
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Woody plant species are important components of agro-ecosystems that determine their productivity and biodiversity conservation in tropical mountain landscapes. This study investigates woody plant species composition and diversity through simple random sampling of sixty (10 × 10 m) plots in crop fields, hedgerows, and homegardens at Uluguru Mountains, Morogoro, Tanzania. In total, 85 woody plant species, representing 82 genera and 40 families were recorded. The most species-rich families were Proteaceae (with 323 individuals), Lauraceae (205), and Meliaceae (185). Among the woody plant species, Grevillea robusta (Proteaceae) showed the highest importance value index of 41% across different agro-ecosystem types. Shannon (1.87), Simpson (0.82), and evenness (0.91) diversity indices were highest in hedgerows compared to other agro-ecosystems. Hedgerows recorded the maximum richness, followed by homegardens and lastly crop fields. Present agro-ecosystems should be promoted to enhance the restoration and conservation of agro-biodiversity in mountain landscapes.
Introduction
Mountains cover 24% of the world’s land area, with approximately 12% of the world’s population in mountainous regions (Schild Citation2008). They harbor a high diversity of plant species and are rich in endemic species (Noroozi et al. Citation2019). Mountain ecosystems provide numerous goods and services to a significant proportion of the global population (Mazzocchi and Sali Citation2022). Millions of people in Africa depend on goods and services from mountains, which support livelihoods at local, national, and regional levels. However, growing population in African countries has increased demands for ecosystem services such as food, and hence pressing needs for sustainable agricultural practices (Rockström et al. Citation2017). This can be achieved through proper management of agro-ecosystems by upholding management practices that conserve biodiversity and provide ecosystem services to meet the demands of the ever-growing population (Malinga Citation2016).
Several dynamics evolving in agricultural activities have resulted in the shift from monoculture systems to the integration of trees and crops on farmlands (Awazi and Tchamba Citation2019). This transformative trend emerged due to the growing recognition of the multifaceted benefits from integration of trees and crops within the same agricultural space. Pressing challenges such as soil degradation, climate change, and the decline of natural resources have influenced the emergency of new practices in agroforestry and other integrated farming systems to face the on-going challenges in agriculture (Samsudin et al. Citation2020). Retaining trees on farmlands contributes to biodiversity conservation and the provision of ecosystem services such as control of soil erosion and improvement of carbon sequestration potential (Pandey et al. Citation2017) as well as the supply of food and income to local communities (Lasco et al. Citation2014).
Agro-ecosystems, as an integral part of the global landscape play a crucial role in supporting the human population and fostering ecological balance (Lescourret et al. Citation2015). The composition and diversity of woody plant species within agro-ecosystems emerge as critical determinants of ecosystem health, resilience, and productivity. The complex interaction between agriculture and ecology enhances the need for a comprehensive understanding of woody plant species inhabiting agro-ecosystems (Ahmed et al. Citation2015). Agro-ecosystems also play a great role in conserving diverse woody plant species for biodiversity conservation as well as for livelihood improvements (Mengistu and Asfaw Citation2016). The choice of trees and shrub-based agro-ecosystems has positive implications for biodiversity conservation and meeting the demands of local communities, including those of fuelwood, construction materials, and non-timber forest products (Duguma and Hager Citation2010).
Although several studies have pointed out the contribution of agro-ecosystems in providing economic and environmental benefits among rural communities in Africa (Mazzocco Citation2007; Diao et al. Citation2010; Branca et al. Citation2021) few have focused on the composition and diversity of woody plant species from mountain agro-ecosystems (Neba Citation2009; Mengistu and Asfaw Citation2016). Lack of detailed information on woody plant species composition and diversity in agro-ecosystems makes it difficult to appreciate the importance of woody plants for agro-biodiversity conservation as well as rural livelihoods. Therefore, the objective of this study is to systematically assess the variation of species composition and diversity in woody plant species among agro-ecosystem types in the Uluguru Mountains, Morogoro, Tanzania. This will enhance the conservation and restoration of agro-ecosystems for the wellbeing of local communities as well as agro-biodiversity conservation in mountain landscapes.
