduce PAI of basal area of Norway spruce (Picea abies) over age 150 (Pretzsch et al Citation2002; Castagneri et al. Citation2013).
Growth percentage
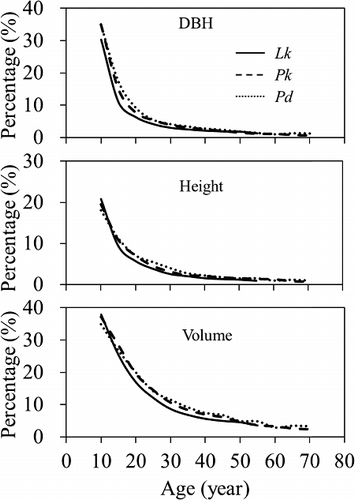
Using Pressler's formula the growth percentages were calculated of DBH, height, and volume for all the tree species. It was found that all the percentages tended to decrease with age in all three tree species () and no significant difference was found between them. While there was no clear difference between the growth percentages of DBH and height, the growth percentage of volume was higher than those of DBH or height in all the tree species. The difference was found to be significant between DBH or height and volume by Duncan's multiple range test (α = 0.05). For all the trees, the growth percentages of DBH and height fell rapidly down to 5% at age 25 and slowly decreased after that, while volume growth percentage was down to 6% at age 40.
Figure 6. Comparing growth percentages of DBH, height, and volume among the tree species.
Conclusion
The study was carried out to find and compare the growth patterns of diameter, height, and stem volume for three major coniferous tree species (Pinus koraiensis, Pinus densiflora, and Larix kaempferi) in South Korea using stem analysis data. TI, PAI, MAI, and growth percentage of DBH, height, and volume were compared numerically and statistically among the three tree species.
With regard to the DBH growth pattern, a noticeable difference in total DBH growth was not shown between Pk and Lk, but was seen between Pk (or Lk) and Pd. The total DBH growth curves of Pk and Lk showed similarly increasing patterns with each other and were located above the total DBH growth curves of Pd at each age. PAI of DBH culminated at the early ages and then slowly decreased. The culmination of MAI was Lk > Pk > Pd in order; MAIs peaked at 0.79 cm at age 20, 0.75 cm at age 30, and 0.63 cm at age 35, respectively, for Lk, Pk, and Pd. The PAI difference was not significant between Pk and Lk, but was significantly different between Pk (or Lk) and Pd.
A noticeable difference of total height growth was shown among Pd, Pk, and Lk and the total height increment was Lk > Pk > Pd as the trees age. The total heights of Pd, Pk, and Lk grew to 3.3 cm, 4.0 cm, 7.3 cm at age 10, 12.5 cm, 14.5 cm, 20.2 cm at age 30, and 17.0 cm, 19.2 cm, 26.4 cm at age 50, respectively. PAI of height culminated at the early ages and then slowly decreased in the same way as DBH. There was a significant difference in MAI of height among the tree species and the MAI peaked at 0.78 cm at age 15, 0.49 cm at age 25, and 0.42 cm at age 30, respectively, for Lk, Pk, and Pd.
Total volume growth was similar to total height growth and the volume difference was noticeably different among the trees; Lk > Pk > Pd. Given no difference of DBH between Lk and Pk the volume difference was due to the difference of height between the two trees. The difference was significant among the volumes of the trees. The total volume grew to 0.0053 m3, 0.0056 m3, 0.0150 m3 at age 10, 0.1930 m3, 0.3144 m3, 0.3707 m3 at age 30, and 0.5803 m3, 0.8010 m3, 1.1402 m3 3 at age 50, respectively. MAI and PAI of volume were differentiated from those of DBH and height in that MAI and PAI of volume did not meet together. This inferred that the volume growth was still vigorous to reach the rotation age.
Growth percentages of DBH, height, and volume tended to decrease with age and no difference was shown between the tree species. While there was no clear difference between the growth percentages of DBH and height, the growth percentage of volume was higher than those of DBH and height in all the trees. For all the trees the growth percentages of DBH and height fell rapidly down to 5% at age 25 and volume growth percentage down to 6% at age 40.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Avery TE, Burkhart HE. 2002. Forest measurements. 5th ed. New York (NY): McGraw-Hill.
- Baker PJ, Bunyavejchewin S, Oliver CD, Ashton PS. 2005. Disturbance history and historical stand dynamics of a seasonal tropical forest in western Thailand. Ecol Monogr 75: 317–343.
- Bergeron Y. 2000. Species and stand dynamics in the mixed woods of Quebec's southern boreal forest. Ecology 81: 1500–1516.
- Brienen RJ, Zuidema PA. 2006. Lifetime growth patterns and ages of Bolivian rain forest trees obtained by tree ring analysis. J Ecol. 94:481–493.
- Castagneri D, Storaunet KO, Rolstad J. 2013. Age and growth patterns of old Norway spruce trees in Trillemarka forest, Norway. Scand J For Res. 28:232–240.