Materials and methods
Study villages
This study was conducted in two villages (Choma and Turo) in the Uluguru Mountains, located in the Morogoro region, eastern part of Tanzania. The two villages are situated at the latitudes 7°02′–7°16′S and longitudes 38°0′–38°12′E (). Uluguru Mountains are found about 200 km west of Dar es Salaam, Tanzania and are among the major biodiversity hotspots of the world because the mountains house a high number of endemic species (Myers et al. Citation2000). The climate of the area is oceanic, characterized by mild and oceanic conditions with no pronounced dry season, but short dry periods occur. Rainfall peaks in April and November, indicating that these months are likely to have higher precipitation levels (Munishi and Shear Citation2005). This study was restricted to the northern part of Uluguru Mountains; where annual rainfall varies with altitude from 900 mm at approximately 500 m above sea level (m.a.s.l) to 2300 mm at 1500 m.a.s.l. The mean annual air temperature ranges from 25 °C around 550 m.a.s.l to 19 °C at 1500 m.a.s.l (Kimaro et al. Citation2008). Vegetation type of the mountains is divided into moist forest on the eastern part and savannah woodland at lower altitudes on western slopes below 1000 m (Burgess et al. Citation2002).
Figure 1. Location map showing study villages in Uluguru Mountains, Morogoro, Tanzania.
The study villages are located in Mlimani ward, and most of the people living in these villages are Luguru (Mhanga and Massawe Citation2021). Choma village is located on the hill side of the Uluguru Mountains and receives high rainfall, which produces streams and small rivers supplying water to farmers. From the studied area, the village recorded an average altitude of 1220 m.a.s.l., which was higher than that for Turo village. The people living in Choma are crop farmers practicing livestock keeping as well as managing small business activities. People practice sustainable agricultural activities that depend on water and are sufficient in the Choma village (Harrison and Mdee Citation2017). Turo village is located at a lower altitude of 1192 m.a.s.l. compared to Choma village. Villagers practice crop production and other non-farm activities such as livestock keeping, food vending, village community banks, and entrepreneurship activities like making home-made soaps to sustain livelihood at Turo (Mhanga & Massawe Citation2021).
Vegetation sampling
A reconnaissance survey was carried out to identify the study villages, verify the agro-ecosystems, and establish an adequate sampling method prior to actual vegetation sampling. Two villages with crop fields, hedgerows, and homegarden agro-ecosystems () were selected and surveyed to assess the composition and diversity of woody plant species. The choice of selected agro-ecosystem types was based on vegetation arrangements of trees, shrubs, and lianas associated with other crops on farmlands. A total of sixty (60) 10 × 10 m sized sample plots were selected using simple random sampling and enumerated in Choma and Turo villages. The plot size of 10 × 10 m was appropriate for this study since a sufficient number of woody plants were included in that plot size to make a meaningful assessment. The sample plots used were independent sampling units and not subplots. Distance between plots was estimated at a minimum interval of 150 m to avoid pseudoreplication. To identify woody plant species within the plots, trees, shrubs, and lianas were independently enumerated. In each plot, all trees and shrubs measuring ≥2 m in height were recorded accordingly (Endale et al. Citation2017). A canopy cover for each individual plant species in each plot was visually estimated. The names of the sampled woody plant species were recorded with the help of the field guide and the botanist. In cases where woody plants were not identified in the field, voucher specimens were collected, processed, and submitted for identification at the Herbarium of the University of Dar es Salaam.
Table 1. Description of the agro-ecosystems in Uluguru Mountains, Morogoro, Tanzania.
Data analysis
Floristic composition and diversity
The comparison of the floristic composition of different agro-ecosystems was done using the Sorensen coefficient of similarity (Ss) given as Ss = 2a/(2a + b + c), where Ss = Sorensen similarity coefficient, a = number of species common in both samples, b = number of species in sample 1, c = number of species in sample 2. To determine the expected number of species in this study, a species accumulation curve representing different agro-ecosystem types was plotted in R (Hsieh et al. Citation2016). Relative canopy cover, relative density, and relative frequency were calculated and used to determine the species important value indices (IVIs). The relative density, relative canopy cover, and relative frequency were calculated using (Mueller-Dombois and Ellenberg Citation1974):
Plant species diversity across agro-ecosystems was computed using Shannon diversity (H), Eveness (J), and Simpson diversity (D) indices given as follows:
Shannon diversity index:
where H = Shannon diversity index, n = number of species in a community and pi = proportion of individuals of the ith species expressed as a proportion of total abundance in the sample.
Evenness measures homogeneous distribution of species in sample plot and was calculated as:
where J = Evenness, H = Shannon-Wiener diversity, Hmax = ln(S), S = the total number of species in the sample.
Simpson’s diversity Index which assesses the most abundant species in the community and is more sensitive to changes in a community (Magurran Citation1988) was calculated as:
where D = Simpson’s index, n = the total number of individuals of a particular species and N = the total number of individuals of all species.
To test if data on richness and diversity were normally distributed, the Shapiro-Wilk test was used, and the data were considered significant at p ≤ 0.05. Levene’s test was applied to compute the equality of variances. In cases where data on richness and diversity were found not to follow a normal distribution, the data were transformed into log values (Eyasu et al. Citation2020). A t-test was used to test for significance between the means of diversity for the two villages. Comparisons of evenness, Shannon diversity, and Simpson diversity indices between the agro-ecosystems of the two villages were made using one-way analysis of variance (ANOVA). Significant means of diversity indices were separated using Tukey’s HSD test at the 5% level of significance in Statistical Package for Social Science (SPSS) version 25.
Results
Woody plant species composition
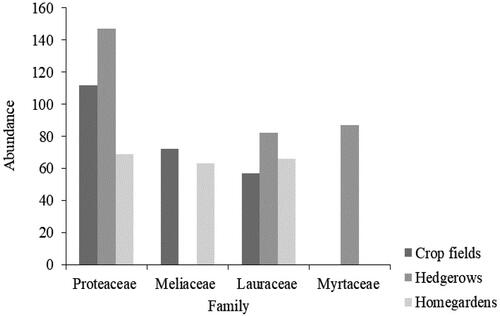
A total of 85 woody plant species belonging to 40 families and 82 genera were recorded from the two villages (). Out of this, hedgerows had more woody plant species (68 from 34 families) than homegardens (34 from 19 families) and crop fields (37 from 22 families). The three dominant families were Proteaceae, Lauraceae, and Meliaceae with abundances of 323, 205, and 185 individuals, respectively. The identified woody plant species were composed of 55 tree species, 28 shrub species, and 2 lianas. Overall, the average number of species in a plot in the three agro-ecosystems was 5.93 (±2.78 SD) ().
Table 2. Characteristics of the study villages in Uluguru Mountains, Morogoro, Tanzania.
The three most abundant families in crop fields were Proteaceae (112), Meliaceae (72) and Lauraceae (57) while for hedgerows were Proteaceae (147), Myrtaceae (87) and Lauraceae (82) and for homegardens were Proteaceae (69), Lauraceae (66) and Meliaceae (63) ().
Figure 2. Abundance of the top three woody plant families in different agro-ecosystems of Uluguru Mountains, Morogoro, Tanzania.
Importance value index
The most important woody plant species with higher IVIs in the agro-ecosystems were Grevillea robusta, Persea americana, and Cedrela odorata. These woody plant species contributed 63.1, 38.9, and 20.5% of the total population of woody plant species in hedgerows, homegardens, and crop fields ().
Table 3. The top fifteen woody plant species with the highest IVIs (%) in Uluguru Mountains, Morogoro, Tanzania.
The five most important woody plant species with the highest IVI’s in each agro-ecosystem were recorded as Grevillea robusta (86%), Persea americana (34%), Acrocarpus fraxinifolius (24%), Prunus persica (21%) and Khaya anthotheca (20%) in crop fields. Top ranking in hedgerows were Grevillea robusta (59%), Persea americana (35%), Bridelia micrantha (23%), Cedrela odorata (15%), and Eucalyptus grandis (11%) and the top-ranking species in homegardens were Grevillea robusta (53%), Persea americana (48%), Cedrela odorata (31%), Artocarpus heterophyllus (26%) and Prunus persica (21%), respectively.