- Chiba Y, Shinozaki K. 1994. A simple mathematical model of growth pattern in tree stems. Ann Bot. 73:91–98.
- Choi JK, Yoo BO. 2006. Diameter growth characteristics of Quercus mongolica and Quercus variabilis in natural deciduous forests. J Korean For Soc. 95:131–138. ( in Korean with English abstract).
- da Silva RP, dos Santos J, Tribuzy ES, Chambers JQ, Nakamura S, Higuchi N. 2002. Diameter increment and growth patterns for individual tree growing in Central Amazon, Brazil. For Ecol Manage. 166:295–301.
- Dang, H, Zhang Y, Zhang K, Jiang M, Zhang Q. 2010. Age structure and regeneration of subalpine fir (Abies fargesii) forests across an alititudial range in the Quinling Mountains, China. For Ecol Manage 259: 547–554.
- Harcombe PA, Marks PL. 1978. Tree diameter distributions and replacement processess in southeast Texas forests. For Sci 24: 153–166.
- Husch B, Beers TW, Kershaw JA Jr. 2003. Forest mensuration. 4th ed. Hoboken (NJ): Wiley.
- Jeon BH, Lee SH, Lee YJ, Kim H, Kang HM. 2007. Estimation of site index and stem volume equations for Larix leptolepis stand in Jinan, Chonbuk. J Korean For Soc. 96:40–47. ( in Korean with English abstract).
- Korea Forest Research Institute. 2012. Stem volume, biomass, and yield table. Seoul: Korea Forest Research Institute. ( in Korean).
- Korea Forest Service. 2016. Statistical yearbook of forestry. Daejeon: Korea Forest Service.
- Kwak DA, Lee WK, Kim SW, Seo JH. 2004. Regional differences in height and taper form of Pinus densiflora in Korea. Kor J For Meas. 7:49–60. ( in Korean with English abstract).
- Lee SH. 2003. Development of site index equation and curves for Cryptomeria japonica. J Korean For Soc. 92:152–159.
- Lee ST, Bae SW, Jang SC, Hwang JH, Chung JM, Kim HS. 2009. A study on the relationship between radial growth and climate factors by regions in Korean pine (Pinus koraiensis). J Korean For Soc. 98:733–739. ( in Korean with English abstract).
- Lee YJ. 2002. Dominant height growth and site index curves for pitch-loblolly hybrid pine (Pinus rigida × P. taeda) plantations. J Korean For Soc. 91:661–666.
- Liao CY, Podrazsky VV, Liu GB. 2003. Diameter and height growth analysis for individual white pine trees in the area of Kostelec nad Cernymi lesy. J For Sci. 49:544–551.
- Lorimer, CG. 1980. Age structure and disturbance history of a southern Appalachian virgin forest. Ecology 61: 1169–1184.
- Ma SK. 1997. Productivity and density control of stands of Japanese larch. J Korean For Soc. 34:21–30. ( in Korean with English abstract).
- Park MS, Chung YG. 1999. Estimation of diameter and height growth in Pinus thunbergii stands using linear and nonlinear growth functions. J Korean For Soc. 88:47–54. ( in Korean with English abstract).
- Perez D. 2008. Growth and volume equations developed from stem analysis for Tetona grandis in Costa Rica. J Trop For Sci. 20:66–75.
- Pretzsch H, Biber J, Dursky J. 2002. The single tree-based stand simulator SILVA: construction, application and evaluation. For Eco Mgt. 162:3–21.
- Sawata S, Nishizono T, Awaya Y, Nobori Y. 2007. Analysis of stem growth pattern in Japanese cedar (Cryptomeria japonica) trees in a natural forest in Akita, northeastern Japan. J Jap Forest Soc. 89:200–207. ( in Japanese with English abstract).
- Seo YW, Balekoglu S, Choi JK. 2014. Growth pattern analysis by stem analysis of Korean white pine (Pinus koraiensis) in the central northern region of Korea. For Sci Tech. 10:220–226.
- Seo YW, Lee DS, Choi JK. 2015. Growth analysis of Korean red pine (Pinus densiflora) by stem analysis in the eastern region of Korea. J For Env Sci. 31:47–54.
- Soliz-Gamboa CC, Rozendaal DM, Ceccantini G, Angyalossy V, van der Borg K, Zuidema PA. 2011. Evaluating the annual nature of juvenile rings in Bolivian tropical rain forest trees. Trees 25: 17–27.
- Son YM, Lee KH, Kweon SD. 2004. Estimation of stem taper equation and growth pattern in Pinus koraiensis by two districts. Kor J For Meas. 7:9–14. ( in Korean with English abstract).
- Stewart GH. 1986. Population dynamics of a montane conifer forest, western Cascade Range, Oregon, USA. Ecology 67: 534–544.
- Xing P, Zhang Q-B, Baker P. 2012. Age and radial growth pattern of four tree species in a subtropical forest of China. Trees. 26:283–290.
- Zuidema PA, Vlam M, Chien PD. 2011. Ages and long-term growth patterns of four threatened Vietnamese tree species. Trees. 25:29–38.