Woody plant species similarity between agro-ecosystems
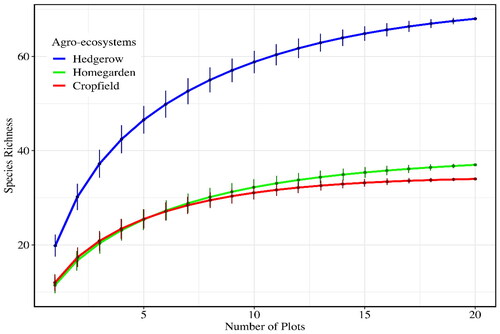
The highest similarity (59%) in woody plant species composition was recorded between crop fields and homegardens. This was followed up by a comparison between home gardens and hedgerows (54%), and the lowest was recorded between hedgerows and crop fields (52%). A comparison of woody plant species among all three agro-ecosystems indicates that there is an overall high similarity (60%) in woody plant species across the three systems. Species accumulation curves showed that hedgerows had higher species richness than crop fields and homegardens. Sufficient numbers of samples were considered during the determination of woody plant species diversity in agro-ecosystems ().
Figure 3. Species accumulation curve comparing species richness among agro-ecosystems in the Uluguru Mountains, Morogoro, Tanzania.
Woody plant species diversity
The average Shannon diversity of woody plant species in a plot was crop fields 1.09 (±0.48 SD), hedgerows 1.87 (±0.33 SD) and homegardens 1.40 (±0.38 SD). Average Simpson diversity recorded 0.58 (±0.20 SD) for crop fields, 0.82 (±0.07 SD) for hedgerows and 0.68 (±0.14 SD) for homegardens. Recorded values for evenness were 0.80 (±0.22 SD), 0.91 (±0.09 SD), and 0.84 (±0.10 SD) for crop fields, hedgerows and homegardens, respectively. Overall, the average values for Shannon, Simpson, and evenness diversity indices for woody plant species in a plot were 1.46 (±0.51 SD), 0.69 (±0.17 SD) and 0.85 (±0.15 SD), respectively. The diversity indices showed high values in the hedgerows compared to crop fields or homegardens. There was a significant difference (p < 0.001) between mean values of the diversity indices of woody plant species within plots in each village ().
Table 4. Means (± SD) of different diversity indices recorded in the two villages of Uluguru Mountains, Morogoro, Tanzania.
An independent t-test indicated that Choma village had a higher diversity of woody plant species than Turo village. The two villages showed a significant difference in the Simpson diversity index (p = 0.04). However, there was no significant difference in richness, evenness, or Simpson diversity between the agro-ecosystems of the two villages ().
Table 5. Means (±SD) of diversity indices of the agro-ecosystems of the two villages in Uluguru Mountains, Morogoro, Tanzania.
Discussion
Woody plant species composition
The current study revealed that indeed mountain agro-ecosystems host diverse woody plant species ranging from lianas, shrubs, and trees. These findings enhance the significance of considering various agro-ecosystems, specifically hedgerows, homegardens, and crop fields, as distinctive units for assessing plant diversity and the restoration of agrobiodiversity (Isbell et al. Citation2017). In particular, the highest number of woody plant species was recorded in hedgerows compared to home gardens and crop fields. This suggests that hedgerows serve as a crucial reservoir of biodiversity, potentially providing ecological services and supporting an immense assemblage of flora. While homegardens and crop fields exhibited low species richness, the possible reason for the variation in species composition among these agro-ecosystems could be attributed to land management practices and disturbance regimes (Hall et al. Citation2020). A species found in hedgerows only or rare to other agro-ecosystems included Grevillea robusta (timber tree), Persea americana (fruit tree), and Eucalyptus grandis (timber and firewood tree).
Proteaceae emerged as the most abundant family, indicating its ecological significance and potential adaptability to the local environmental conditions in mountain landscapes. Members of this plant family are characterized by the provision of biodiversity support such as nectar and pollen. Ecosystem resilience is also another attribute that demonstrates that woody plant species from this family can adapt to and withstand certain environmental stressors (Oyanoghafo Citation2022). The prominence of Grevillea robusta, Paersea americana, and Cedrela odorata in the three agro-ecosystems indicates that these species play multiple significant roles in the local communities. These key species fulfill ecological functions such as soil stabilization (Grevillea robusta), provisioning of food sources (Persea americana), and valuable timber (Cedrela odorata). They also suggest potential opportunities for agroforestry and sustainable land management practices. For example, Grevillea robusta supports nitrogen fixation, which enhances soil fertility in agroforestry systems (Muthuri et al. Citation2023). Similarly, these findings are partly in agreement with Samuel Citation2020, who reported Grevillea robusta and Persea americana to be among the important tree species in farmers home gardens in Ethiopia.
The higher similarity of woody plant species between crop fields and homegardens could be attributed to common land management practices, such as the introduction of specific plant species for different purposes. The lower similarity of woody plant species between hedgerows and other agro-ecosystems could be due to distinctive management practices that contribute to more specialized species associations. Overall, high similarity (60%) in woody plant species across all three agro-ecosystems is due to ecological connectivity and shared functions among the three agro-ecosystems (Santos et al. Citation2019). A few of woody plant species common in all the three agro-ecosystems included Grevillea robusta (timber tree), Persea americana (fruit tree), and Mangifera indica (fruit tree). In this study, it has been depicted that there are differences in species richness among the different agro-ecosystems. Higher species richness was observed in hedgerows, followed by homegarden, and lastly crop fields. These findings are in line with (Duguma and Hager Citation2010) who reported that there was a significant difference in species richness among land use practices in central highlands of Ethiopia.
Woody plant species diversity
It was also interesting to note that diversity values were higher in hedgerows than in home gardens and crop fields. Higher Shannon diversity values in hedgerows suggest that hedgerows have a greater variety of woody plant species or a more even distribution of woody plant species. Also, a likely explanation for this is that the absence of intensive management practices such as thinning and pruning in hedgerows influences the diversity of woody plant species in hedgerows more than in other agro-ecosystems. High values of Simpson diversity in hedgerows were influenced by the large number of dominant species in hedgerows compared to other agro-ecosystems. Some of these dominant species in hedgerows were Grevillea robusta, Persea americana, Khaya anthotheca, and Prunus persica. Shannon and Simpson diversity values recorded in homegardens in this study are higher than those found in homegardens in a study in Tigray, northern Ethiopia (H = 1.31, D = 1.64), perhaps due to differences in methodology (Eyasu et al. Citation2020).
The lowest diversity values were recorded in crop fields in comparison to other agro-ecosystems. This might be due to management practices such as intensive pruning and thinning of woody plant species to reduce competition with crops. This can also be due to the rareness of some woody plant species or the nonexistence of woody plant species that were found in hedgerows and home gardens. Farmers’ preferences for woody plant species might have also indirectly influenced lower diversity values in crop fields. These findings are unlike those from a previous study (Amare Citation2018), which recorded higher diversity indices in homegardens in the south-central highlands of Ethiopia, partly due to differences in methodology.
Choma village exhibited a great diversity of woody plant species in comparison to Turo village. This could be due to differences in elevation gradients and management practices by farmers. Choma village is located at a high altitude of 1220 m.a.s.l. and Turo at an altitude of 1191 m.a.s.l. Variation in elevation might have influenced the richness of woody plant species in the two villages (Thakur and Yadava Citation2021). Another reason influencing the variation in the number of species between the two villages is the extent of management practices (Angessa et al. Citation2020) for woody plant species across agro-ecosystems. Simpson being significant at (p < 0.04) implies that the observed differences are unlikely to have occurred by chance, but rather are attributed to factors such as the dominance of woody plant species.
This study highlights that mountain agro-ecosystems exhibit diverse range of woody plant species, including trees, shrubs, and lianas, whose diversity emphasize the ecological complexity of these systems. Hedgerows appear to host higher numbers of woody plant species compared to other agro-ecosystems suggesting that hedgerows play a crucial role as reservoirs of biodiversity. Into such accounts, the potential role of woody plant species on conservation of biodiversity should not be restricted only to natural forests, but it should consider expanding to human managed farmlands or agro-ecosystems. This is because they play a great role in developing biodiversity islands which host a large number of woody plant species for conservation of flora and fauna (Guo Citation2000). Improved agro-ecosystems will indeed help to reduce pressure on natural forests brought about by increasing human population and their demands.
Disclosure statement
Authors declare that there is no conflict of interest to declare.
References
- Ahmed S, Stepp JR, Xue D. 2015. Cultivating botanicals for sensory quality: from good agricultural practices (GAPs) to taste discernment by smallholder tea farmers. Botanicals: methods for quality and authenticity. Boca Raton: CRC Press, Taylor & Francis Group, LLC.
- Amare A. 2018. Woody plant species diversity in agroforestry system of homegarden and crop lands in Gedebano Gutazer Welenne Wereda, Gurage Zone, SNNPR, Ethiopia [MSc Thesis]. School of Graduate Studies of Arba Minch University.
- Angessa AT, Lemma B, Yeshitela K, Fischer J, May F, Shumi G. 2020. Woody plant diversity, composition and structure in relation to environmental variables and land-cover types in Lake Wanchi watershed, central highlands of Ethiopia. Afr J Ecol. 58(4):627–638. doi: 10.1111/aje.12731.
- Awazi NP, Tchamba NM. 2019. Enhancing agricultural sustainability and productivity under changing climate conditions through improved agroforestry practices in smallholder farming systems in sub-Saharan Africa. Afr J Agric Res. 14(7):379–388.
- Branca G, Arslan A, Paolantonio A, Grewer U, Cattaneo A, Cavatassi R, Lipper L, Hillier J, Vetter S. 2021. Assessing the economic and mitigation benefits of climate-smart agriculture and its implications for political economy: a case study in Southern Africa. J Cleaner Prod. 285:125161. doi: 10.1016/j.jclepro.2020.125161.
- Burgess N, Doggart N, Lovett JC. 2002. The Uluguru mountains of eastern Tanzania: the effect of forest loss on biodiversity. Oryx. 36(2):140–152. doi: 10.1017/S0030605302000212.
- Diao X, Hazell P, Thurlow J. 2010. The role of agriculture in African development. World Dev. 38(10):1375–1383. doi: 10.1016/j.worlddev.2009.06.011.
- Duguma LA, Hager H. 2010. Woody plants diversity and possession, and their future prospects in small-scale tree and shrub growing in agricultural landscapes in central highlands of Ethiopia. Small-Scale For. 9(2):153–174. doi: 10.1007/s11842-009-9108-0.
- Endale Y, Derero A, Argaw M, Muthuri C. 2017. Farmland tree species diversity and spatial distribution pattern in semi-arid East Shewa, Ethiopia. Forests Trees Livelihoods. 26(3):199–214. doi: 10.1080/14728028.2016.1266971.
- Eyasu G, Tolera M, Negash M. 2020. Woody species composition, structure, and diversity of homegarden agroforestry systems in southern Tigray, Northern Ethiopia. Heliyon. 6(12):e05500. doi: 10.1016/j.heliyon.2020.e05500.
- Guo Q. 2000. Climate change and biodiversity conservation in great plains agroecosystems. Global Environ Change. 10(4):289–298. doi: 10.1016/S0959-3780(00)00035-2.
- Hall RM, Penke N, Kriechbaum M, Kratschmer S, Jung V, Chollet S, Guernion M, Nicolai A, Burel F, Fertil A, et al. 2020. Vegetation management intensity and landscape diversity alter plant species richness, functional traits and community composition across European vineyards. Agric Syst. 177:102706. doi: 10.1016/j.agsy.2019.102706.
- Harrison E, Mdee A. 2017. Successful small-scale irrigation or environmental destruction? The political ecology of competing claims on water in the Uluguru Mountains, Tanzania. J Political Ecol. 24(1):406–424. doi: 10.2458/v24i1.20881.
- Hsieh TC, Ma KH, Chao A. 2016. INEXT: an R package for rarefaction and extrapolation of species diversity (H ill numbers). Methods Ecol Evol. 7(12):1451–1456. doi: 10.1111/2041-210X.12613.
- Isbell F, Gonzalez A, Loreau M, Cowles J, Díaz S, Hector A, Mace GM, Wardle DA, O’Connor MI, Duffy JE, et al. 2017. Linking the influence and dependence of people on biodiversity across scales. Nature. 546(7656):65–72. doi: 10.1038/nature22899.
- Kimaro DN, Poesen J, Msanya BM, Deckers JA. 2008. Magnitude of soil erosion on the northern slope of the Uluguru Mountains, Tanzania: interrill and rill erosion. CATENA. 75(1):38–44. doi: 10.1016/j.catena.2008.04.007.
- Lasco RD, Delfino RJP, Catacutan DC, Simelton ES, Wilson DM. 2014. Climate risk adaptation by smallholder farmers: the roles of trees and agroforestry. Curr Opin Environ Sustain. 6:83–88. doi: 10.1016/j.cosust.2013.11.013.
- Lescourret F, Magda D, Richard G, Adam-Blondon A-F, Bardy M, Baudry J, Doussan I, Dumont B, Lefèvre F, Litrico I, et al. 2015. A social–ecological approach to managing multiple agro-ecosystem services. Curr Opin Environ Sustain. 14:68–75. doi: 10.1016/j.cosust.2015.04.001.
- Magurran A. 1988. Ecological diversity and its measurements. New Jersey: Princeton University Press, United States of America.
- Malinga R. 2016. Ecosystem services in agricultural landscapes: A study on farming and farmers in South Africa and Sweden [Doctoral dissertation]. Stockholm University. http://urn.kb.se/resolve?urn=urn:nbn:se:su:diva-133485.
- Mazzocchi C, Sali G. 2022. Tourists’ perception of ecosystem services provided by mountain agriculture. Sustainability. 14(19):12171. doi: 10.3390/su141912171.
- Mazzocco M. 2007. Household intertemporal behaviour: a collective characterization and a test of commitment. The Review of Economic Studies. 74(3):857–895. doi: 10.1111/j.1467-937X.2007.00447.x.
- Mengistu B, Asfaw Z. 2016. Woody species diversity and structure of agroforestry and adjacent land uses in Dallo Mena District, South-East Ethiopia. Nat. Resour. 07(10):515–534. doi: 10.4236/nr.2016.710044.
- Mhanga M, Massawe FA. 2021. The role of conservation agriculture in bridging gender gaps in Tanzania: the case of Sustainable Agriculture Tanzania. 2nd Conference. p. 353–361.
- Mueller-Dombois D, Ellenberg H. 1974. Aims and methods of vegetation ecology. New York: John Willey and Sons Inc.
- Munishi PKT, Shear TH. 2005. Rainfall interception and partitioning in afromontane rain forests of the Eastern Arc Mountains, Tanzania: implications for water conservation. J Trop for Sci. 17(3):355–365.
- Muthuri CW, Kuyah S, Njenga M, Kuria A, Öborn I, van Noordwijk M. 2023. Agroforestry’s contribution to livelihoods and carbon sequestration in East Africa: a systematic review. Trees, Forests People. 14:100432. doi: 10.1016/j.tfp.2023.100432.
- Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature. 403(6772):853–858. doi: 10.1038/35002501.
- Neba NE. 2009. Management of woody plants in indigenous land use systems of the Sahel: example of north Cameroon. Int NGO J. 4(11):480–490.
- Noroozi J, Naqinezhad A, Talebi A, Doostmohammadi M, Plutzar C, Rumpf SB, … & Schneeweiss GM. 2019. Hotspots of vascular plant endemism in aglobal biodiversity hotspot in Southwest Asia suffer from significant conservation gaps. Biol. Cons. 237:299-307. doi: 10.1016/j.biocon.2019.07.005.
- Oyanoghafo OO. 2022. Vulnerability and adaptive capacity of Proteaceae family to drought.
- Pandey R, Aretano R, Gupta AK, Meena D, Kumar B, Alatalo JM. 2017. Agroecology as a climate change adaptation strategy for smallholders of Tehri-Garhwal in the Indian Himalayan Region. Small-Scale For. 16(1):53–63. doi: 10.1007/s11842-016-9342-1.
- Rockström J, Williams J, Daily G, Noble A, Matthews N, Gordon L, Wetterstrand H, DeClerck F, Shah M, Steduto P, et al. 2017. Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio. 46(1):4–17. doi: 10.1007/s13280-016-0793-6.
- Samsudin YB, Puspitaloka D, Rahman SA, Chandran A, Baral H. 2020. Community-based peat swamp restoration through agroforestry in Indonesia. Agrofor Degrad Landsc. 1:349–365.
- Samuel T. 2020. Diversity and composition of useful plant species along altitudinal gradient in selected homegardens of Kembata Tembaro zone, Snnpr of Ethiopia [Doctoral dissertation]. Hawassa University.
- Santos M, Cagnolo L, Roslin T, Marrero HJ, Vázquez DP. 2019. Landscape connectivity explains interaction network patterns at multiple scales. Wiley Online Library.
- Schild A. 2008. ICIMOD’s position on climate change and mountain systems. Mt Res Dev. 28(3/4):328–331. doi: 10.1659/mrd.mp009.
- Thakur S, Yadava AK. 2021. Influence of environmental factors on tree species diversity and composition in the Indian Western Himalaya Govind Ballabh Pant. National Institute of Himalayan Environment and Development.